
不同生态区谷子品种表型鉴定及SSR遗传多样性分析
2023-01-18 14:39吕建珍王宏勇马建萍
核农学报 2023年3期
吕建珍 王宏勇 任 莹 马建萍 赵 凯
(山西农业大学农学院,山西 太原 030031)
谷子(Setaria italica.Beauv.)起源于我国,具有生长周期短、抗旱耐瘠薄、粮草兼用、医食同补等特点,是我国北方干旱、半干旱地区重要的特色杂粮作物[1]。按照国家谷子区域试验布局,谷子生态区划分为华北夏谷区、西北春谷早熟区、西北春谷中晚熟区和东北春谷区,目前全国谷子种植面积约150万公顷[2]。21世纪以来,全国谷子育种单位加大了新品种选育力度,谷子新品种的数量明显增多,表型性状及产量遗传改良效果均得到显著提升,为推动谷子产业发展起到了重要作用[3-4]。
种质资源的遗传多样性评价和群体结构分析对整体把握资源材料信息、了解谷子品种的遗传基础、有效利用谷子种质资源进行亲本选择、种质创新及作物改良等具有重要意义[5-6]。国内外对谷子表型性状的研究已有不少报道。王海岗等[7]对来自世界各地878份谷子核心种质的15个表型性状进行研究,发现育成品种的遗传多样性水平低于农家种,且株高和穗长均低于农家种。田伯红[8]对482份谷子地方品种和近30年育成品种的11个农艺性状进行分析,同样发现育成品种遗传多样性水平较低。相吉山等[9-10]对不同生态区谷子种质资源进行表型分析,发现不同生态区品种生育期、农艺性状和产量性状差异显著。丁银灯等[11]在新疆通过鉴定274份谷子种质资源的遗传多样性,筛选出适宜南疆复播及北疆冷凉地区种植的谷子种质。王晓娟等[12]对甘肃省谷子地方种质表型遗传多样性分析结果表明,茎长度、主穗长等性状遗传变异较大。谷子的基因组较小,约430 Mb,是继水稻、小麦、玉米等之后完成全基因组测序的禾本科重要模式作物[13-15]。目前已有一些基于全基因组序列信息,运用有效的分子标记研究谷子品种的遗传多样性的报道。Jia等[16]采用SSR标记将谷子品种分为春播型和夏播型两类。贾小平等[17]采用37对SSR标记对40份谷子品种进行遗传多样性研究,发现农家品种的生态类型与聚类群存在一致性。王珊珊等[18]利用28对SSR引物分析了中国辽西地区谷子品种的遗传差异及不同品种间的亲缘关系。国外其他学者对谷子的遗传多样性也进行了广泛研究[19-22]。通过表型和分子标记同时对参试材料进行遗传多样性分析,可以更准确地了解参试材料间的遗传关系。杨慧卿等[23]对68份分蘖型谷子进行表型和分子标记遗传多样性分析指出,除出谷率外,数量性状表现了丰富的遗传变异,并鉴定出10份优势谷子分蘖种质。丁银灯等[24]利用124份谷子种质资源证明依据表型和SSR的聚类分析均存在显著的地理特征。杨延兵等[25]对不同生态区12份骨干谷子品种进行表型和SSR水平的遗传分析,表明华北夏谷区间的品种差异小于春谷区。
新品种登记以来,我国完成登记的谷子新品种有497个[3],不同生态区谷子品种具有不同的表型特征,但基于表型和SSR标记同时对不同生态区谷子品种进行整体评价的研究相对较少。鉴于此,本研究通过对近年不同生态区谷子育成品种(系)及山西农家种进行表型和SSR标记遗传多样性及遗传结构分析,综合评价不同生态区谷子品种特点及同一生态区不同类型材料间的遗传变异水平,旨在为谷子育种亲本选择、遗传改良和品种选育提供理论依据。
1 材料与方法
1.1 试验材料
