
黄淮麦区(南片)小麦粒重基因 TaGS-D1和 TaCwi-A1等位变异检测及效应分析
2023-04-25 09:10冉午玲陈建辉赵永涛朱保磊徐永贵宋佳静宋全昊赵立尚朱统泉
麦类作物学报 2023年4期
陈 杰,冉午玲,陈建辉,赵永涛,朱保磊,徐永贵,白 冬,宋佳静,宋全昊,金 艳,赵立尚,朱统泉
(1.河南省驻马店市农业科学院,河南驻马店 463000; 2.河南省种子管理站,河南郑州 450016;3.河南省对外科技交流中心,河南郑州 450003; 4.河南省漯河市农业科学院,河南漯河 462300;5.河南省信阳市农业科学院,河南信阳 464000)
小麦是中国重要的粮食作物,在耕地面积逐步减少和人口持续增长的背景下,提高小麦单产是提升中国小麦总产量的重要途经。小麦单产与单位面积穗数、穗粒数以及粒重密切相关,其中粒重的提高对于提升小麦单产贡献较大[1-2]。因此,研究小麦粒重对于提升中国小麦产量具有重要 意义。
小麦粒重主要受遗传因素的影响,同时也是遗传较稳定、广义遗传率较高的产量构成因素之一[3]。随着小麦全基因组序列信息的发布以及现代分子技术的应用,小麦粒重相关基因的定位、克隆和分子标记开发也取得了较快进展。从目前的定位结果来看,几乎所有染色体上都能检测到与小麦粒重相关的QTL,其中第2同源染色体组上检测到的QTL,可解释小麦粒重4.8%~20.1%的表型变异,第7同源染色体组上检测到的QTL,可解释小麦粒重7.7%~16.3%的表型变异[4-9]。随后,Zhang等[8]在小麦7B染色体短臂(7BS)上克隆出了TaGS-D1基因,并开发设计了功能标记GS7D,用于检测等位基因TaGS-D1a(与高千粒重相关)和TaGS-D1b(与低千粒重相关)。Ma等[9]在小麦2A染色体长臂(2AL)上克隆出了TaCwi-A1基因,并开发设计了功能标记CWI21和CWI22,用于检测等位基因TaCwi-A1a(与高千粒重相关)和TaCwi-A1b(与低千粒重相关)。以上功能标记GS7D、CWI22和CWI21已经在宁夏、云南、新疆等小麦材料中进行了应用[10-17]。
黄淮麦区(南片)是中国小麦重要的生产区域,提高该区域小麦的粒重对于提升国内小麦总产量具有重要意义。尽管已有该区域小麦粒重基因的研究报道[10,14],但在该区域综合TaGS-D1和TaCwi-A1两个小麦粒重基因的研究报道还很少见。因此,本研究以94份黄淮麦区(南片)新育成的小麦品种(系)为试验材料,利用电子天平对供试材料的粒重进行测定,同时利用功能标记GS7D、CWI22和CWI21对供试材料中TaGS-D1和TaCwi-A1位点的等位基因进行检测,了解该区域新育成小麦TaGS-D1和TaCwi-A1位点等位基因的分布情况,并进一步分析不同等位基因以及等位基因组合与粒重之间的关系,以期为黄淮麦区(南片)小麦粒重的遗传改良提供参考 信息。
