
干旱胁迫下大苞萱草WRKY 基因家族成员鉴定及生物信息学分析
2023-04-07 07:52:40陈丽飞李嘉峻刘云怡慧周蕴薇
河南农业科学 2023年2期
陈丽飞,李嘉峻,刘云怡慧,白 云,孟 缘,李 江,周蕴薇
(1.吉林农业大学 林学与草学学院,吉林 长春 130118;2.定西市水土保持科学研究所,甘肃 定西 743000;3.吉林农业大学 园艺学院,吉林 长春 130118)
萱 草(Hemerocallisspp.)属 阿 福 花 科(Asphodelaceae)萱草属(Hemerocallis),多年生草本植物,民间多称其“忘忧草”[1-2]。萱草属植物分布广泛、适应性强、品种繁多、观赏价值高,被称为中国的“母亲花”。大苞萱草(Hemerocallis middendorffii)是现代萱草的原始亲本之一,主要分布在我国东北、华北等地。大苞萱草抗旱能力强、花色艳丽,而且具有二次开花特性,可广泛应用于各类园林绿地或进行盆栽装饰。
干旱不仅限制着植物的正常生长发育,还会影响植物体的代谢功能,降低光合速率、蒸腾速率以及抑制植物生长[3]。干旱是植物非生物胁迫的主要原因之一,迫使植物发生形态和生理变化[4]。植物对干旱胁迫的响应主要表现为根系和叶片结构的改变,生理水平主要表现为光合作用、渗透调节代谢、抗氧化代谢及激素物质等方面的变化,而分子水平则是调控抗旱调节基因和功能基因的表达,改变功能性蛋白和有机分子合成酶的编码,进而影响干旱信号转导以及激素、渗透物质的合成和积累[5]。
WRKY 转录因子的主要特征在于其包含60 个左右氨基酸DNA 结合区的WRKY 域,其中包含一个高度保守的七肽序列(WRKYGQK)。WRKY 转录因子在植物防御生物及非生物胁迫中发挥着关键作用[6]。超表达的WRKY 家族基因能够帮助植物在干旱环境中维持较好的水分状况,缓解因干旱所造成的萎蔫及生长抑制[7]。在大豆研究中,过表达的GmWRKY54可能通过激活ABA 以及Ca2+信号通路来控制气孔的关闭,增强大豆的抗旱能力[8]。在干旱胁迫下,植物会受到诸如活性氧(ROS)损伤等次生胁迫损伤。VlWRKY48能提高葡萄过氧化氢酶、过氧化物酶和超氧化物歧化酶等抗氧化酶活性,清除活性氧来提高其抗旱性[9]。
目前,对于大苞萱草的开发利用还不充分,关于大苞萱草在分子层面响应干旱胁迫的研究鲜见报道,不利于深入研究大苞萱草的抗旱机制。鉴于此,以大苞萱草为试验材料,基于转录组数据,采用生物信息学分析方法对干旱胁迫下的大苞萱草WRKY 基因(HmWRKY)进行鉴定、构建进化树、分析和预测蛋白质理化性质以及蛋白质二级结构等,根据HmWRKY基因的表达特性,明确候选基因,为进一步研究HmWRKY基因功能奠定基础。
1 材料和方法
1.1 材料及处理