试验材料为不同生态区谷子育种单位选育的谷子新品种(系)及山西农家种共175份。夏谷区共30份,由河北省农林科学院谷子研究所15份(简称河北)和河南省安阳市农业科学院15份(简称安阳)组成;春谷早熟区21份,均来自赤峰市农业科学院(简称赤峰);春谷中晚熟区共107份,来源于3个育种单位的4种类型,其中山西农业大学谷子研究所31份(简称长治)、山西农业大学经济作物研究所22份(简称汾阳)、山西农业大学农学院54份,包含春谷育成种30份(简称太原Ⅰ)和春夏谷杂交品系24份(简称太原Ⅱ);山西农家种17份。以晋谷21号为亲本的材料23份(含晋谷21号),均为春谷中晚熟区品种;以豫谷18为亲本的材料12份(含豫谷18),均为夏谷区品种。
1.2 田间试验设计及表型鉴定
2020—2021年在山西农业大学晋中榆次东阳试验基地(37.6°N,112.7°E)种植,种植方式为每个品种(系)2行,行长3.0 m,行距36.7 cm,株距6~8 cm。试验地前茬为玉米,肥水管理及病虫害防治根据当地大田习惯实施。
参考《谷子种质资源描述规范和数据标准》[26]和《NY∕T 2425-2013植物新品种特异性、一致性和稳定性测试指南(谷子)》[27],调查9个质量性状,包括幼苗叶鞘色、花药色、叶枕、刚毛长度、刚毛色、穗型、穗颈形状、粒色和米色。并参照文献[28]对质量性状赋值,叶鞘色:1=绿,2=浅紫,3=紫;花药色:1=黄,2=白,3=褐;叶枕:1=无,2=弱,3=中,4=强;刚毛长短:1=短,2=中,3=长;刚毛色:1=绿,2=紫;穗型:1=纺锤,2=棍棒,3=筒形,4=鸡嘴;穗颈形状:2=中弯,3=弯曲;粒色:1=黄,2=白,3=灰,4=黑,5=褐,6=红;米色:1=黄,2=浅黄,3=灰绿,4=白,5=灰色;测定15个数量性状,包括分蘖数、主茎节数、主茎直径、主茎长度、倒二叶长、倒二叶宽、穗长、穗颈长、主穗直径、单穗码数、单码粒数、单穗重、穗粒重、千粒重、出谷率;观察记载整个生育期间的出苗、抽穗、开花及成熟日期,并计算不同生育期阶段日数。
1.3 SSR分子标记鉴定
1.3.1 DNA提取 谷子抽穗后取倒二叶,装入一次性密封硫酸纸袋,-80 ℃超低温冰箱保存备用。DNA提取采用天根生化科技(北京)公司的植物基因组DNA提取试剂盒。
1.3.2 PCR扩增 参照文献[29-31]选择分布在1号~9号染色体上的36对多态性引物(附表1),平均每条染色体4对,引物序列由上海生工生物技术有限公司合成。反应体系为10 µL:模板DNA 2 µL,10 mmol·L-1正、反向引物各0.4 µL,去离子水2.2 µL,PCR扩增混合液5 µL。PCR反应程序:95 ℃预变性5 min;95 ℃变性30 s,退火温度下30 s,72 ℃延伸1 min,共34个循环;72 ℃终延伸10 min。
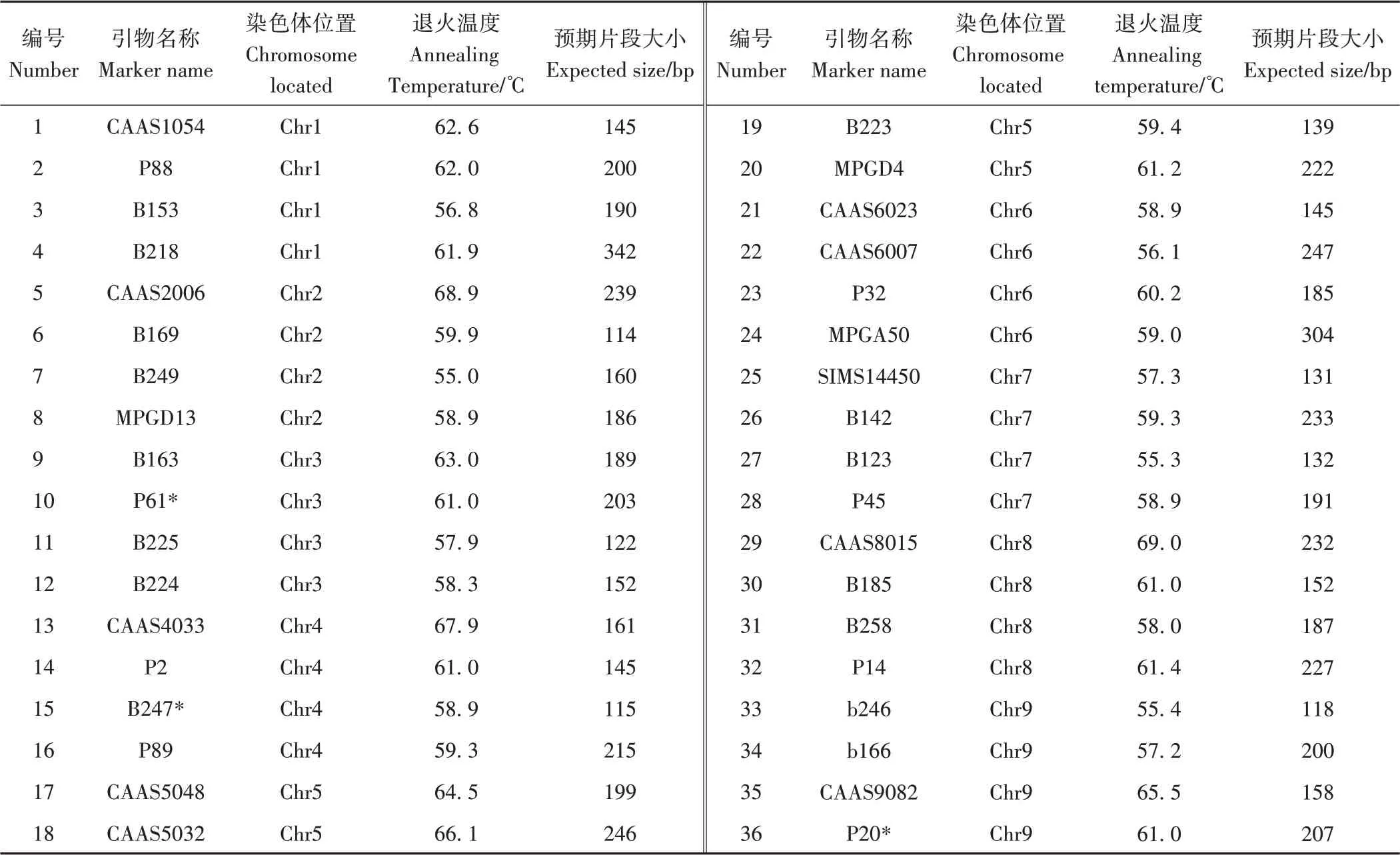
附表1 所用引物详细信息Table S1 The SSR primers used in this study
1.3.3 凝胶电泳与银染显影 试验扩增产物用8%聚丙烯酰胺凝胶电泳法进行分离。电泳缓冲液用0.5×Tris-boric acid-EDTA(TBE),上样量为2 µL,采用JY-CX3B电泳槽和JY3000E电泳仪(北京君意东方电泳设备有限公司)在255 V恒定电压下进行电泳,待Marker条带到胶板下部约2~3 cm处停止电泳,然后采用银染法显色,拍照,记录结果。
1.4 数据分析
1.4.1 表型性状数据 利用Excel 2007和DPS 7.05进行性状频次分布、平均值、标准差和多样性指数的计算,并对不同类型材料进行方差分析和Duncan新复极差法多重比较。表型遗传多样性指数(Shannon-Wiener diversity index,H´)[28]计算公式为:H´=-∑ PilnPi,式中,Pi为某一性状第i级别内材料份数占总份数的百分比,ln为自然对数。表型聚类图利用R语言绘制。