1 材料与方法
1.1 试验材料
选用2020-2021年度参加黄淮冬麦区南片水地组区域试验的全套小麦新品种(系)为试验材料,剔除组别间重复的对照品种,共计94份(表1)。这些材料均是经过两年品种比较试验晋级上来的优秀品种(系),能够很客观地反映本麦区目前的小麦育种情况。供试材料采用随机区组排列,设置3个重复。试验材料于2020-2021年度种植于驻马店市农业科学试验站(海拔73.7 m,北纬33.01°,东经114.04°),水肥管理、防治虫害、除草等田间管理措施与当地大田生产相同。

表1 供试材料的千粒重及其基因型
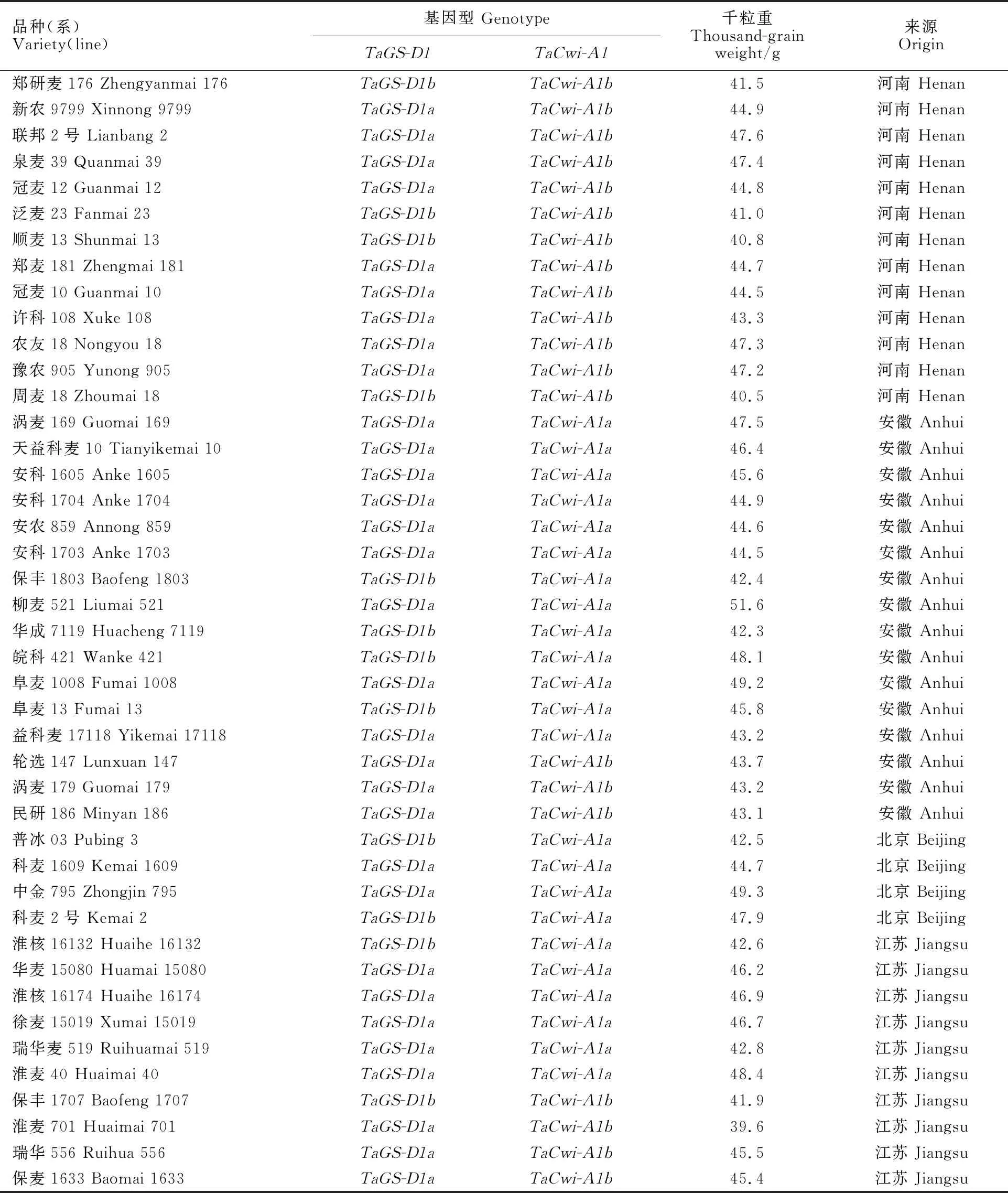
(续表1 Continued table 1)
1.2 千粒重测定
用德国Pfeuffer公司生产的全自动数粒仪(型号:Contador)对每份供试材料随机数取 1 000粒种子,用常熟市佳衡天平仪器有限责任公司生产的电子天平(型号:DT-300A)进行称重,每份材料均重复称重3次(如果两两之间的千粒重差值大于0.5 g,则需要重做),计算平均值。
1.3 等位基因的分子检测
按照陈 杰等[18]的方法快速提取试验材料的基因组DNA。利用共显性功能标记GS7D对供试材料7DS染色体上TaGS-D1位点的等位基因进行检测,扩增出562 bp 条带的材料记为TaGS-D1a等位基因类型,扩增出522 bp条带的材料记为TaGS-D1b等位基因类型。利用互补性功能标记CWI22和CWI21对供试材料2AL染色体上TaCwi-A1位点的等位基因进行检测,扩增出402 bp 条带的材料记为TaCwi-A1a等位基因类型,扩增出404 bp条带的材料记为TaCwi-A1b等位基因类型。所用标记的引物序列等详细信息见表2。PCR反应体系和扩增程序参考陈 杰等[18]的方法。扩增结束后,用1.9%的琼脂糖对PCR扩增产物进行电泳分离,利用美国Protein Simple公司生产的凝胶成像系统(AlphaImager HP)扫描成像。

表2 本研究所用小麦粒重相关基因功能标记
1.4 数据分析