供试材料大苞萱草取自吉林农业大学野生花卉资源圃,选取田间长势一致且植株健壮的大苞萱草幼苗移栽至高13 cm、直径10 cm 的塑料花盆中,单株单盆,栽培基质为草炭、珍珠岩按体积比3∶1混合配制成的营养土,进行正常的盆栽管理。缓苗后,将供试材料分为对照组(CK 组)和干旱胁迫处理组(D组)。干旱处理前使各组均处于饱和含水状态,处理后,CK 组每3 d 浇一次水,D 组采取自然失水胁迫的方式处理。于5 d 后的10:00 对CK 组和D组进行取样,每组处理设置3个重复,将嫩叶和新根放入液氮中带回并存放在-80 ℃冰箱保存,备用。RNA 的提取、质控、建库及转录组的三代测序均由北京诺禾致源科技股份有限公司完成。
1.2 HmWRKY基因家族成员鉴定
以大苞萱草转录组测序结果为材料。在TAIR网 站(https://www.arabidopsis.org/)下 载 拟 南 芥WRKY 基因家族的转录因子序列和蛋白质序列,共72 个。通过本地blast+进行比对,筛选出大苞萱草的WRKY 转录因子。利用Smart(http://smart.emblheidelberg.de/)在线软件进行鉴定以确保含有WRKY 保守域,通过NCBI(https://blast.ncbi.nlm.nih.gov)在线软件进行结构域完整性的预测。
1.3 HmWRKY基因家族成员生物信息学分析
1.3.1 HmWRKY 基因家族聚类分析及多重序列比对 利用MEGA 7 对HmWRKY 转录因子蛋白构建进 化 树(Neighbor-joining,NJ),Bootstrap 设 置 为1 000次重复,其余参数选择默认值[10]。利用序列分析软件DNAMAN 6.0 对HmWRKY 蛋白序列进行多重序列比对,截取包含WRKY 结构域与锌指结构的氨基酸序列,通过TBtools 软件的SeqLogo 绘制HmWRKY转录因子保守结构域的logo图。
1.3.2 HmWRKY 基因家族的理化性质分析 利用ExPASy ProtParam tool(https://web.expasy.org/protparam/)对HmWRKY蛋白的理化性质进行预测,其中包括氨基酸序列长度、分子质量、蛋白质等电点、总平均亲水性、不稳定系数以及脂溶性系数。使 用CELLO v.2.5(http://cello.life.nctu.edu.tw/)对HmWRKY 家族成员进行植物细胞的亚细胞定位,整理数据进行汇总。
1.3.3 HmWRKY 蛋白二级结构预测 利用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa-sopma.html)在线软件对大苞萱草WRKY蛋白质二级结构进行预测。
