
小麦穗型突变体表型及其转录组分析
2023-01-18 14:39陈士强刘建凤王建华范德佳何震天
核农学报 2023年3期
张 容 陈士强 刘建凤 王建华 范德佳 韩 燕 何震天
(江苏里下河地区农业科学研究所,江苏 扬州 225007)
小麦(Triticum aestivumL.)是世界上重要的粮食作物之一,提供人类约20%的食物能量。小麦穗部性状由穗长、小穗数、穗粒数和穗紧密度等性状综合决定[1],控制这些性状的主要基因通常在数量性状位点(quantitative trait loci, QTL)水平上呈现为多效应或连锁现象[2]。通过研究小麦穗部发育异常突变体,有利于促进穗部性状遗传机理的解析[3]。宋全昊等[4]在育种材料中发现了一个穗部发育突变体sda1,并将突变基因SDA1定位于6B染色体,推测该基因是利用植株中同化产物的关键基因,同时影响花器官发育和抽穗期;周丽敏等[5]通过质谱分析表明TaSDA1基因具有多效性;Faris等[6]对野生小麦的紧凑穗基因进行分析,发现穗长和每穗小穗数是由不同基因控制;顾晶晶[7]研究穗发育障碍突变体sms1,推测位于6B染色体上SPT5-like是sms1的候选基因;杜启迪等[8]经甲基磺酸乙酯(ethyl methyl sulfonate,EMS)诱变获得了表型稳定的顶端小穗退化突变体asd1,将目标突变基因定位在7A染色体短臂上,推断该区段存在一个新的控制小麦花器官发育及穗部形态发育的重要基因。由上述前人研究可以看出,小麦穗部性状突变类型丰富,控制这些突变性状的基因位点也各不相同。鉴定出更多的小麦穗型突变体,有利于进一步挖掘新的基因位点,继而促进育种应用。
小麦穗部穗轴节缩短影响穗长和穗密度,同时影响小麦产量及抗性[9]。挖掘小麦穗轴节长短调控基因,阐述穗部发育异常的分子机制有助于选育穗部性状优良的高产小麦品种[10],提升小麦产量、品质及抗性[11]。因此,本研究利用60Co-γ射线诱变处理,获得一个穗轴节和穗下节明显缩短的稳定穗型突变体sui1,并对其进行表型鉴定,利用转录组测序技术对不同样品间的差异表达基因进行功能注释、富集分析和关键功能基因的挖掘,以期为小麦穗型突变体的分子机理研究提供理论依据。
1 材料与方法
1.1 试验材料