利用Microsoft Excel 2003软件进行基础数据处理和表格制作,用SPSS 18.0软件进行方差分析(ANOVA法)和多重比较(LSD法)。
2 结果与分析
2.1 TaGS-D1和 TaCwi-A1位点等位基因的检测结果
在TaGS-D1位点,通过用标记GS7D进行检测,安科1605、驻麦586、西农172、郑研麦176等76份材料扩增出562 bp的条带,说明这些材料含有TaGS-D1a等位基因,分布频率为80.85%;保丰1707、西农1125、淮核16132、郑麦20等18份材料扩增出522 bp的条带,说明这些材料含有TaGS-D1b等位基因,分布频率为19.15%。图1为部分材料的检测结果。
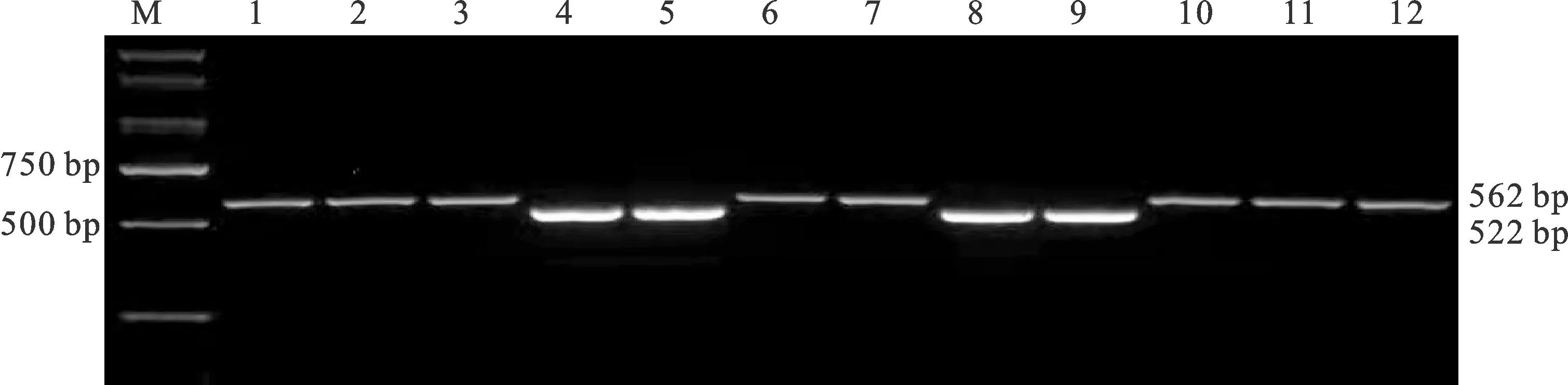
M:DL2000;1:安科1605;2:驻麦586;3:西农172;4:保丰1707;5:西农1125;6:郑研麦176;7:华麦15080;8:淮核16132;9:郑麦20;10:西农609;11:郑麦181;12:豫农905。图2同。
在TaCwi-A1位点,利用标记CWI22和CWI21对供试材料进行分子检测。结果显示,安科1605、驻麦586、西农172等63份材料扩增出402 bp的条带,说明这些材料含有TaCwi-A1a等位基因,分布频率为67.02%;保丰1707、西农1125、淮核16132等31份材料扩增出404 bp的条带,说明这些材料含有TaCwi-A1b等位基因,分布频率为32.98%。部分材料的检测结果如图2所示。
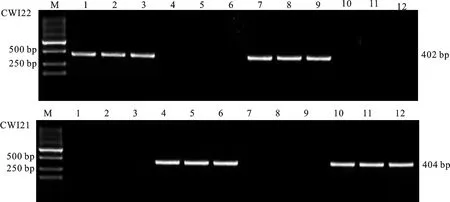
图2 标记CWI22(上)和CWI21(下)对部分供试小麦品种 TaCwi-A1位点的扩增结果
2.2 TaGS-D1和 TaCwi-A1位点等位基因与粒重的关系
从表3可知,在TaGS-D1位点,含有TaGS-D1a和TaGS-D1b等位基因材料的千粒重平均值分别为46.0和43.0 g,差异达到显著水平,表明基因型TaGS-D1a与高千粒重相关,TaGS-D1b与低千粒重相关;在TaCwi-A1位点,含有TaCwi-A1a和TaCwi-A1b等位基因材料的千粒重平均值分别为46.3和43.8 g,差异也达到显著水平,表明TaCwi-A1a等位基因与高千粒重相关,TaCwi-A1b等位基因与低千粒重相关。
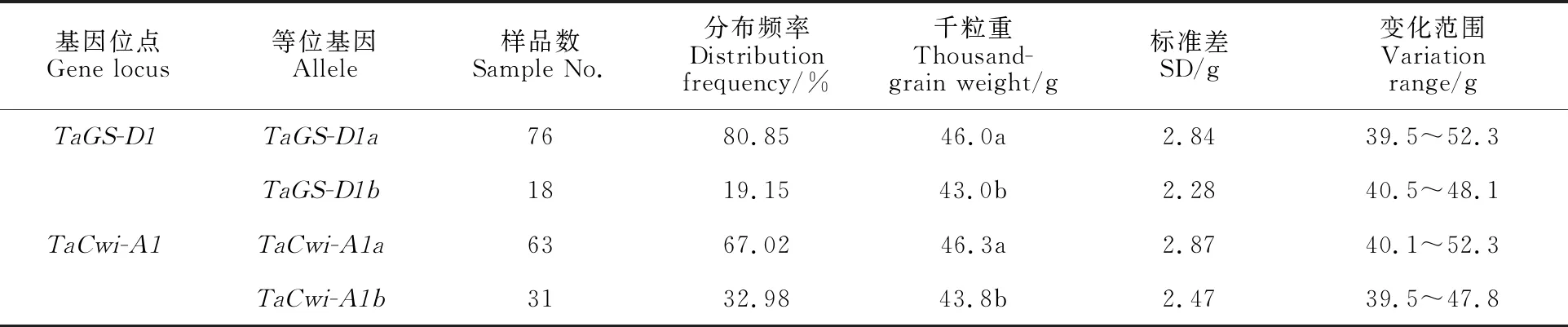
表3 不同位点等位基因对小麦粒重的影响
2.3 不同等位基因组合的分布及其与粒重的关系
