
土壤养分、林分结构和地形因子对毛竹更新的影响1)
2023-03-28 01:13:24宋贤芬赵各进严夏帆陆雪婷刘健余坤勇
东北林业大学学报 2023年3期
宋贤芬 赵各进 严夏帆 陆雪婷 刘健 余坤勇
(福建农林大学,福州,350002)
In order to explore the regeneration effect of Phyllostachys pubescens after strip cutting and the key factors affecting the restoration and regeneration, we studied the effect of soil nutrients, stand structure and topographic factors on the restoration and regeneration of P. pubescens before the cutting by taking the pure forest of P. pubescens in the following year after strip cutting as the object. The soil nutrient content had a significant effect on Hsinchu’s average DBH, average height, and the number of plants in the plot, among which soil total carbon was the most significant factor. The average height of the original stand had a significant impact on the average diameter at breast height and average height of Hsinchu, the age uniformity of the original stand directly affected the average height of Hsinchu, and the original stand structure had no significant effect on the number of Hsinchu trees. There are significant differences in the number of Hsinchus in different terrains, and there is no significant difference in the average diameter at breast height and average height. The number of Hsinchu trees in different terrains is in the descending orders of downslope; middle slope and uphill; shady slope and sunny slope;and slope 1°-20° and slope 21°-40°, respectively. The interaction of topographic factors has no significant effect on the average diameter at breast height, average