1.4.2 SSR标记数据 根据SSR扩增产物的电泳结果,参照DNA ladder maker记录不同条带类型,应用Popgene32软件统计引物的基础多样性遗传参数及Shannon指数,使用Powermarker 3.25[32]软件计算每个位点多态性信息含量(polymorphism information content,PIC)。采用Structure2.3.4[33]软件分析群体遗传结构,估计最佳群体组群数K,其取值范围为2~7,3次重复;参数iterationgs设为10 000,burn-in period设为100 000,每个K值重复运行20次。根据InP(D)计算△K,并依据△K值选择合适的K值,对不同类型材料进行聚类分析。
2 结果与分析
2.1 表型性状多样性
参试材料质量性状平均遗传多样性指数为0.663 9,其中粒色遗传多样性指数最高(1.078 7),穗颈形状最低(0.149 3)。数量性状平均遗传多样性指数为2.004 3,不同生育期阶段平均遗传多样性指数为1.659 4。由表1可知,数量性状单穗码数遗传多样性指数最高(2.082 8),分蘖最低,仅为1.582 8;产量因子单穗重的遗传多样性指数为2.064 7。不同生育阶段中开花至成熟日数遗传多样性指数最高(2.012 5),播种至出苗日数最低,为0.782 1。
由表1还可知,不同数量性状指标单码粒数变幅最大(53.40~221.40),标准差最大(31.73),其次为株高(77~157 cm),千粒重变幅最小(2.01~3.72 g),标准差最低。生育期各阶段,全生育期变幅最大(97~137 d),标准差最大(10.29),播种至出苗日数变幅最小(6~10 d),标准差最小,仅为0.63。
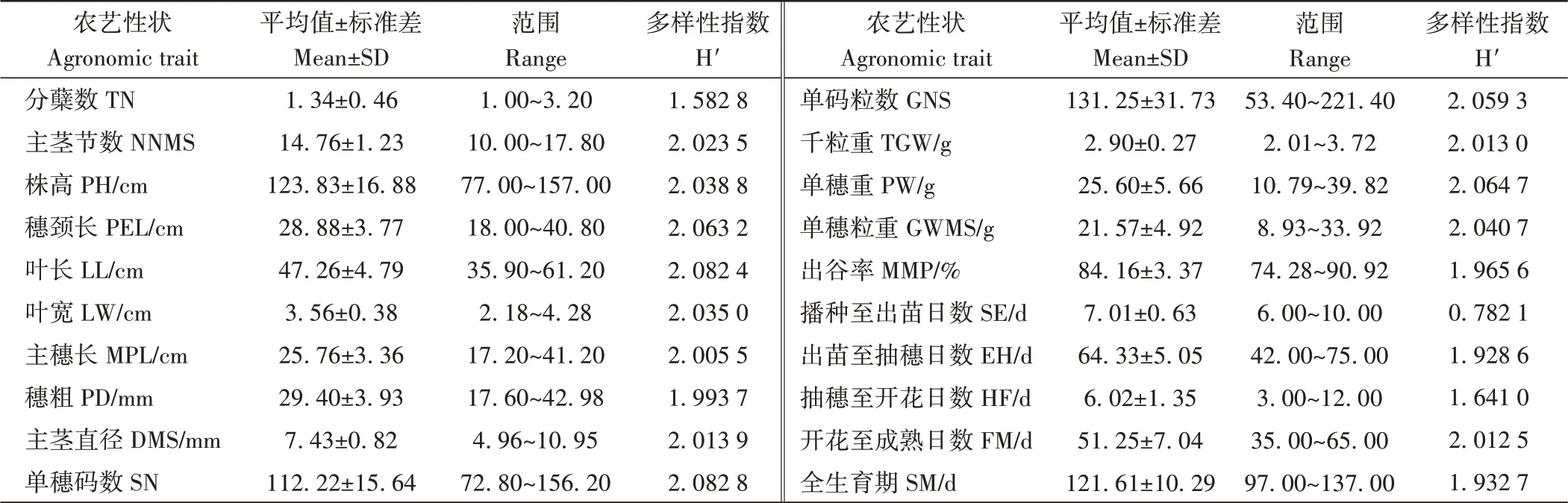
表1 参试材料数量性状及生育期遗传多样性分析Table 1 Variation of quantitative traits in tested accessions
2.2 不同生态区育成种表型性状差异
2.2.1 质量性状 夏谷区幼苗叶鞘以绿色(93.33%)为主,叶枕各类型比例与叶鞘色一致;白色花药黄色花药分别占53.33%和46.67%;刚毛为短刚毛,绿色和紫色各占50%;穗型以纺锤形为主(80%);穗颈强弯,粒色以黄色(86.67%)为主,少数褐色(10%);米色分黄(13.33%)或浅黄(86.67%)两类。春谷早熟区叶鞘也以绿色(85.71%)为主,花药色以黄色和白色为主,但褐色花药比例(19.05%)高于其他生态区;刚毛以绿色(76.19%)为主,中、长刚毛比例高于夏谷区和春谷中晚熟区;穗型也以纺锤形(80.95%)为主;粒色有4种,以黄色(38.10%)和白色(33.33%)为主,红色籽粒比例(23.81%)高于其他生态区;米色与夏谷区一致,黄色和浅黄色分别为14.29%和85.71%。春谷中晚熟区浅紫色(17.76%)和紫色(10.28%)叶鞘均高于夏谷区和春谷早熟区;刚毛以绿色、短刚毛为主;穗型有4个类型,其中纺锤形穗型占42.99%;粒色以黄色(54.21%)和白色(42.06%)为主,少数黑色(3.74%);米色5种类型均有分布,其中黄色米比例(23.36%)高于其他生态区。由表2可知,不同生态类型遗传多样性指数中,春谷中晚熟区叶鞘色、叶枕、穗型及米色4个性状遗传多样性指数最高;春谷早熟区花药色、刚毛长、穗颈形状及粒色遗传多样性指数最高;夏谷区仅刚毛色遗传多样性指数较高。春谷中晚熟区9个质量性状平均遗传多样性指数最高(0.669 2),其次为春谷早熟区(0.654 9),夏谷区最低(0.373 5)。
