
不同经营方式的云冷杉针阔混交林土壤真菌群落结构1)
2023-03-28 01:13:58刘志宇张忠辉杨凯麟张军姜润华吴则甫王琦李文华夏富才
东北林业大学学报 2023年3期
刘志宇 张忠辉 杨凯麟 张军 姜润华 吴则甫 王琦 李文华 夏富才
(吉林省林业科学研究院,长春,130033)(北华大学)
To assess the effects of different management practices in spruce-fir forests, the Spruce-fir mixed forest in Wangqing Forestry Bureau of Jilin Province was taken as the research object, and four management methods were set up. The traditional management: the canopy density was above 0.7 after felling.Target tree management 1: felling was carried out according to target tree single plant management, and the target tree density of timber was reserved to 100 ind·ha-1. Target tree management 2: felling was carried out according to target tree single plant management, and the target tree density of timber was kept at 150 ind·ha-1. The 3 uncut pairs are also denoted as CK. Through high-throughput sequencing technology and soil physicochemical analysis, the differences of soil bacterial community structure under different management methods were compared. The results showed that: (1)There were significant differences in soil chemistry between management practices (p<0.05). (2)Logging increased the number of OTU and OTU of fungi in Spruce-fir mixed forest. (3)α diversity analysis showed that target tree management 1 had the most abundant fungal community, while traditional tree management had the highest fungal community diversity. (4)β diversity analysis showed that the difference between different management methods was significantly higher than that between groups, suggesting that logging changed the species composition of fungal community. Therefore, the