height and plant number of Hsinchu. The interaction of soil nutrients, stand factors and topographic factors explained the large-scale variation in DBH and bamboo height in Hsinchu. Improving the growth and regeneration of Hsinchu in the following year after strip cutting can be achieved by controlling environmental factors such as soil nutrients.
毛竹(Phyllostachysedulis)是我国南方主要竹种,根据第九次全国森林资源调查结果显示,毛竹资源面积占竹林资源72.96%[1]。但是随着毛竹经营成本的增加、人力资源的紧缺以及经营难度提高等因素的影响[2],部分区域的毛竹林逐渐出现荒芜的现象,极大地影响了毛竹资源的有效利用。带状采伐是一种效率高、成本低的采伐方式[3]。毛竹资源的规模化、机械化经营是解决成本上升、人力资源不足、区域竹资源荒芜的关键。
植被恢复与生态系统的演化高度相关,地形、土壤、植被、水文都会导致植被动态分布格局、物种、生物量和群落结构的空间异质性[4]。幼苗更新与环境条件、林分结构、人为干扰等因子相关[5];土壤养分是立地质量重要组成成分,也直接影响林地的生产力水平和植被更新[6];土壤氮、磷是限制林分发育的重要元素,也是限制毛竹的生长的营养元素[7];林分状况不同,其幼林的恢复更新状况也有所差异[8],竹龄和林分密度直接影响幼苗的更新密度[9];地形是重新分配自然资源的主要因素之一[10],地形通过影响土壤水分、温度、光照的移动分配影响幼苗的生长[11],坡向和坡位等地形因子影响植物的物种数量、生物量和碳储量的分布[12-13]。
毛竹带状采伐后林下群落结构动态[14]、土壤微生物群落结构[15]及土壤理化性质的变化[16]等对毛竹林的恢复更新具有重要影响。为了揭示毛竹带状采伐恢复更新的趋势,以及采伐前土壤养分、林分结构和地形因子对带状采伐后毛竹恢复更新的影响,制定更有效的毛竹经营措施,促进区域林农的经济收入。本研究以福建省永安市上坪乡毛竹林作为对象,布设28个面积为20 m×9 m的样地,实施带状采伐处理,分析带状采伐前林分资源对采伐后1年生毛竹林恢复更新的影响,为带状采伐经营技术提供理论基础。
1 研究区概况
研究区位于永安市上坪乡,上坪乡位于永安市东部,距市区22 km,海拔1 120 m,属高山区,昼夜温差大,气温-5~39 ℃,与青水、槐南乡交接,交通便利。土壤类型主要以黄壤为主,乔木主要有少量马尾松(Pinusmassoniana)和杉木(Cunninghamialanceolata),灌木主要有石楠(Photiniaserrulata)、冬青(Ilexchinensis)、山茶(Camelliajaponica)、红花檵木(Loropetalumchinense)、鹅掌柴(Scheffleraoctophylla)等,草本主要有沿阶草(Ophiopogonbodinieri)、芒萁(Dicranopterisdichotoma)、狗脊(Cibotiumbarometz)、冷水花(Pileanotata)等。
2 材料与方法
2.1 方案设计及样地设置