1.3.4 HmWRKY 蛋白保守基序分析 通过在线软件MEME(https://meme-suite.org/meme/tools/meme)对HmWRKY 蛋白保守基序进行搜索,搜索参数设置为最大搜索数量20 个,ZOOPS 参数设置为Zero or one occurrence per sequence,其余参数使用网页默认设置。最后使用TBtools将HmWRKY转录因子蛋白保守基序可视化。
1.3.5 HmWRKY 基因家族在叶和根中的表达分析 利用北京诺禾致源科技股份有限公司提供的售后平台工具NovoMagic(https://magic.novogene.com/customer/main#/login)分别制作处理组与对照组叶和根中HmWRKY基因家族41个成员的表达模式热聚图。
1.4 HmWRKY 基因实时荧光定量PCR(qRTPCR)分析
将公司测序后剩余的样品进行液氮研磨,使用RNApure Plant Kit(DNase Ⅰ)植物RNA 提取试剂盒(DNase Ⅰ)提取叶片总RNA,使用NovoScript®Plus All-in-one 1st Strand cDNA Synthesis(gDNA Purge)反转录试剂盒合成cDNA,使用NovoStart®SYBR qPCR SuperMix Plus 试剂盒进行qRT-PCR,使用Primer 5.0进行qRT-PCR 引物设计(表1),内参基因为HfEF-1α(GenBank No.MT096368)[11]。qRT-PCR反应体系为2×SYBR Mix 主混合物10 μL,上下游引物各1 μL,模板(cDNA)2 μL,ddH2O 6 μL。反应程序:预变性95 ℃1 min;95 ℃20 s、60 ℃20 s、72 ℃30 s,40 个循环。试验进行3 次生物学重复,采用2-ΔΔCt方法计算基因的相对表达量[12]。

表1 HmWRKY基因qRT-PCR引物序列Tab.1 qRT-PCR primer sequences for HmWRKY gene
2 结果与分析
2.1 HmWRKY基因家族成员鉴定
通过本地blast+与拟南芥WRKY 转录因子进行比对,从转录组测序结果中共筛选出182 个大苞萱草WRKY 转录因子。利用在线软件Smart鉴定以确保含有WRKY 保守域,通过NCBI 在线软件预测结构域的完整性。最终得到41 个大苞萱草WRKY 转录因子,并将这些基因命名为HmWRKY1—HmWRKY41。
2.2 HmWRKY基因家族成员生物信息学分析
2.2.1 HmWRKY 基因家族聚类分析及多重序列比对 将41 个HmWRKY 蛋白与72 个拟南芥WRKY蛋白进行多序列分析并通过软件构建进化树(图1)。WRKY DNA 结构域(DBD)的特征是在其N 端有一个不变的七肽WRKYGQK 氨基酸基序,在其C端包含一个锌指结构。根据WRKY 结构域的数量和锌指状基序的特征将WRKY 转录因子分为Ⅰ、Ⅱ、Ⅲ三类,其中Ⅱ类分为5个亚类。