dominant microbial community of soil fungi had different responses to disturbance, and the soil quality decreased due to logging. After logging, the composition of soil fungi tended to be supported by saprophytic bacteria. Soil fungal community structure showed different similarities under different management methods, and harvesting changed the fungal community structure compared to the control operation, indicating that the soil fungi did not stabilize after a short period of recovery after harvesting. In conclusion, harvesting can affect soil fungi for a short period of time, and it facilitates nutrient cycling by soil fungal communities. From the analysis of fungal abundance, community structure and similarity, target tree management 2 was closer to the control group, so target tree management 2 was superior to other management methods.
土壤中微生物是陆地生态系统中的重要组成部分,是森林生态系统中能量流动、物质循环、信息传递的重要驱动者[1],是全球生物多样性的重要组成部分[2],对陆地生态系统有着重要作用[3]。土壤真菌通过调节一些关键的生化过程来影响森林生产力,如有机质分解、养分循环、土壤肥力的维持、土壤碳的固定、植物养分吸收等[4-6]。土壤性质例如碳氮质量比、pH、速效磷质量分数等在决定土壤真菌群落的结构、多样性方面作用巨大[7-8],并且土壤理化性质又受到森林经营方式的显著影响[9-11]。
随着研究的深入,近几年的研究表明:农田地和造林土壤的优势真菌门为子囊菌门(Ascomycota)、接合菌门(Zygomycota)和担子菌门(Basidiomycota),但造林土壤中接合菌门的相对丰度较农田地大[12];森林采伐或树冠间伐将显著增加土壤异养真菌的呼吸,从而增加土壤CO2排放总量[13];对黑松林分进行间伐处理,发现其表层土壤内真菌微生物丰度和微生物群数均显著高于对照处理的同龄林分土壤[14];辽东山区日本落叶松人工林的土壤真菌差异随林龄增加而加大,并且真菌比细菌更为显著[15];在贵州茂兰喀斯特森林,阳坡的凋落物对真菌群落的影响较大,并且凋落物中的碳、磷质量分数与真菌群落有极显著的正相关性[16]。以上研究表明,经营方式通过改变林分空间结构,影响森林内的土壤环境,进而影响了土壤真菌,说明真菌对外界变化敏感。因此,研究不同经营方式下土壤真菌群落结构变化,是评价森林经营效果的重要手段。