94份供试材料中,TaGS-D1和TaCwi-A1位点上共检测到四种等位基因组合类型(表4)。其中含有TaGS-D1a/TaCwi-A1a等位基因组合的材料最多,有53份,分布频率为56.38%;含有TaGS-D1b/TaCwi-A1b等位基因组合的材料最少,仅有8份,分布频率为8.51%;含有TaGS-D1b/TaCwi-A1a和TaGS-D1a/TaCwi-A1b等位基因组合的材料分别有10和23份,分布频率分别为10.64%和24.47%。这表明供试材料以TaGS-D1a/TaCwi-A1a等位基因组合类型分布为主。
进一步分析不同等位基因组合对粒重的影响。结果如表4所示,可以看出,含有TaGS-D1a/TaCwi-A1a等位基因组合的材料,千粒重平均值最高,与其余三种类型差异显著。含有TaGS-D1b/TaCwi-A1b等位基因组合的材料,千粒重平均值最低,与其余三种类型也差异显著。含有其余两种等位基因组合类型的材料之间,千粒重平均值差异不显著。这表明TaGS-D1a/TaCwi-A1a等位基因组合与高千粒重相关,TaGS-D1b/TaCwi-A1b等位基因组合与低千粒重相关,TaGS-D1b/TaCwi-A1a和TaGS-D1a/TaCwi-A1b等位基因组合与中等千粒重相关。
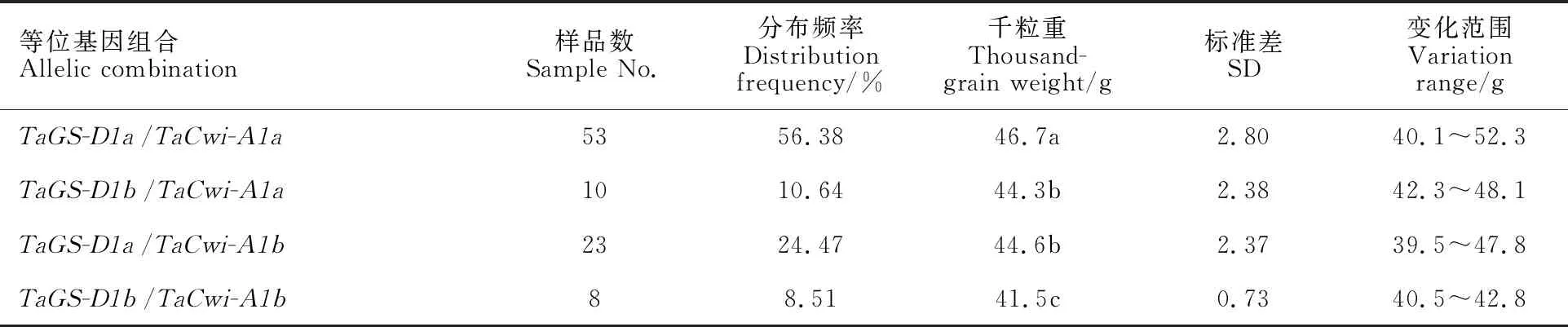
表4 不同等位基因组合对粒重的影响
3 讨 论
了解TaGS-D1和TaCwi-A1位点等位变异的分布情况,有利于小麦粒重的遗传改良。研究表明,TaGS-D1a和TaGS-D1b等位基因在黄淮麦区小麦中的分布频率分别为66.7%和33.3%[10],在新疆小麦中的分布频率分别为70.6%和29.4%[11],在宁夏小麦中的分布频率分别为89.83%和10.17%[12],在北部冬麦区小麦中的分布频率分别为85.5%和14.5%[13];TaCwi-A1a和TaCwi-A1b等位基因在黄淮麦区小麦中的分布频率分别为65.03%和34.97%[14],在新疆小麦中的分布频率分别为77.2%和22.8%[15]。本研究结果表明,等位基因TaGS-D1a、TaGS-D1b、TaCwi-A1a和TaCwi-A1b在94份黄淮麦区(南片)小麦新品种(系)中的分布频率分别为80.85%、 19.15%、67.02%和32.98%。根据以上数据分析可知,与高粒重相关的等位基因TaGS-D1a和TaCwi-A1a在上述各个麦区和各个省份小麦中的分布频率虽然有差异,但是均高于与低粒重相关的等位基因TaGS-D1b和TaCwi-A1b的分布频率。