2020年12月,在研究区布设28个面积为20 m×9 m的毛竹纯林样地,样地之间的隔离带大于3 m。在每个样地内进行林分和地形调查,并采集土壤用于理化性质测定。2020年12月对设置的所有样地进行采伐处理,翌年4月份在毛竹的出笋期对新竹进行调查。
2.2 数据获取及处理
2.2.1 林分因子
林分因子包括空间结构和非空间结构因子。空间结构因子包括角尺度和大小比数等;非空间结构因子包括平均胸径、平均树高、林分年龄、林分密度等。2020年12月,对样地内胸径大于3 cm的所有毛竹每木检尺,包括胸径和竹高调查,记录每棵毛竹的年龄,年龄包含Ⅰ度竹、Ⅱ度竹、Ⅲ度竹等,并实测每株毛竹的相对位置。在2021年4月,对带状采伐后恢复更新的1年生毛竹进行调查,调查中要求只测定毛竹胸径大于3 cm的毛竹,记录毛竹胸径、树高、株数。将采集到的数据进行整理,计算林分平均胸径;根据不同龄级毛竹株数的分布均匀程度来代表年龄结构[17]。
年龄均匀度(A):
式中:S为龄级数,Pi是第i龄级的株数比例。
根据毛竹的相对坐标及胸径计算角尺度和大小比数,参考仇建习等[18]林分结构参数计算方法。
角尺度是最近相邻木角小于标准角a0的个数占相邻木4株的比例,用公式表示为:
式中:Wi为第i株对象竹角尺度,当第j个a角小于标准角a0时,aij=1,反之,aij=0。
大小比数是大于对象木的最近相邻木占所考察的4株最近相邻木的比例,公式为:
式中:Ui为第i株对象竹大小比数;相邻竹j胸径大于对象竹i时,Kij=1,反之,Kij=0。
2.2.2 地形因子
实际调查每个样地的地形因子,调查内容包括坡度、坡向、坡位。以样地的分布情况为基础,将地形因子分为不同水平。参考范叶青等[19]的划分标准,将坡度分为缓坡、陡坡;坡位分为上坡、中坡和下坡;坡向分为阴坡(北面、东北面、东面、东南面)和阳坡(南面、西南面、西面、西北面)。
2.2.3 土壤样品采集及测定
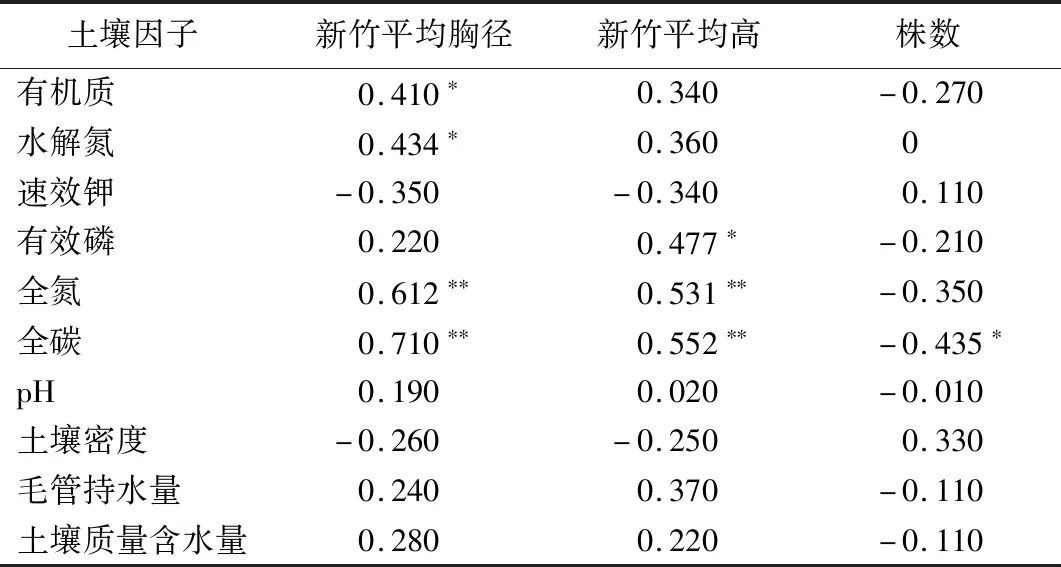
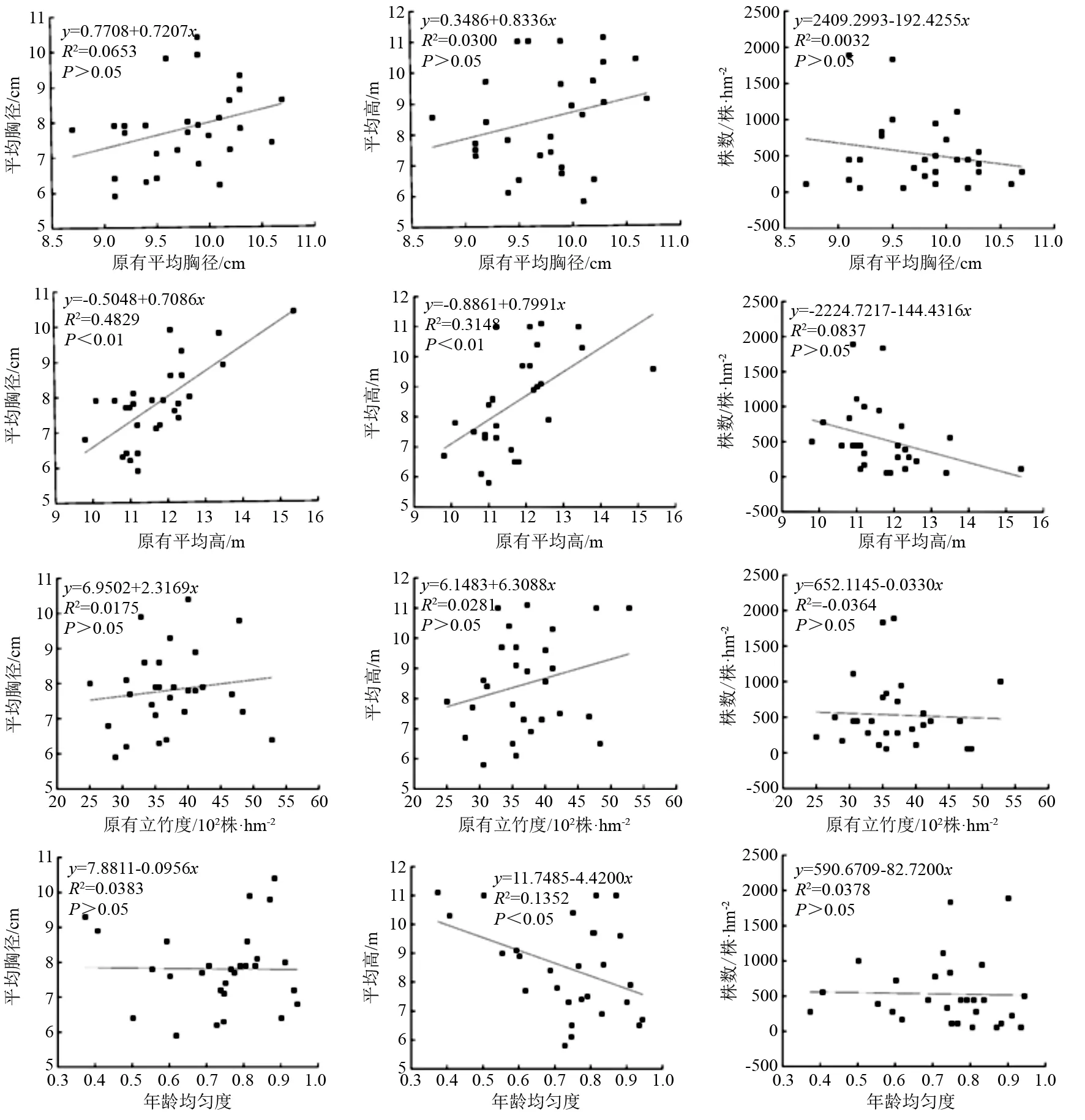
土壤采集时间为2020年12月。在样地内沿对角线选择3个点进行土样采集,每个点深挖40 cm,去掉土壤表层凋落物,分别采集深度(h)分别为0 土壤pH的测定采用水浸提电位测定法;采用重铬酸钾氧化-外加热法测定土壤有机质的质量分数;采用碱解-扩散法测定土壤水解氮的质量分数;使用VARIO MAX碳氮元素分析仪(产商:德国-elementar)测定土壤碳、氮的质量分数;使用PE OPTIMA 8000电感耦合等离子体质谱仪(产商:美国珀金埃尔默企业管理)测定土壤有效磷和速效钾的质量浓度;土壤含水率、土壤密度等采用环刀法。样地基本情况见表1。 表1 样地林分资源概况 统计量速效钾质量浓度/mg·L-1有效磷质量浓度/mg·L-1全氮质量分数/mg·kg-1全碳质量分数/mg·kg-1土壤密度/g·cm-3毛管持水量/g·kg-1含水量/g·kg-1pH最大值53.290.70390.465590.821.45586.57388.744.85平均值39.200.29312.584131.791.22477.52294.704.34最小值18.490.09206.692699.901.05322.50182.563.89标准差8.820.1444.61866.780.1176.3250.010.21变异系数/%22.5148.4014.2720.988.8715.9816.974.91 2.2.4 数据处理 使用Excel2013和SPSS25进行数据统计和分析,运用相关性分析、线性拟合和逐步回归分析法进行分析。 