云冷杉针阔混交林是由云杉(Piceajezoensis)和冷杉(Abiesnephrolepis)为主要树种的天然异龄混交林[17],其林分蓄积约为22亿m3,占全国总蓄积量的20%以上[18],是长白山地区重要的森林类型。云冷杉针阔混交林不仅具有重要的生态、社会及经济价值,而且是主要的用材树种,曾被大量采伐[19]。由于当时的技术和理论限制,对云冷杉针阔混交林没有及时进行科学的恢复和保护,导致现在的云冷杉林林分质量低下。如何恢复云冷杉林的结构和功能是东北林业亟需解决的问题之一[20-21]。过去的20 a,许多专家学者以近自然林经营理论为指导,以目标树经营为方法,从蓄积增长[22]、林分空间结构[23]、森林演替趋势[24]、种子雨动态[25]等多方面对云冷杉林进行研究,并取得了很多重要成果。但是研究不同经营方式对长白山地区云冷杉林土壤真菌组成影响的文章鲜有报道。所以本文根据在吉林省汪清县云冷杉林设置的永久监测样地,从采伐干扰对土壤真菌的影响角度评估不同经营方式的效果,对云冷杉针阔混交林可持续经营提供参考。
1 研究区域概况
研究区域位于吉林省汪清林业局金沟岭林场辖区内,属于云冷杉林典型过伐林区,地理坐标130°5′~130°20′E,43°17′~43°25′N[26]。林区地貌属低山丘陵,海拔在300~1 200 m,除个别陡坡在35°以上,其余坡度在5°~25°。该区是受季风影响的温带大陆性山地气候,1月份最低气温-32 ℃,7月份最高气温32 ℃,全年平均气温为4 ℃;年降水量600~700 mm,且多集中在7月份;土壤多为灰棕壤[27]。主要针叶树种为鱼鳞云杉(Piceaasperata)、臭冷杉(Abiesnephrolepis)、长白落叶松(Larixolgensis)等;主要阔叶树种为枫桦(Betulacostata)、紫椴(Tiliaamurensis)、白桦(Betulaplatyphylla)等;主要灌木为胡枝子(Lespedezabicolor)、毛榛(Corylusmandshurica)、忍冬(Lonicerajaponica)等;主要草本有蕨(Adiantumspp.)、宽叶苔草(Carexsiderosticta)、繁缕(Stellariamedia)等[28]。
2 研究方法
2.1 样地设置
2020年9月,于吉林省汪清林业局金沟岭林场28林班采用随机区组设计设置对照样地,记作CK,不进行任何措施;常规经营样地记作TM,即按照GB/T 15781—2009《森林抚育规程》的要求进行抚育采伐设计;目标树经营密度1记作M1,即按目标树单株经营进行采伐设计,用材目标树密度为100株/hm2;目标树经营密度2记作M2,即按目标树单株经营进行采伐设计,用材目标树密度为150株/hm2。采伐强度由强到弱为传统经营、目标树经营1、目标树经营2、对照经营。在每个林分中各设置3块30 m×30 m的固定样地,将每个样地分为9个小样方,每10 m设置1个水泥桩。经过随机区组后各样地特征值之间没有显著差异,进而研究不同经营方式下对土壤真菌群落的影响。各样地基本情况见表1。
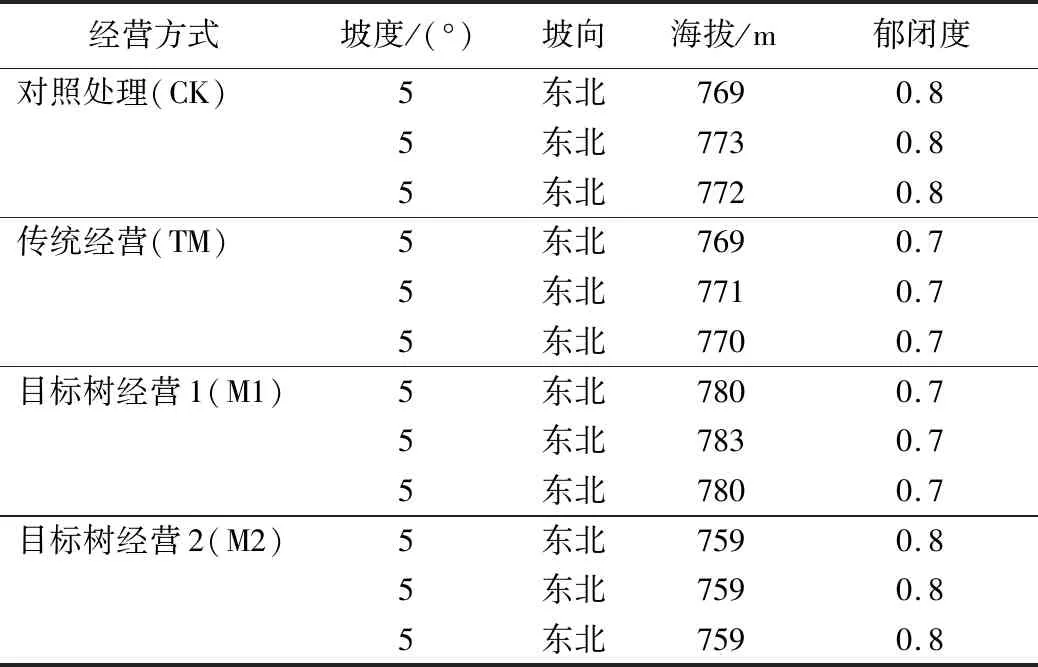
表1 不同经营方式的云冷杉针阔混交林样地基本概况
2.2 土壤样品采集
2020年9月,在每个样地内采用“S”型取样法随机设置18个采样点,剥离凋落物后利用内径为5 cm的土钻采集0~10 cm深处土壤样品,从中挑出动植物残体和石块等杂质后,混合均匀装入自封袋内,低温保存带回实验室。采集的鲜土过2 mm筛,一份用于测定土壤理化性质,另一份置于冰箱内-80 ℃保存,进行高通量测定。
2.3 土壤理化性质测定
土壤有机碳(SOC)质量分数采用重铬酸钾氧化-水合加热法测定;全氮(TN)质量分数采用K8400全自动定氮仪测定;速效氮(AN)质量分数采用碱解-扩散法测定;全钾(TK)质量分数采用氢氧化钠碱熔-火焰光度法测定;全磷(TP)质量分数采用碱熔-钼锑抗比色法测定;速效钾(AK)质量分数采用乙酸铵浸提-火焰光度法测定;速效磷(AP)质量分数采用盐酸-硫酸溶液浸提法测定;pH采用电位法测定。