高产是育种家们追求的重要育种目标之一,粒重又是影响小麦高产的主要因素之一,在育种过程中育种家们普遍选择了高粒重的小麦材料,因此造成了TaGS-D1a和TaCwi-A1a基因型在中国小麦中的分布频率高于TaGS-D1b和TaCwi-A1b基因型。
明确TaGS-D1和TaCwi-A1位点等位基因型与粒重之间的关系,可以为小麦粒重的分子标记辅助选择提供理论基础。本研究结果表明,含有TaGS-D1a和TaCwi-A1a等位基因材料的千粒重均显著高于其对应位点上的TaGS-D1b和TaCwi-A1b等位基因。这与Zhang等[8]和Ma等[9]的研究结果一致,验证了TaGS-D1a和TaCwi-A1a等位基因与高粒重相关,TaGS-D1b和TaCwi-A1b等位基因与低粒重相关,这也表明标记GS7D、CWI22和CWI21可作为小麦粒重分子辅助选择的有效工具。本研究也发现TaGS-D1a/TaCwi-A1a等位基因组合与高千粒重相关,TaGS-D1b/TaCwi-A1b等位基因组合与低千粒重相关,但是从被研究材料中可以看出含有TaGS-D1a/TaCwi-A1a等位基因组合小麦材料的千粒重数值也有低于含有TaGS-D1b/TaCwi-A1b等位基因组合小麦材料的状况,造成这种情况的原因可能是在小麦其他染色体上也存在影响粒重的基因。Jiang等[19]和Yan等[20]研究表明,普通小麦2B染色体上的TaSus2-2B基因和7B染色体上的TaGw8-B1基因也会对粒重造成影响。另外,本研究筛选的西农161、漯麦66、憨丰3468、平安12、郑麦163、柳麦521、富麦701和泛麦26不仅千粒重高于50 g,而且均携带有与高粒重相关的TaGS-D1a/TaCwi-A1a等位基因组合,可作为选育高粒重小麦新品种的亲本资源,在组配杂交组合过程中进行利用。
猜你喜欢
种子(2023年9期)2023-11-22
作物学报(2022年3期)2022-01-22
智慧健康(2021年17期)2021-07-30
麦类作物学报(2021年3期)2021-05-25
北京农学院学报(2020年1期)2020-03-09
种业导刊(2019年5期)2019-01-04
种业导刊(2018年11期)2018-11-17
现代检验医学杂志(2016年5期)2016-08-20
法医学杂志(2015年4期)2016-01-06
甘肃农业科技(2014年8期)2014-01-16