图1 大苞萱草和拟南芥WRKY转录因子聚类分析Fig.1 Cluster analysis of WRKY transcription factors in Hemerocallis middendorffii and Arabidopsis thaliana
WRKY 结构域高度保守,多重序列比对结果如图2 和图3 所示。WRKYGQK 的氨基酸序列存在替换的现象。Ⅰ类共有8个HmWRKY转录因子,含有2 个WRKY 结构域,锌指结构为CX7CX24~25HXH 型。Ⅱ类共有30 个HmWRKY 转录因子,只含有1 个WRKY 结构域且锌指结构为CX7CX25HXH 型,Ⅰ类和Ⅱ类转录因子均有不同程度的七肽结构变异和锌指结构缺失的现象。Ⅲ类有4 个HmWRKY 转录因子,含有1 个WRKY 结构域,锌指结构为CX7CX25HXC 型,Ⅲ类HmWRKY 转录因子相对保守,七肽结构不发生变异且锌指结构完整。

图2 HmWRKY转录因子家族保守结构域分析Fig.2 Conservative domain analysis of HmWRKY transcription factor family

图3 HmWRKY转录因子家族保守结构域分析Fig.3 Fig.3 Conservative domain analysis of HmWRKY transcription factor family
2.2.2 HmWRKY 基因家族的理化性质分析HmWRKY 基因家族成员的理化性质差异性较大(表2),HmWRKY 转录因子的氨基酸数目在92(HmWRKY11)~641(HmWRKY7),分 子 质 量 在10 551.76(HmWRKY11)~69 972.1(HmWRKY7)ku,理论等电点在4.97(HmWRKY26)~9.99(HmWRKY24),脂溶性指数在47.08(HmWRKY26)~75.93(HmWRKY6),41 个HmWRKY 转录因子的亲水性平均值均为负值,判定为亲水性蛋白。除HmWRKY2、HmWRKY4、HmWRKY5、HmWRKY27、HmWRKY41 为稳定蛋白外,其余均为不稳定蛋白(不稳定系数>40)。HmWRKY 家族成员亚细胞定位结果如表3所示,除HmWRKY41 定位在线粒体中外,其余的HmWRKY转录因子均定位在细胞核中。

表2 HmWRKY转录因子蛋白的理化性质Tab.2 Physicochemical properties of HmWRKY transcription factor proteins