2.4 高通量测序
采用MOBIO公司的强力土壤DNA提取试剂盒对土壤微生物DNA进行提取,利用琼脂糖凝胶电泳对提取后的DNA产物进行检测,将检测合格的DNA样品保存于-80 ℃备用。在95 ℃预变性2 min,随后98 ℃变性10 s,62 ℃退火30 s,68 ℃延伸30 s,共27个循环,最后68 ℃延伸10 min的程序下,利用带有barcode序列为ITS1F(5’-CTTGGTCATTTAGAGGAAGTAA-3’),ITS1R(5’-GCTGCGTTCTTCATCGATGC-3’)的特异引物,对提取样本中真菌rDNA的ITS1区域进行PCR扩增。扩增后对切胶回收的产物利用QuantiFluorTM荧光计进行定量,并等量混合纯化的扩增所得产物,连接测序接头,依据Illumina说明进行构建测序文库,使用Hiseq2500测序仪的PE250模式上机测序。
2.5 数据处理
利用R(v.3.2.1)软件vegan包,聚类分析不同营林措施下土壤真菌的群落结构的β多样性,采用PAST进行ANOSIM分析检验微生物群落结构组间差异性,通过单样本的多样性分析、α多样性反映微生物群落的丰度和多样性。
3 结果与分析
3.1 不同经营方式下土壤理化性质
由表2可知,土壤全氮质量分数对照经营与传统经营、目标树经营1差异显著(p<0.05),传统经营与目标树经营2差异显著(p<0.05),其中传统经营全氮质量分数最高,对照经营全氮质量分数最低;全钾质量分数对照经营与传统经营、目标树经营2差异显著(p<0.05),其中对照经营全钾质量分数最高,传统经营最低;速效氮质量分数对照经营与传统经营、目标树经营1、目标树经营2有显著差异(p<0.05),其中对照经营速效氮质量分数最高,目标树经营2最低;速效钾质量分数对照经营与传统经营和目标树经营2有显著差异(p<0.05),传统经营与目标树经营1、目标树经营2有显著差异(p<0.05),目标树经营1与目标树经营2有显著差异(p<0.05),其中传统经营方式速效钾质量分数最高,目标树经营2最低;土壤有机碳质量分数目标树经营1、目标树经营2分别与传统经营和对照经营有显著差异(p<0.05),其中传统经营土壤有机碳质量分数最高,目标树经营2最低;速效磷质量分数对照经营分别与传统经营、目标树经营2有显著差异(p<0.05),目标树经营1与传统经营、目标树经营2有显著差异(p<0.05),其中对照经营速效磷质量分数最高,目标树经营2最低。

表2 不同经营方式下样地土壤理化性质
3.2 不同经营方式下土壤真菌群落多样性情况
通过高通量测序分析了12个样地的混合土壤样品,真菌有效序列数925808。对所有序列进行操作分类单元(OTU)划分,按照97%相似性阈值对非重复序列(不含单序列)进行OTU聚类,12个样地共得到6670个OTU,去除掉重复OTU,得到真菌1254个OTU,12个样地文库覆盖度达到99.8%以上,可以充分反应云冷杉针阔混交林土壤真菌群落的真实情况(表3)。
根据真菌的α多样性指数(如表3)可知,传统经营的ACE指数与目标树经营1的ACE指数有显著差异(p<0.05),对照经营与目标树经营2之间没有显著差异,且ACE指数从大到小依次为:目标树经营1、目标树经营2、传统经营、对照经营,说明在以上4种经营方式之间目标树经营1的物种最丰富且各物种分配最均匀;传统经营的Chao1指数分别与目标树经营1和目标树经营2有显著差异(p<0.05),对照经营与传统经营、目标树经营1、目标树经营2没有显著差异,且Chao1指数从大到小依次为:目标树经营1、目标树经营2、对照经营、传统经营,说明目标树经营1中所含有的物种数最多,传统经营中的物种数最少;传统经营的Simpson指数与对照经营有显著差异(p<0.05),目标树经营与对照经营和传统经营没有显著差异;对照经营的Shannon指数与传统经营的Shannon指数有显著差异(p<0.05),而与目标树经营1和目标树经营2没有显著差异,且传统经营的Simpson指数、Shannon指数均为最大,说明在以上经营方式中传统经营下的真菌群落多样性最高。综上所述,随着采伐强度的增加,采伐显著改变了云冷杉林土壤真菌群落的α多样性指数。