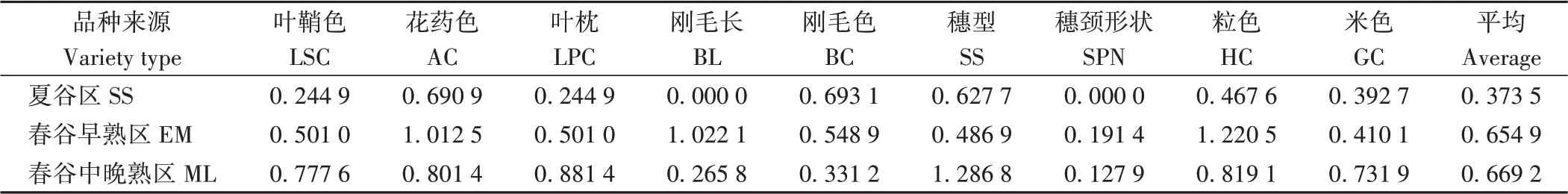
表2 不同生态区品种质量性状遗传多样性指数Table 2 The qualitative traits and the genetic diversity index of different ecological regions of tested varieties
2.2.2 数量性状 由表3可知,春谷中晚熟区品种13个数量性状遗传多样性指数最高,平均遗传多样性指数(1.967 6)高于夏谷区(1.884 2)和春谷早熟区(1.878 1)。春谷早熟区分蘖数遗传多样性指数最高,夏谷区株高遗传多样指数最高。夏谷区品种主穗长、主茎直径、单穗码数及出谷率4个性状指标最高,其他居中;其中主茎直径极显著高于春谷早熟区,但其他3个性状指标不同生态区间差异未达显著水平。春谷中晚熟区品种主茎节数、株高、穂颈长、叶长、叶宽、穗粗、单码粒数、千粒重、单穗重及单穗粒重10个性状指标均最高;其中株高、穗颈长及叶宽极显著高于其他生态区,主茎节数、叶长、千粒重、单穗重和单穗粒重极显著高于春谷早熟区,单码粒数显著高于夏谷区,不同生态区间穗粗差异未达显著水平。春谷早熟区分蘖数最高,且不同生态区间差异达极显著水平。
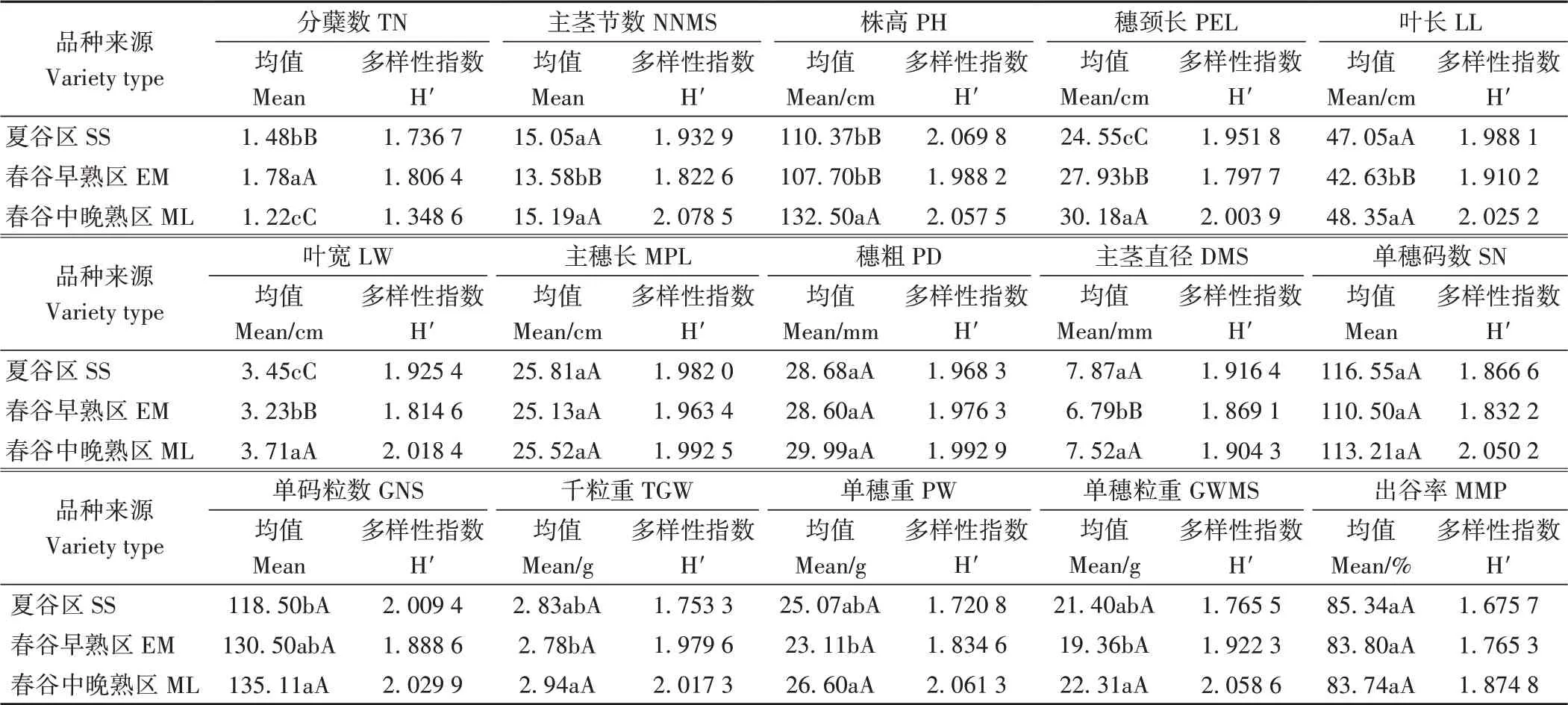
表3 不同生态区品种数量性状显著性比较及遗传多样性指数Table 3 Genetic diversity and significant difference of quantitative traits of different ecological regions of tested varieties
2.2.3 生育期 由表4可知,春谷中晚熟区品种平均遗传多样性指数最高(1.660 5),春谷早熟区次之(1.488 9),夏谷区最低(1.440 7)。不同生态区的开花至成熟阶段遗传多样性指数均最高,播种至出苗最低。在播种至出苗和抽穗至开花两个生育阶段,不同生态区间差异未达显著水平;春谷早熟区品种出苗至抽穗日数最低(57.90 d),极显著低于其他生态区,春谷中晚熟区最高(66.18 d),极显著高于春谷早熟区;春谷中晚熟区品种开花至成熟日数最高(55.35 d),极显著高于其他生态区;春谷中晚熟区品种全生育期最长(127.56 d),春谷早熟区最低(107.48 d),不同生态区间差异达极显著水平。
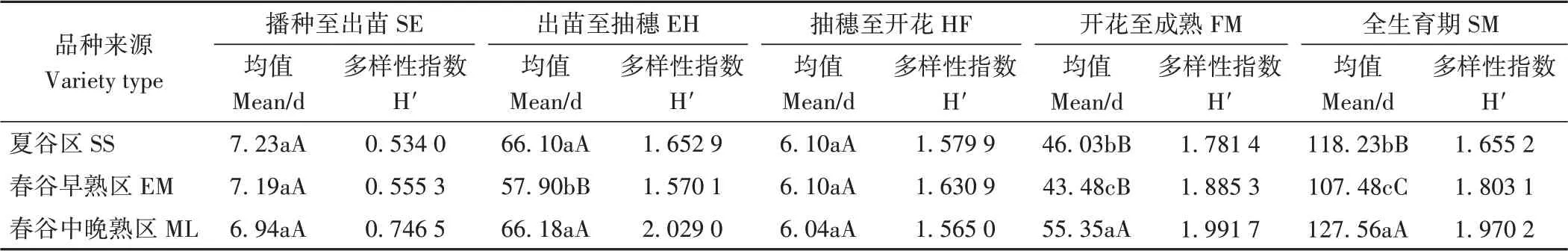
表4 不同生态区品种生育期显著性比较及遗传多样性指数Table 4 Genetic diversity and significant difference of growth period of different ecological regions of tested varieties
2.3 春谷中晚熟区不同类型材料表型性状差异
2.3.1 质量性状 太原Ⅰ和长治品种紫色叶鞘比例较高,分别为23.33%和22.58%。长治和汾阳品种花药色以黄色为主,太原Ⅰ黄色和白色各占一半,太原Ⅱ以白色为主。长治和汾阳品种穗型丰富多样,各类型比例均衡,太原Ⅰ和太原Ⅱ以纺锤和筒形穗为主。长治和汾阳粒色以白色为主,太原Ⅰ和太原Ⅱ以黄色为主。汾阳和长治小米商品性好,标记为1(黄)的品种比例分别为47.62%和32.26%。