2019—2020年以高产小麦品种扬辐麦4号(YFM4)作为野生型(wild-type, WT)对照,以突变体sui1为供试材料,按照25 cm的行距及10 cm的株距种植于江苏里下河地区农业科学研究所万福基地试验田,试验田前茬种植水稻,水肥管理同大田。sui1是由江苏里下河地区农业科学研究所核技术应用研究室通过辐射诱变创制的穗型突变体。300 Gy60Co-γ射线辐射诱变YFM4小麦干种子,密播控分蘖种植M1代,随机收取1 000个单穗,进行300 Gy60Co-γ射线二次辐照后种植成穗行。穗行中出现株高略矮、穗型极短突变体。单株收获,经多代自交,获得稳定穗型发育突变体,命名为sui1。
1.2 试验方法
1.2.1 表型性状调查 从小花原基分化期到灌浆期,分别于拔节前期、拔节期、孕穗期、花后20 d取sui1和YFM4的幼穗,进行解剖后利用PhenixXTL-165系列体式显微镜(凤凰,江西)观察穗部发育情况。成熟期调查株高、穗长、结实小穗数和单穗粒数等主要农艺性状,统计分析sui1与YFM4各个农艺性状之间的差异,采用独立样本t检验分析数据的差异显著性。抽穗期田间抛洒含赤霉病菌麦粒诱导自然发病,用于鉴定sui1和YFM4赤霉病发病情况。
1.2.2 转录组测序分析 分别于孕穗期(T1)、灌浆期(T2)取样,T1取穗轴节和穗下节,T2取穗轴节,2个生物学重复。取样后迅速放到液氮中,然后移至-80 ℃冰箱中备用。
采用Trizol法分别提取突变体与野生型的总RNA。采用NanoDrop 2000超微量分光光度计(赛默飞,美国)测定RNA浓度和纯度,采用Agilent 2100生物分析仪(安捷伦,美国)分析评估RNA完整性。样品检测合格后,送往北京百迈客公司进行转录组测序(RNA-Seq)。
利用BioMag-Oligo(dT)的磁珠富集纯化mRNA;在富集到的mRNA中加入Fragmentation Buffer,将mRNA随机打断;以打断的mRNA为模板,用六碱基随机引物合成第一条cDNA链(由北京百迈客公司合成),然后加入缓冲液、dNTPs、RNase H和DNA polymerase I合成第二条cDNA链,利用AMPure XP beads纯化cDNA;对纯化的双链cDNA再进行末端修复、加A尾并连接测序接头,然后用AMPure XP beads进行目的片段(300~400 bp)选择;最后通过PCR富集得到cDNA文库,由北京百迈客公司完成。文库构建完成后,用Illumina高通量测序平台对cDNA文库进行测序。