表3 真菌群落Alpha多样性指数
3.3 不同经营方式下土壤真菌群落结构分析
使用UNITE数据库通过朴素贝叶斯分类器对特征序列进行分类学注释,可得到每个特征对应的物种分类信息,并在各水平(门、纲、目、科、属、种)统计各样品群落组成。由于是将真菌序列片段打散,进而测序,会导致一个真菌物种对应多个OTU的情况出现,所以真菌共得到9门、27纲、69目、156科、319属、418种。
在门水平上,共发现9门真菌,主要优势门为担子菌门(Basidiomycota)、子囊菌门(Ascomycota)、被孢菌门(Mortierellomycota)、隐菌门(Rozellomycota)、壶菌门(Chytridiomycota)(表4)。其中担子菌门、子囊菌门的相对丰度累计达到80%以上,说明这2个菌门具有重要的作用。
通过方差分析可知(如表4),担子菌门相对丰度传统经营显著低于对照经营(p<0.05),子囊菌门相对丰度传统经营显著高于目标树经营(p<0.05),并且目标树经营的相对丰度显著高于对照经营(p<0.05)。在门水平,壶菌门的相对丰度从大到小依次为:对照经营、传统经营、目标树经营1、目标树经营2;被孢菌门的相对丰度从大到小依次为:传统经营、目标树经营1、对照经营、目标树经营2;隐菌门的相对丰度从大到小依次为:传统经营、目标树经营2、目标树经营1、对照经营。综上所述,除壶菌门外,其他优势菌门与对照经营相比,采伐显著改变了真菌群落4个优势菌门的群落结构。

表4 不同经营方式下优势真菌相对丰度
利用相似性检验(ANOSIM,如图1)从统计学的角度分析经营措施对土壤真菌群落的影响,分析结果显示出经营措施显著影响了真菌群落的物种组成(p<0.05),即不同经营措施处理下的样地内部差异性要小于不同经营方式间的差异,非相似性检验结果表明不同经营措施改变了土壤微生物的物种组成,但无法揭示不同微生物分类单元的变化规律。

图1 不同经营措施的云冷杉针阔混交林真菌群落相似性检验
对表征云冷杉针阔混交林土壤真菌群落进行主成分分析,如图2,发现第一主成分(PC1)和第二主成分(PC2)分别解释了土壤真菌群落结构变异的24.73%和18.03%。经过目标树经营和传统经营后的样地明显与对照经营分开,对照样地在PC1的右侧,而2种目标树经营和传统经营在PC1的左侧;对照经营在PC2的上方,而2种目标树经营和传统经营在PC2的下方,说明与对照经营相比,2种目标树经营和传统经营改变了真菌的群落结构。
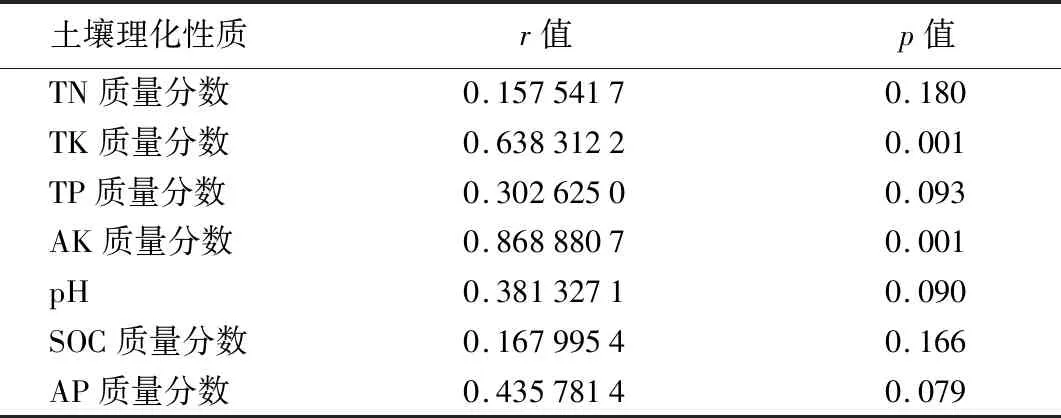
表5 真菌群落结构与土壤理化性质的相关性结果
3.4 土壤环境因子与微生物的相关性
通过Mantel分析检验云冷杉针阔混交林土壤真菌群落结构与土壤环境因子之间的相关关系(如表5),结果显示真菌群落结构与土壤TK、AK质量分数呈现显著相关(p<0.05),其中AK质量分数影响最大(r=0.868 8,p=0.001),TK质量分数对真菌群落也有显著影响(p<0.05)。
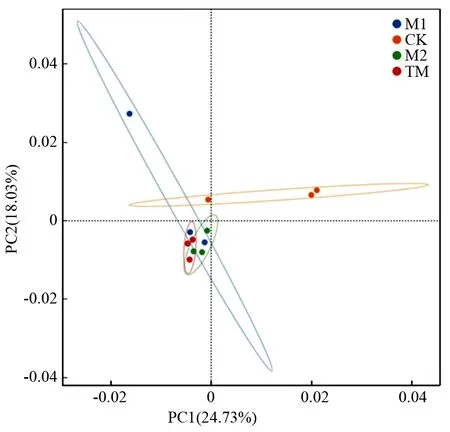
PC1和PC2分别为主成分分析第一主轴和第二主轴,括号中百分数为对应主轴的累计贡献率。
4 讨论
4.1 经营方式对土壤真菌群落结构特征的影响