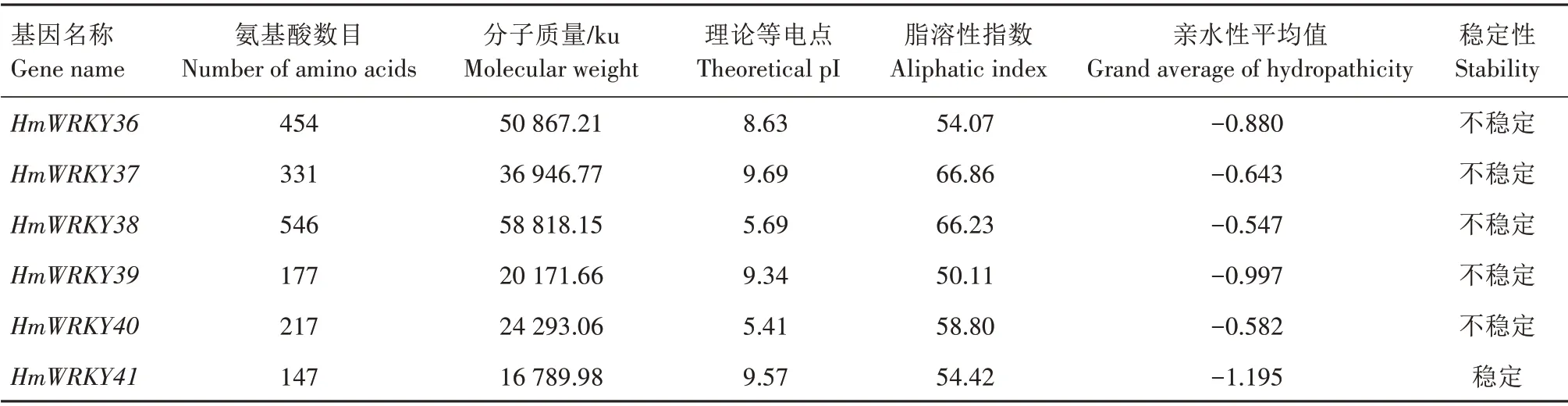
续表2 HmWRKY转录因子蛋白的理化性质Tab.2(Continued)Physicochemical properties of HmWRKY transcription factor proteins
2.2.3 HmWRKY 蛋白的二级结构预测 41 个HmWRKY 转录因子蛋白二级结构均含有α-螺旋、β-折叠、延伸链以及无规卷曲,无其他二级结构(表4)。绝大多数HmWRKY 蛋白都表现出无规卷曲数>α-螺旋数≥延伸链数>β-折叠数的特点,但少数蛋白质,如HmWRKY11、HmWRKY16、HmWRKY17等表现出无规卷曲数>延伸链数>α-螺旋数>β-折叠数的特点。

表4 HmWRKY转录因子蛋白二级结构Tab.4 Secondary structure of HmWRKY transcription factor proteins %

续表4 HmWRKY转录因子蛋白二级结构Tab.4(Continued)Secondary structure of HmWRKY transcription factor proteins %
2.2.4 HmWRKY 蛋白保守基序分析 HmWRKY蛋白序列的保守基序预测结果见图4。Motif 1 是WRKY 转录因子的特征基序且组成了WRKY 结构域。除HmWRKY24、HmWRKY26、HmWRKY27 和HmWRKY30外,其他成员均包含Motif 1。Motif 2和Motif 5 共同构成C2H2型的锌指结构域,存在于大多数HmWRKY 转录因子中。Ⅰ类成员几乎全包含Motif 5、Motif 17 以 及 在N 端、C 端 分 别 含 有1 组Motif 1 和Motif 2。Ⅱ类成员的Motif种类繁多,但呈现出在进化树中亲缘关系较近的各亚类成员,包含Motif 种类也基本一致的规律,如Ⅱa 亚类与Ⅱb 亚类 除HmWRKY2 外,均 包 含Motif 19;Ⅱc 亚 类HmWRKY33 和HmWRKY39 的Motif 种类完全相同且成员多数包含Motif 4;Ⅱd 亚类HmWRKY14 和HmWRKY37包含的Motif种类以及排列顺序完全相同,其他成员几乎都包含Motif 7。Ⅲ类成员中存在的Motif种类比较单一,只包含Motif 1和Motif 2。

图4 HmWRKY转录因子家族保守基序Fig.4 Conserved motif of HmWRKY transcription factor family