由表2可知, 28个样地内共调查毛竹267株,随着毛竹平均胸径逐渐增大,每个径阶株数呈现先增大后减少的抛物线趋势;其中,6径阶的株树最多,12径阶的株树最少。中等胸径毛竹的株数占多数,向其两端径阶的毛竹株数逐渐减少的单峰的山状曲线,直径结构近似遵从正态分布。 表2 直径结构分布 由表3可知,新竹平均胸径与土壤有机质质量分数、水解氮质量分数显著相关(P<0.05);与土壤全氮质量分数和全碳质量分数极显著相关(P<0.01),相关系数分别为0.612、0.710;毛竹平均胸径随着有机质、水解氮、全氮、全碳质量分数的增加而增加,与有机质、水解氮和全氮相比,全碳质量分数与平均胸径的拟合度更高,即土壤全碳质量分数对平均胸径的影响更显著。新竹平均高与土壤有效磷质量浓度显著相关(P<0.05),与全氮和全碳质量分数极显著相关(P<0.01),相关系数分别为0.531、0.552;新竹平均高随着有效磷质量浓度、全氮和全碳质量分数的增加而增加,其中,全氮质量分数对平均高的影响更显著。样地株数与土壤全碳质量分数呈负相关(P<0.05),相关系数为-0.435。土壤全碳质量分数是影响新竹林分因子的主要养分,对新竹平均胸径、平均高、立竹度均有显著影响,土壤碳质量分数的增加对新竹平均胸径和平均竹高有促进作用,但对新竹的株数有抑制作用。 表3 土壤养分与新竹林分因子的Pearson相关性 3.3.1 林分非空间结构因子对毛竹林分因子的影响 由图1可知,通过采伐前林分平均胸径、平均高、立竹度、年龄均匀度与采伐后恢复更新的林分因子进行线性拟合发现,新竹平均胸径与原有林分平均高极显著相关(R2=0.482 9,P<0.01),相关系数为0.709,新竹平均胸径与原有林分平均胸径、立竹度、年龄均匀度的相关性不显著;新竹平均高与原有林分平均高极显著相关(R2=0.314 8,P<0.01),与年龄均匀度显著相关(P<0.05),但与原有林分平均胸径和立竹度的相关性不显著;新竹株数与原有林分平均胸径、平均高、立竹度、年龄均匀度的相关性均不显著。 图1 林分非空间结构因子与新竹林分因子线性拟合 3.3.2 林分空间结构因子对毛竹林分因子的影响 由图2可知,构建原有林分空间结构因子角尺度和大小比数与带状采伐后恢复更新的毛竹林分因子的拟合曲线发现,原有林分角尺度和大小比数对新竹的平均胸径、平均高、株数影响不显著;随着角尺度的增加,新竹平均胸径和平均高有减少的趋势,新竹株数有增加的趋势;随着大小比数的增加,新竹平均胸径呈增加的趋势,平均高和株数呈减少的趋势。 图2 林分空间结构因子与新竹林分因子线性拟合 由表4可知,新竹平均胸径在不同坡位、不同坡向差异不明显,但新竹平均胸径陡坡(8.0 cm)大于缓坡(7.6 cm)。新竹平均高在上坡最大,平均高为9.1 m;在不同坡向差异不明显;新竹平均高陡坡(8.6 m)大于缓坡(8.3 m)。新竹密度在不同坡位由大到小顺序表现为下坡(644株·hm-2)、中坡(510株·hm-2)、上坡(397株·hm-2);新竹密度阴坡(580株·hm-2)大于阳坡(439株·hm-2);新竹株数缓坡(690株·hm-2)大于陡坡(410株·hm-2)。 表4 平均胸径、平均高和样地株数在不同地形的变化 由表5可知,坡度对带状采伐后新竹株数的恢复影响较显著,显著性接近0.05;坡位对恢复更新的新竹的平均胸径、平均高均有影响但不显著,对新竹株数的影响显著(P<0.05);坡向对毛竹平均胸径的大小有影响但不显著,对毛竹株数和平均高影响也不显著。坡向×坡度的交互作用对平均胸径的影响较为显著(P值接近0.05),其他两两作用对林分因子的作用不显著。 