测序的原始数据经过滤得到干净数据(clean data),利用HISAT2软件对小麦参考基因组进行序列的比对,利用StringTie对比对上的读长(reads)进行组装和定量;使用DESeq2软件对突变体和野生型间差异表达基因(differentially expressed genes, DEGs)进行筛选,在百迈客云平台(www.biocloud.net)对DEGs做基因本体论(gene ontology, GO)功能 分类 和Kyoto Encyclopedia of Genes and Genomes(KEGG)Pathway富集分析。
2 结果与分析
2.1 穗型突变体表型与生物学特征
与野生型YFM4相比,突变体sui1分蘖数增加,孕穗期至成熟期株高降低(图1-A),穗部排列紧密(图1-B),穗轴节及穗下节长度缩短(图1-C、D),小穗、剑叶及叶鞘长度sui1与YFM4差异不明显(图1-E~G)。

图1 小麦穗型突变体表型Fig.1 Phenotype of wheat spike mutant
由图2可以看出,拔节之前,sui1与YFM4幼穗发育差异不明显(图2-A);拔节后,YFM4幼穗穗轴开始伸长,sui1幼穗穗轴伸长缓慢(图2-B);孕穗期之后,YFM4穗轴继续伸长,而sui1穗轴未伸长,小穗在穗轴上紧密排列(图2-C、D);花后20 d调查自然发病田块突变体和野生型赤霉病发病情况,可知sui1较YFM4赤霉病发病程度加重(图2-E)。

图2 YFM4和sui1穗型发育Fig.2 Spike growth of YFM4 and mutant sui1
农艺性状调查结果显示,突变体sui1株高、穗长、穗下节间长度极显著低于野生型,可孕穗数、每穗粒数、千粒重与野生型无显著差异(表1)。由此可见,突变体表型差异主要来自穗轴节和穗下节间长度的缩短,与小穗数、穗粒数及粒重等穗部性状关系不明显。推断突变体穗轴长度的变化与幼穗早期发育、小穗和小花分生组织及花器官原基的分化无明显关系,突变来自拔节抽穗后期的生长发育。
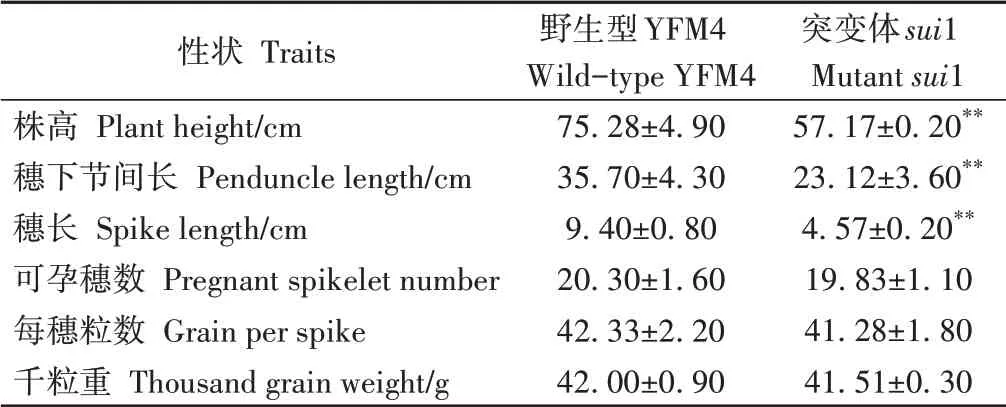
表1 YFM4和突变体sui1主要农艺性状Table 1 Major agronomic traits of YFM4 and mutant sui1
2.2 穗型突变体转录组分析