2.2.5 HmWRKY 基因家族在叶和根中的表达分析 根据转录组数据的分析,对比HmWRKY 基因家族的41 个成员在不同处理下叶和根中的表达见图5,在叶中有10 个上调基因,29 个下调基因,其中HmWRKY25和HmWRKY32在 叶 中 无 表 达。HmWRKY22、HmWRKY9上 调 较 明 显,HmWRKY2、HmWRKY11、HmWRKY12、HmWRKY20、HmWRKY26、HmWRKY30、HmWRKY34和HmWRKY36下 调 较 明显。41 个HmWRKY基因在根中均有表达,其中有20 个上调基因,21 个下调基因。HmWRKY4、HmWRKY8、HmWRKY14、HmWRKY15、HmWRKY22、HmWRKY23、HmWRKY24、HmWRKY35在受到干旱胁迫时,表达量上调较为明显。HmWRKY2、HmWRKY16、HmWRKY27、HmWRKY30、HmWRKY41在下调基因中表达量变化较为明显。
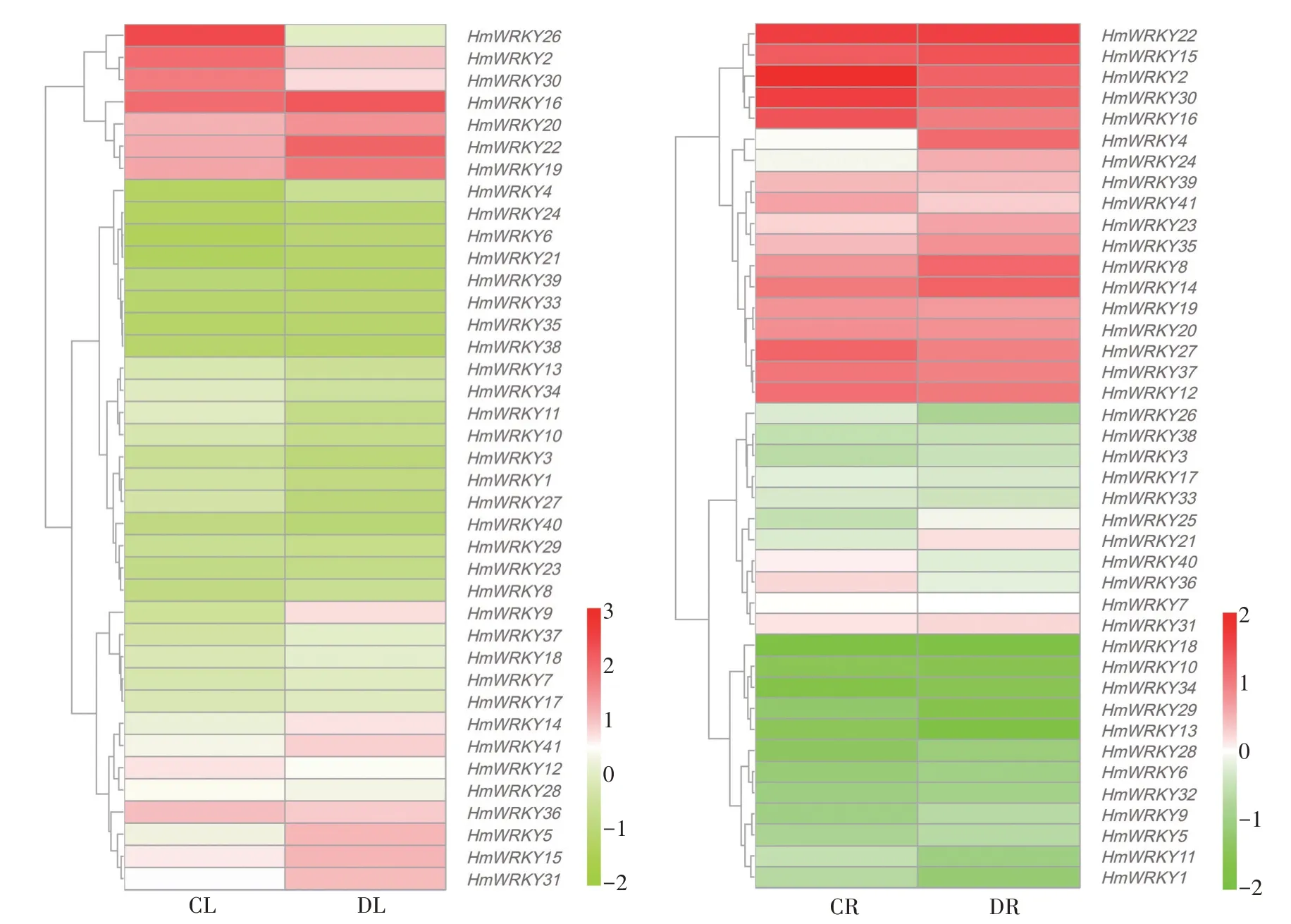
图5 大苞萱草叶片和根中HmWRKY基因在不同处理下的表达模式热聚图Fig.5 Thermopolygram of HmWRKY gene expression pattern under different treatments in leaves and roots of Hemerocallis middendorffii
HmWRKY差异基因的表达变化如图6 所示,在叶中共发现10 个差异基因,1 个上调基因,9 个下调基因。大苞萱草受到干旱胁迫后,只有HmWRKY9的表达量升高。下调的差异基因中HmWRKY2、HmWRKY26和HmWRKY30的表达量变化显著。在根中发现4 个差异基因,其中3 个上调基因,1 个下调基因。HmWRKY4和HmWRKY24上调趋势比较明显,可能在根系中起到积极的调控作用。此外,通过叶和根的差异基因比对发现,HmWRKY2在2个组织内均有表达且均为负表达,表达量变化十分明显。
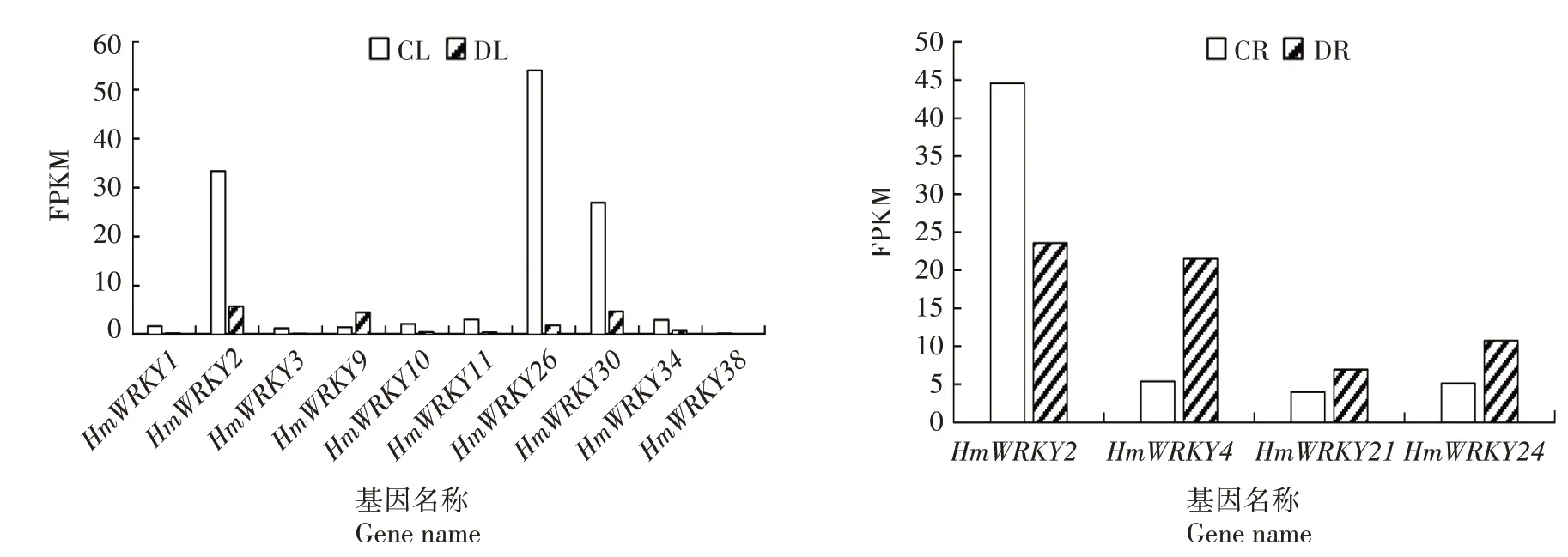
图6 大苞萱草叶片和根中HmWRKY差异基因在不同处理下的表达水平Fig.6 Expression of HmWRKY differential gene under different treatments in leaves and roots of Hemerocallis middendorffii
2.3 候选差异基因的qRT-PCR验证
为了验证候选差异基因HmWRKY9在叶中表达量的正确性,进行qRT-PCR 分析。结果如图7 所示,HmWRKY9在受到干旱胁迫后,表达量升高,正向响应了干旱胁迫。HmWRKY9的qRT-PCR 结果与转录组测序的变化趋势基本一致,从而验证了转录组数据的准确性。
3 结论与讨论