本研究表明担子菌门、子囊菌门等5菌门是长白山云冷杉林的优势真菌,这些菌群也广泛存在于其他的森林土壤中,并且会时刻影响着群落结构[29-31],说明森林的物质循环是由这些真菌参与并从中发挥主导作用的。
真菌群落作为物质循环的重要组成部分,是土壤生态系统适应生境变化的重要指标[32]。大部分真菌群落对采伐是敏感的,采伐后真菌群落不会迅速恢复至最初的状态,并且真菌群落的组成会因生态系统的变化而变化[32-34]。本研究经过短期干扰,土壤真菌群落特征适应显著。经过采伐后真菌数量显著降低[33],这是因为对森林进行采伐造成的[34]。以上结果表明真菌群落的改变是由于不同的森林经营方式造成的。
研究证明不同经营方式可以改变主要微生物群落的数量。同时,微生物生境和养分条件的改变,会改变土壤理化性质及微生物群落结构的稳定性,最终影响土壤微生物菌群的变化[32,35-38]。另外,土壤微生物群落结构也会随土壤养分的变化而变化[39]。因此,微生物群落组成的改变在某种程度上反映出土壤养分的状况[40]。本研究对真菌群落种群的相对丰度差异分析表明:与对照相比,传统经营显著降低了担子菌门的相对丰度,但显著增加了子囊菌门的相对丰度;而目标树经营下的担子菌门、子囊菌门、被孢菌门在真菌的门水平上变化趋势与对照相似。这是因为干扰树对土壤养分造成显著影响。
4.2 土壤真菌群落结构的适应机制
土壤真菌群落的功能复杂,且组成多样,生境在一定程度上改变时,真菌不会彻底改变,所以使得土壤真菌有一定的抵抗力来应对采伐[41]。真菌主营寄生,多集中在腐木、倒木以及凋落物附近,并非像细菌一样随机分布,地上部分的树木被采伐,凋落物被移除或破坏致使土壤暴露在地表时对土壤真菌群落结构影响相对较大[34]。除此之外,无法避免在采伐过程中由于人为导致土壤的紧实度增加,土壤团粒间空间下降,土壤中空气流通能力减弱,形成低氧或无氧空间,使部分厌氧菌生长[34]。真菌群落对外界环境变动的抵抗能力越强,其群落结构具有相似性越高,反之相似性越低[42]。本研究中,干扰后土壤真菌群落结构变化明显,是由于针对云冷杉林进行的目标树经营的低强度采伐在一定程度上影响了土壤真菌群落结构,但没有彻底改变土壤真菌群落。土壤微生物的相似性可以作为衡量土壤抵抗力稳定的指标,这是由土壤微生物群落结构和功能所决定的[43],土壤微生物群落抵抗力越高说明其内部多样性越高[44]。
5 结论
干扰后的云冷杉林土壤真菌群落特征发生过显著变化。(1)土壤真菌的优势菌群对干扰的适应程度不同,采伐导致土壤质量下降,经过采伐后土壤真菌的组成更加趋向于营腐生菌群。(2)土壤真菌群落结构在不同经营方式下表现出不同的相似性,与对照经营相比,采伐改变了微生物的群落结构,说明采伐后经过短时间恢复,土壤真菌未趋于稳定。综上,采伐可以短时间内影响土壤真菌,并且采伐有利于土壤真菌群落进行养分循环。(3)从真菌丰度、群落结构及相似度和土壤理化性质来综合分析,目标树经营1更接近对照经营,所以近自然林经营的角度,目标树经营1优于其他经营方式。
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29 00:51:58
现代园艺(2021年23期)2021-12-01 07:47:44
食品安全导刊(2021年20期)2021-08-30 06:40:50
林业勘查设计(2020年1期)2021-01-18 02:40:48
新农业(2020年18期)2021-01-07 02:17:08
辽金历史与考古(2019年0期)2020-01-06 07:44:36
现代营销·学苑版(2016年12期)2017-01-23 13:23:53
中国广播(2016年10期)2016-11-18 13:11:11
当代经济(2016年26期)2016-06-15 20:27:13
水生生物学报(2015年1期)2015-02-28 16:01:05