不同类型材料遗传多样性比较发现(表5),太原Ⅰ叶鞘色、叶枕2个性状遗传多样性指数最高;汾阳花药色遗传多样性指数最高;各类型材料刚毛长度、颜色和穗颈形状遗传多样性指数均较低;穗型遗传多样性指数均较高,汾阳最高(1.294 5);长治粒色遗传多样性指数最高(0.703 7),汾阳米色遗传多样性指数最高(0.845 5)。不同类型材料质量性状平均遗传多样性指数排序为:汾阳>太原Ⅰ>长治>太原Ⅱ。
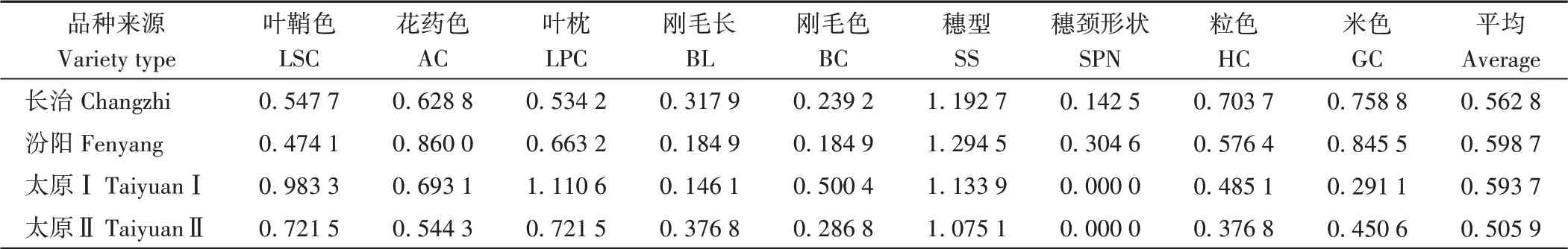
表5 不同类型材料质量性状遗传多样性指数Table 5 The qualitative traits and the genetic diversity index of tested varieties
2.3.2 数量性状 由表6可知,长治品种主茎节数、主穗长及单码粒数3个性状测试指标最低,分蘖数、穗粗及单穗码数3个性状指标最高,其中主穗长极显著低于太原Ⅱ,穗粗极显著高于太原Ⅰ。汾阳品种叶长、单穗码数、单穗粒重及出谷率4个性状指标最低,主茎节数、株高、叶宽及千粒重4个性状指标最高;其中叶长极显著低于太原Ⅱ,出谷率显著低于长治,极显著低于太原Ⅰ和太原Ⅱ,株高极显著高于太原Ⅱ,显著高于长治。太原Ⅰ分蘖数、叶宽、穗粗、主茎直径及千粒重5个性状指标最低,穂颈长、单穗重及单穗粒重3个性状指标最高;其中分蘖数极显著低于其他类型,穗粗显著或极显著低于其他类型,主茎直径与太原Ⅱ间差异显著,穂颈长显著或极显著高于其他类型。太原Ⅱ株高、穂颈长及单穗重3个性状指标最低,叶长、主穗长、主茎直径、单码粒数及出谷率5个性状指标最高。主茎节数、叶宽、单穗码数、单码粒数、千粒重、单穗重和单穗粒重7个性状指标不同类型间差异未达显著水平。
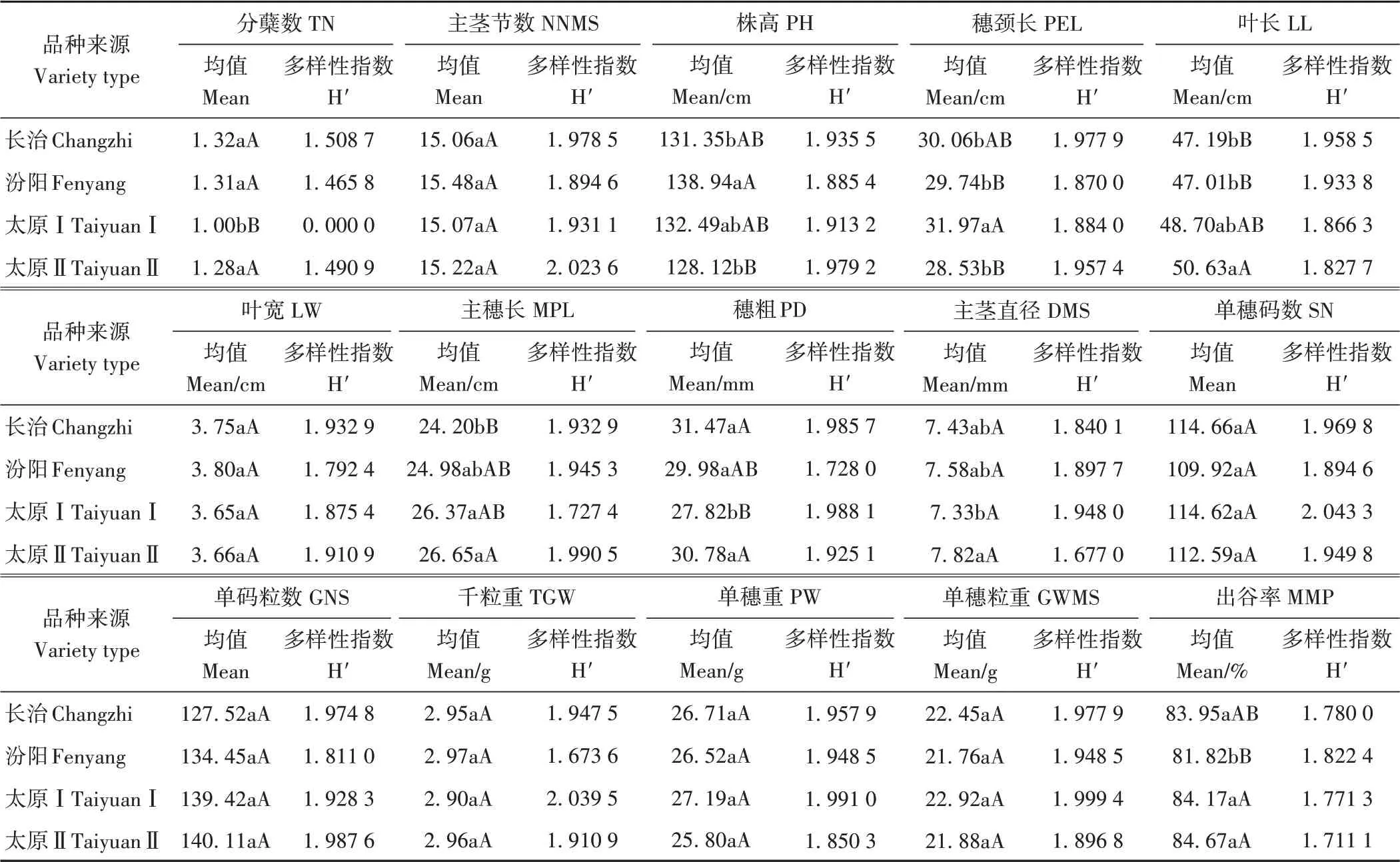
表6 不同类型材料数量性状显著性比较及遗传多样性指数Table 6 Genetic diversity and significant difference of quantitative traits of different groups of tested varieties
对不同类型材料遗传多样性进行比较发现,长治分蘖数、穗颈长、叶长、叶宽及穗粗5个性状遗传多样性指数最高,其他性状居中。汾阳出谷率遗传多样性指数最高,主茎节数、株高、穂颈长、叶宽、穗粗、单穗码数、单码粒数及千粒重8个性状最低。太原Ⅰ主茎直径、单穗码数、千粒重、单穗重及单穗粒重5个性状遗传多样性指数最高,分蘖数和主穗长2个性状最低。太原Ⅱ主茎节数、株高、主穗长及单码粒数4个性状遗传多样指数最高,叶长、主茎直径、单穗重、单穗粒重及出谷率5个性状最低。平均遗传多样性指数排序为:长治>太原Ⅱ>汾阳>太原。
2.3.3 生育期 由表7可知,太原Ⅰ出苗最快,播种至出苗日数极显著短于其他类型,遗传多样性指数变幅为0.286 8(太原Ⅱ)~0.684 2(太原Ⅰ)。出苗至抽穗和抽穗至开花两个生育阶段,不同类型间差异均未达显著水平,太原Ⅰ品种出苗至抽穗遗传多样性指数最高(1.836 8),太原Ⅱ品种抽穗至开花遗传多样性指数最高(1.592 7)。开花至成熟和全生育期阶段,不同类型间仅汾阳和太原Ⅰ品种间未达显著水平,其他类型间差异均达极显著水平,长治品种开花至成熟日数和全生育期均最长;开花至成熟遗传多样性指数变幅为1.696 4(汾阳)~1.935 6(太原Ⅱ),全生育期变幅为1.420 9(太原Ⅰ)~1.893 3(太原Ⅱ)。各阶段平均遗传多样性指数排序为:太原Ⅱ>长治>太原Ⅰ>汾阳。