WRKY 基因家族在植物体中响应生长发育、生物胁迫以及非生物胁迫等过程,在非生物胁迫中对干旱胁迫反应的调控网络发挥重要作用。CmWRKY10通过ABA 途径来调控菊花的干旱耐受性[13],水稻在干旱胁迫条件下过表达OsWRKY11可增强其耐旱性[14],过表达AtWRKY30转基因植株能够显著提高小麦的耐旱能力[15]。以上研究均揭示了WRKY转录因子家族成员可提高植物抗旱能力。
本研究基于转录组测序,共挖掘出41 个HmWRKY 转录因子。将HmWRKY 基因家族与已公布的拟南芥WRKY 基因共同构建进化树并进行聚类分析,HmWRKY 基因家族的分类与拟南芥WRKY 基因家族的分类一致,表明本试验转录组数据具有可靠性,HmWRKY 基因家族在进化上相对保守。WRKY 转录因子的七肽和锌指结构是WRKY 基因行使功能的两大基团,随着植物进化,这两大基团的氨基酸组成也有所变化。Ⅰ类HmWRKY 转录因子中只有HmWRKY41N 端的七肽结构发生变异,变异为“WRKYGPK”,Ⅱ类HmWRKY 转录因子的七肽结构易发生变异,其中变异为“WKKYGQK、WRKYGKK、WRKYGEK”[16-17]。Ⅲ类HmWRKY 转录因子的结构域相比前两类保守,没有发生变异情况。ZHANG 等[16]针对真核生物WRKY 基因的研究发现,WRKY 家族成员有19 个WRKY 结构域的变异,WRKYGEK 和WRKYGKK 是2 个常见的变异。EULGEM 等[17]在香蕉和杨树中也发现七肽变异类型为WRKYGKK 的WRKY 转录因子蛋白,其中的变异类型与本研究结果一致。YU等[18]认为高度保守的WRKY 结构域可以特异识别靶基因启动子区内的W-BOX 元件从而调控该基因的活性,进一步发挥其生物学功能。因此,具有变异的WRKY 结构域的HmWRKY 转录因子可能会影响其发挥生物学功能。保守基序分析表明,绝大多数HmWRKY 蛋白含有的特征基序构成不同,如Ⅰ类成员多数含有2 组Motif 1、Motif 2 和Motif 5,Ⅱa和Ⅱb 亚类的特征基序是Motif 19,Ⅱc 亚类特征基序为Motif 4,Ⅱd和Ⅱe亚类的特征基序均为Motif 7,而Ⅲ类成员中只存在2 种Motif,保守基序种类不同说明这些成员在进化过程中发生了不同程度的变异,这可能与基因家族的分类及功能调控有关。
通过对干旱胁迫下HmWRKY基因的表达分析发现,41个HmWRKY基因在叶和根中的表达量变化均有不同,少数HmWRKY基因如HmWRKY6、HmWRKY29等在干旱胁迫前后叶和根中的表达量变化不明显,而HmWRKY22、HmWRKY36等在处理前后叶和根中的表达量变化相对显著。另外,HmWRKY6在干旱处理前不表达,处理后开始表达,HmWRKY21和HmWRKY38则相反,说明HmWRKY6、HmWRKY21和HmWRKY38可能与大苞萱草的抗旱性相关。