表5 协方差分析结果 由表6可知,土壤全碳质量分数、原有林分平均高和坡向是影响毛竹平均胸径的主要因子,解释平均胸径变异的75%;原有平均高、土壤全氮质量分数和有效磷质量浓度是影响平均高的主要因子,解释平均高变异的52.6%;土壤全碳是影响新竹株数的主要因子,解释株数变异的15.8%。 表6 新竹林分因子与土壤养分、林分结构和地形因子的多元逐步回归模型 土壤养分与林分因子的相关性分析表明,土壤有机质、水解氮、全氮、全碳以及有效磷对新竹更新均有影响。因此,在毛竹林的经营管理中应适量增加土壤有机质、全氮和磷肥的施用,降低土壤密度,更有利于植物根系发育。 原有林分平均高对新竹的平均胸径和平均高有显著影响,这是因为毛竹是克隆植物,根茎和茎秆系统呈相关性,幼龄分株和发育所需要的营养需要从较老的分株获得,母竹系统的营养状况直接影响到发笋的数量、质量以及幼竹生长[20];毛竹在春笋幼竹时期吸收养分的能力较弱,对母竹系统的依赖性较大,且春笋幼竹时期不同龄级母竹的养分含量变化程度不同,不同龄级的母竹对幼竹的贡献不一样[21]。本研究发现原有林分的毛竹年龄均匀度对新竹平均高有显著影响,这是样地内毛竹龄级差异所造成。 区域内下坡位的新竹株数最高,且随着坡位上升,株数逐渐降低;不同坡向的新竹数量表现为阴坡大于阳坡;不同坡度表现为缓坡大于陡坡。不同坡位新竹数量差异与土壤状况有关,在重力及淋洗作用下,土壤有机质随水流向下移动[22],导致下坡土壤肥沃;太阳辐射和土壤水分引起的有机分解速率是不同的,从而表现为阴坡的土壤养分状况优于阳坡[23]。研究显示不同坡位、坡向和坡度的新竹平均胸径和平均高差异不明显,这与调查时间和毛竹立竹度相关,本研究在毛竹长出当年4月进行调查,毛竹的高生长还没有停止。根据毛竹的直径分布结构可看出小胸径的株数分布较多,研究区中下坡位、阴坡和缓坡的新竹数量明显大于其他地形水平,并且根据实地调查发现下坡位、阴坡和缓坡的毛竹多为小胸径。 本研究根据采伐前的土壤养分、林分因子、地形因子等数据,以及调查带状采伐后翌年更新的新竹林分因子,分析了土壤养分、原有林分结构和地形因子对新竹平均胸径、平均高、数量的影响。结果发现,土壤养分对毛竹林分因子的生长有显著影响;采伐前的林分结构因子影响新竹的生长,其中原有林分的平均高是影响新竹平均胸径和平均高的主要林分因子;不同坡位、坡向、坡度的新竹株数存在显著差异,林分平均胸径和平均高差异不显著。

3 结果与分析
3.1 带状采伐后毛竹恢复更新现状

3.2 土壤养分对毛竹恢复更新的影响
3.3 林分结构对毛竹恢复更新的影响

3.4 地形因子对毛竹恢复更新的影响


3.5 土壤养分、林分结构和地形因子对毛竹更新的影响

4 结论与讨论
猜你喜欢
意林·少年版(2020年13期)2020-08-02 11:02:50
临床检验杂志(电子版)(2020年1期)2020-04-03 04:00:16
新农民(2019年9期)2019-02-19 14:19:13
学苑创造·B版(2015年12期)2016-06-23 09:36:36
福建农业科技(2015年3期)2015-02-27 10:20:44
福建农业科技(2015年1期)2015-02-27 10:20:39
福建农业科技(2015年1期)2015-02-27 10:20:38
小天使·一年级语数英综合(2014年6期)2014-07-22 07:15:07
家家乐·上半月(2013年7期)2013-04-29 00:44:03
家家乐·上半月(2013年6期)2013-04-29 00:44:03