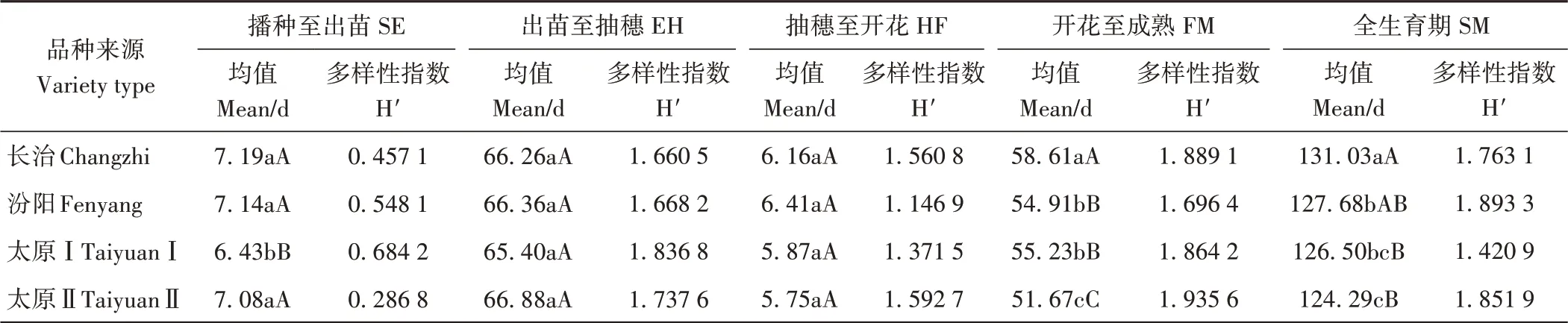
表7 不同类型材料生育期显著性比较及遗传多样性指数Table 7 Genetic diversity and significant difference of growth period of different groups of tested varieties
2.4 SSR标记遗传多样性
2.4.1 参试材料SSR多态性 利用36对SSR引物,在175份谷子材料中共检测到401个等位基因(表8),平均每对引物11.14个,变化范围为5~19个;平均有效等位基因数为6.665 3,其中B142检测到的等位基因数和有效等位基因数最多,其次为B225,CAAS6023检测到的等位基因数和有效等位基因数最少;平均每条染色体46个,第4条最多;频率<5%的稀有等位基因达160个,占39.9%;有些等位基因仅存在于某一个材料中,即特异等位基因,19个SSR位点共检测出32个特异等位基因,变幅为1~4。36个SSR位点Shannon指数(I)变幅为1.213 4(CAAS6023)~2.603 2(B142),平均为2.017 2,其中21对引物Shannon指数达2.0以上。多态性信息含量变化范围为0.576 1(CAAS6023)~0.907 3(B142),其中PIC值0.8以上的引物共24对,平均PIC值为0.810 6。
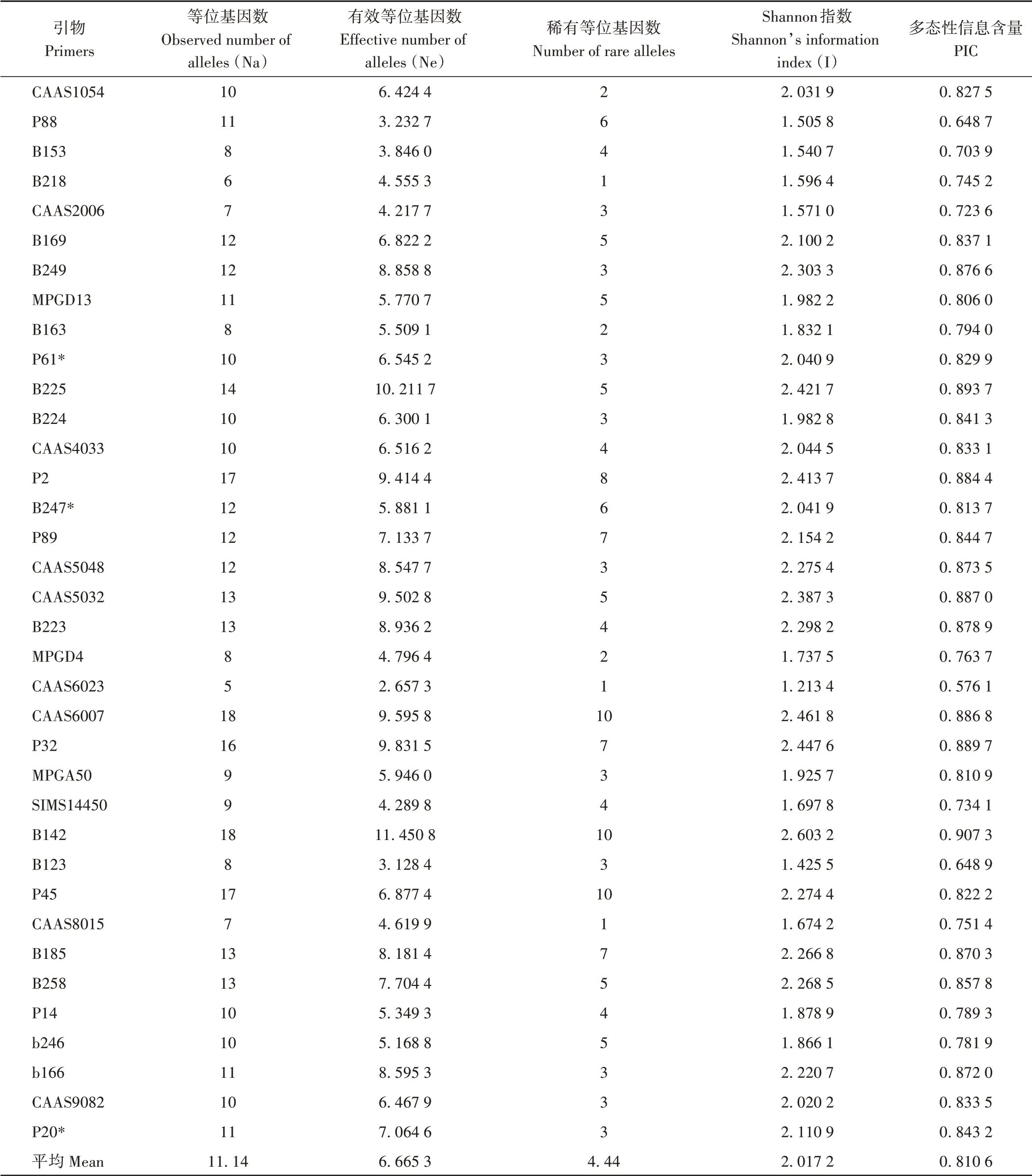
表8 36对SSR分子标记在175份谷子种质检测到的遗传变异参数Table 8 Genetic variation parameter detected by 36 SSR markers in 175 foxtail millet accession
2.4.2 不同类型材料SSR遗传变异 如表9所示,36对引物在春谷中晚熟区检测的等位基因、有效等位基因最多,Shannon指数也最高,春谷早熟区次之,夏谷区最低。春谷中晚熟区不同类型材料中,长治检测的等位基因数最多,但汾阳检测的有效等位基因数最多,Shannon指数也最高;太原育成种(太原Ⅰ)和春夏谷杂交品系(太原Ⅱ)获得等位基因数一样,但太原Ⅱ的有效等位基因数和Shannon指数较高。
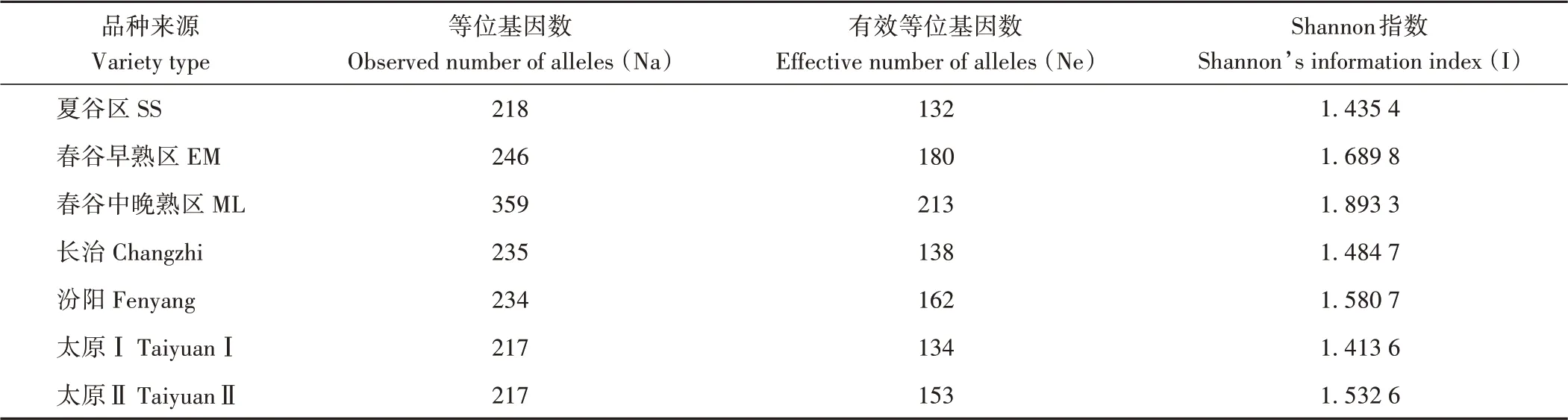
表9 36对SSR标记在不同类型材料中的遗传多样性参数及显著性分析Table 9 Polymorphism of 36 SSRs among eight different accessions
2.5 参试材料表型聚类及SSR遗传结构分析
2.5.1 基于表型的聚类 基于表型的聚类结果显示(图1),参试材料划分为4个类群(Ⅰ、Ⅱ、Ⅲ、Ⅳ),类群Ⅰ含71份材料,以高秆、大穗春谷中晚熟谷子品种为主,少数为夏谷品种,平均生育期128 d。类群Ⅱ含32份材料,夏谷区、春谷早熟区及山西农家种比例较高,矮秆、穗小,叶片短、窄;该类品种生育期短,平均113 d。类群Ⅲ含57份材料,包括13份春谷早熟区材料,16份夏谷区材料,16份春谷中晚熟区材料,5份山西农家种,综合表型性状指标居中,平均生育期为117 d。类群Ⅳ材料数量最少,仅15份,主要为春谷中晚熟品种(12份),平均生育期125 d,该类群平均穗粗、码粒数、单穗重和单穗粒重最高,穗颈最长,与其他3个类群差异达显著水平。23份以晋谷21号为亲本的品种在类群Ⅰ、类群Ⅱ和类群Ⅲ均有分布,其中类群Ⅰ含73.9%;12份以豫谷18为亲本的品种分布在类群Ⅱ和类群Ⅲ,其中类群Ⅲ含53.8%。