已有研究表明,拟南芥中部分WRKY 转录因子可在植物抵御干旱胁迫的过程中起到重要调控作用。AtWRKY28和AtWRKY57均能提高植株的耐旱性[19-20]。过表达AtWRKY29可使拟南芥植株变小且叶片颜色变深,增强其抗旱能力[21]。AtWRKY33可通过直接调控CesA8 参与干旱胁迫[22]。AtWRKY40可以通过增强植株的渗透调节能力和抗氧化能力来维持细胞吸水,从而提高植株对干旱胁迫的抵抗力[23]。过表达AtWRKY57的拟南芥耐旱性显著提高[19]。本研究通过大苞萱草转录组测序结果得到13个差异基因,部分差异基因与上述拟南芥抗旱基因具有较近的亲缘关系,如HmWRKY2、HmWRKY3、HmWRKY4、HmWRKY9、HmWRKY30和HmWRKY34。这 也 间 接 表 明 ,HmWRKY2、HmWRKY3、HmWRKY4、HmWRKY9、HmWRKY30和HmWRKY34可能通过促进或抑制自身的表达来参与大苞萱草响应干旱胁迫的过程。大苞萱草受到干旱胁迫的前后,HmWRKY2均存在于叶和根中,HmWRKY2的表达量变化均为显著下降且HmWRKY2与AtWRKY40同 源 性 较 高。因 此,HmWRKY2可能通过抑制其表达在大苞萱草抵御干旱胁迫的过程中发挥重要作用。
本研究基于大苞萱草的转录组数据,首次对大苞萱草WRKY 基因家族进行鉴定,初步确定了41个HmWRKY基因。通过对HmWRKY 转录因子的结构特征及在不同处理下叶和根中表达量的综合分析,揭示了大苞萱草WRKY 基因家族成员可能与大苞萱草抗旱方面相关,为HmWRKY 转录因子家族的功能研究以及抗性基因筛选提供参考。确定HmWRKY9为候选基因,后续将开展HmWRKY9的功能验证工作,探讨其是否与大苞萱草的抗旱性有关。
猜你喜欢
新医学(2023年10期)2023-12-09 15:04:51
南方医科大学学报(2022年3期)2022-04-13 01:51:26
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10 07:07:36
紫禁城(2020年5期)2021-01-07 02:13:34
广州大学学报(自然科学版)(2019年1期)2019-05-07 01:33:26
天津科技大学学报(2016年1期)2016-02-28 16:59:45
海峡姐妹(2016年7期)2016-02-27 15:21:17
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10 08:41:53
人间(2015年19期)2016-01-04 12:46:44
园艺与种苗(2015年10期)2015-02-27 11:27:51