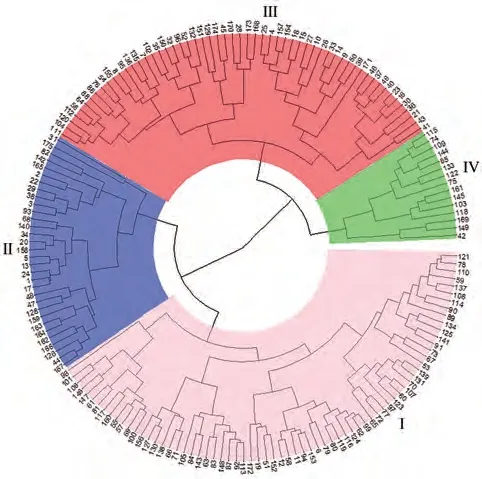
图1 175份参试材料基于29个表型性状的聚类图Fig.1 Cluster dendrogram of 175 foxtail millet accessions based on 29 agronomic traits
2.5.2 遗传结构分析 参照Evanno等[34]的方法,当K=4时,ΔK值最大。175份材料划分为4个类群(图2),类群Ⅰ含64份材料,由45份春谷中晚熟区材料(长治9份、太原Ⅰ24份、太原Ⅱ12份)、12份夏谷区材料(河北5份、安阳7份)、1份春谷早熟区及6份山西农家种组成。类群Ⅱ含32份材料,由17份春谷中晚熟区材料(汾阳17份)和15份夏谷区材料(河北7份、安阳8份)组成。类群Ⅲ含47份材料,19份春谷早熟区材料、18份春谷中晚熟区材料(太原Ⅱ11份、长治2份、汾阳5份)和7份山西农家种,夏谷区仅3份。类群Ⅳ含32份材料,以春谷中晚熟区育成种(长治、太原Ⅰ)及山西农家种为主。
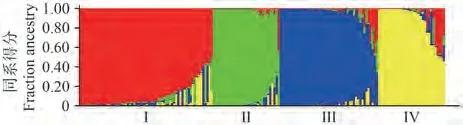
图2 基于SSR标记数据的群体遗传结构Fig.2 Population genetic structure of eight groups based SSR marker
以晋谷21号为亲本的材料分散于4个类群,类群Ⅰ含7份(南1070、M测07、SM4010、长分1号、长分2号、长谷4号及长农40号);类群Ⅱ含6份(晋汾106、晋汾107、晋汾109、晋汾113、晋汾57及汾选8号);类群Ⅲ含6份,为汾阳早期选育的5个谷子品种(晋谷21号、晋谷29号、晋谷40号、晋谷54号、晋谷57号)和南1042;类群Ⅳ含4份(晋谷42号、太选谷23、长农41号、C21)。以豫谷18号为亲本的材料分布在3个类群,类群Ⅰ含5份(豫谷38、豫谷41、安16h-8334、安17h-81611、15h-8306),类群Ⅱ含6份(冀谷45、冀谷46、豫谷18、豫谷31、豫谷32、豫谷35),类群Ⅲ仅1份(冀谷168)。
3 讨论
3.1 参试材料表型和SSR标记遗传多样性
表型性状和SSR标记是作物遗传多样性分析中常用的方法。SSR标记反映DNA水平上的差异,不易受环境影响,表型性状则是基因和环境互作的结果,易受生态条件影响。本研究中数量性状遗传多样性指数最高,质量性状最低,其中数量性状单穗码数遗传多样性指数最高(2.082 8),产量因子单穗重遗传多样性指数为2.064 7,高于王海岗等[7]报道的1.840和丁银灯等[24]报道的1.966;生育期开花至成熟阶段指数最高(2.012 5),质量性状粒色最高(1.078 7)。36对SSR引物共检测到401个等位基因,平均为11.14个,高于贾小平等[17]报道的6.16个、李剑峰等[35]报道的6个及郝晓芬等[36]报道的6.6个等位基因,但低于朱学海等[37]报道的14.5个等位基因和Wang等[29]报道的20.9个等位基因。引物B142、B225检测到的等位基因数、有效等位基因数较多,且Shannon指数和多态性信息含量(PIC)最高,为谷子遗传多样性评价的理想SSR标记。造成这种现象的原因,一方面可能是由于研究材料的差异,因为等位基因的检测效率受分析样本的影响;另一方面,可能与研究选取标记的数量和多态性水平有关。朱学海等[37]、Wang等[29]及本研究参试材料类型丰富,所用引物多态性水平高,所以平均检测到的等位基因数较多。
3.2 不同类型品种表型性状差异
谷子品种具有较强的区域适应性,不同生态区谷子品种具有不同的特征。本研究中,3个生态区植株均以幼苗叶鞘绿色,黄谷黄米,纺锤形穗的表型性状为主,但不同生态区表型性状各类型比例具有差异,其中春谷早熟区品种褐色花药比例较高,春谷中晚熟区紫色、浅紫色叶鞘、白色籽粒和黄米比例较高。夏谷区品种主穗长等4个性状指标最高,春谷中晚熟区品种主茎节数等10个性状指标最高;春谷早熟区仅分蘖数最高。春谷中晚熟区全生育期最长,品种间差异最大,春谷早熟区全生育期最短。同一生态区不同类型材料表型性状差异较低,主茎节数等7个性状指标差异未达显著水平。长治穗粗最高;汾阳株高最高,出谷率显著最低;太原Ⅰ分蘖数极显著最少,穂颈长显著最长,穗粗显著最低;太原Ⅱ叶长和主穗最长。太原Ⅰ和长治紫色叶鞘比例较高,汾阳和长治白色籽粒比例高,米色商品性好。优质和高产是品种选育的主要目标,且单穗重、单穗粒重是产量的主要构成因子[37],本研究不同生态区品种单穗重和单穗粒重均有显著提升,表明各育种单位在品种选育过程中均重视了产量因子的提升,这同张艾英等[38-39]、李志江等[40]及张婷等[41]对西北春谷中晚熟区、早熟区、东北早熟区和夏谷区谷子品种的分析结果一致。
3.3 不同类型材料遗传多样性分析
本研究发现,不同生态区基于表型和SSR标记的多样性水平一致,春谷中晚熟区平均遗传多样性指数最高,春谷早熟区次之,夏谷区最低,该结果可能与春谷中晚熟区参试材料数量多有关。不同生态区开花至成熟遗传多样性指数最高。同一生态区内不同类型材料基于不同表型和SSR标记的遗传多样性水平具有一定差异,这与徐福荣等[42]对云南省水稻品种遗传多样性的研究结果相似。汾阳在质量性状和SSR标记遗传多样性比较中最高,长治数量性状遗传多样性指数最高,太原Ⅱ遗传背景复杂,生育期遗传多样性指数最高。
3.4 基于表型和分子标记的群体结构分析
本研究发现,基于表型的聚类结果与基于SSR标记的群体结构具有一定差异,这与丁银灯等[24]的研究结果不一致。基于表型的聚类划分为两个早熟、两个中晚熟共4个类群;具有同一亲本(晋谷21号或豫谷18)的材料分布较集中,聚类结果同品种的地理来源和亲本组成具有一定的相关性。SSR标记的遗传结构分类结果同品种来源具有一定的关联,但同一亲本的材料分布较分散。这表明全国范围的谷子育种交流频繁,虽然不同生态区品种表型具有区域性,但其遗传组成正在逐步打破生态区的界限划分。晋谷21号和豫谷18是我国春谷和夏谷优质谷子育种的里程碑,控制晋谷21号和豫谷18优良性状的相关基因序列可能存在于各生态区品种中,同时群体结构也与选择的标记、标记数量、覆盖水平有关。
4 结论
不同生态区谷子品种具有不同的特征。春谷中晚熟区品种10个性状指标最高,株高、穂颈长和叶宽极显著高于其他生态区。春谷中晚熟区品种全生育期最长,春谷早熟区最短。春谷中晚熟同一生态区内,不同类型材料7个性状指标差异未达显著水平,基于表型和SSR标记的多样性水平具有一定差异。基于表型和SSR标记的聚类结果和遗传结构与品种来源均具有一定关联,但与品种亲本的相关性不同。具有同一亲本的材料在基于表型的聚类中分布较集中,但在分子标记遗传结构中分布较分散。
猜你喜欢
青海湖(2022年9期)2022-02-12
当代陕西(2021年21期)2022-01-19
山西农业科学(2021年1期)2021-01-18
文苑(2020年11期)2021-01-04
今日农业(2020年17期)2020-12-15
山西农业科学(2020年9期)2020-09-14
江西建材(2018年4期)2018-04-10
中国蔬菜(2015年9期)2015-12-21
现代农业(2015年5期)2015-02-28
中国烟草学报(2012年4期)2012-04-09