2.2.1 RNA-Seq结果分析 为了明确YFM4背景的sui1突变体穗型变化的分子基础,分别选取T1野生型穗轴节(A1、A2)、T1突变体穗轴节(B1、B2)、T2野生型穗轴节(C1、C2)、T2突变体穗轴节(D1、D2)和T1野生型穗下节(E1、E2)、T1突变体穗下节(F1、F2)为材料提取总RNA,进行RNA-Seq。经过测序质量控制(去除含有接头的reads;去除N比例大于10%的reads;去除质量值Q≤10的碱基数占整条read的50%以上的reads),共得到1.7×105M clean data,各样品Q30碱基百分比均≥95.26%。将每个样本的高质量reads比对到小麦中国春参考基因组(RefSeq v.1.1)上,对比率在91.32%~94.73%之间(表2)。
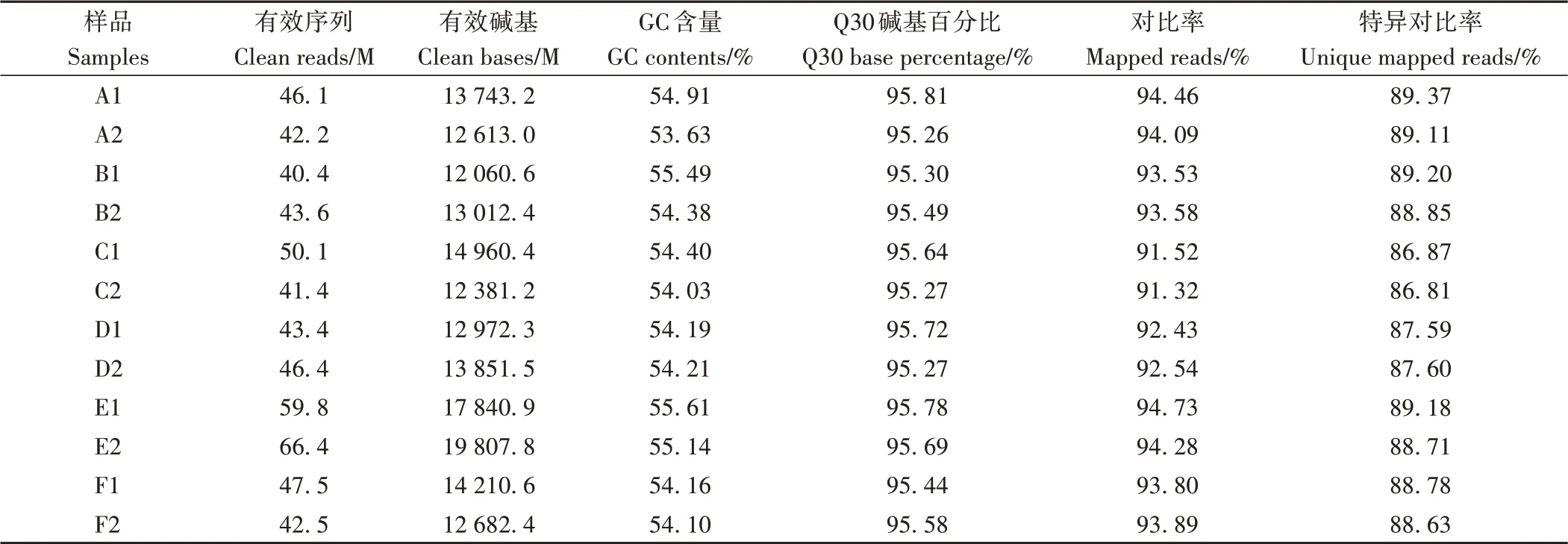
表2 测序数据评估Table 2 Statistics results of sequencing raw data
2.2.2 单核苷酸多态性(single nucleotide polymorphism, SNP)位点 根据SNP位点碱基替换的不同方式,可以将SNP位点分为转换(transition)和颠换(transversion)两种类型。根据SNP位点的等位(allele)数目,可以将SNP位点分为纯合型SNP位点(只有一个等位)和杂合型SNP位点(两个或多个等位)。对各样品筛选出的SNP位点数目、转换类型比例、颠换类型比例以及杂合型SNP位点比例进行统计(表3),SNP位点杂合性比例低于16.04%,SNP位点碱基转换多于颠换。
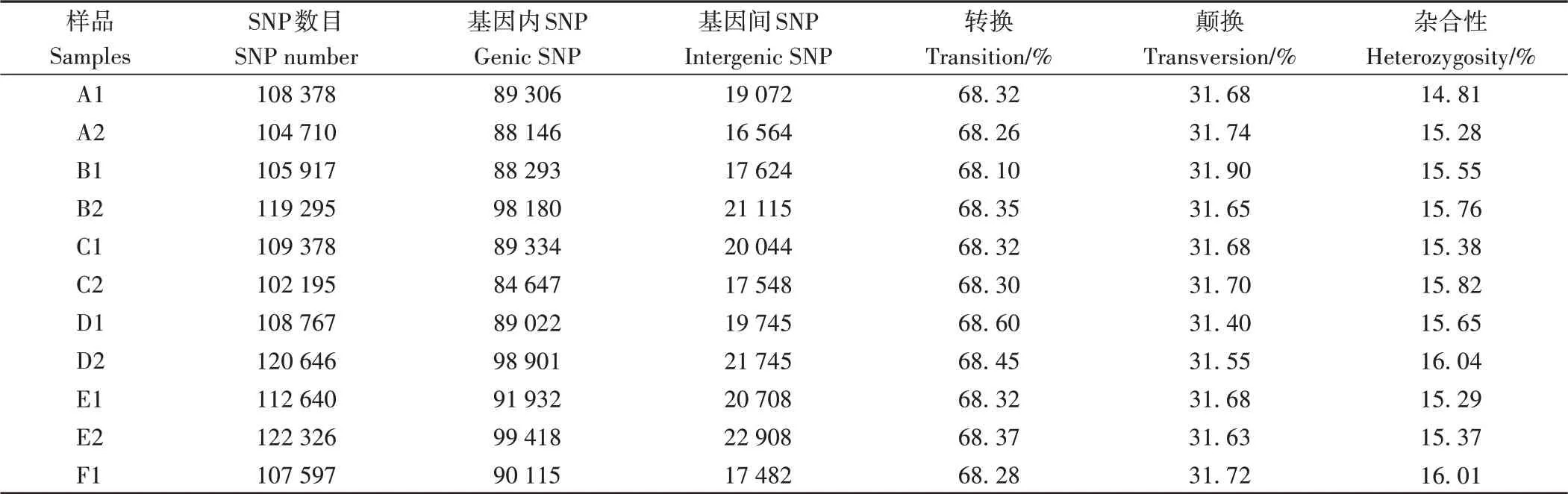
表3 SNP位点统计Table 3 Statistics results of SNP locus
2.2.3 差异表达基因 基因表达具有时间和空间特异性,为了确定YFM4和sui1之间基因表达的差异,根据每千个碱基转录每百万映射读取的值(fragments per kilobase of exon model per million mapped fragments, FPKM)对基因表达水平进行标准化。所有唯一映射读取的fragments被用来计算基因FPKM值。根据DESeq2分析结果,以差异倍数(fold change)≥2,且错误发现率(false discovery rate, FDR)<0.01作为标准进行筛选。由表4和图3可知,T1阶段,穗轴节在突变体和野生型之间共筛选出11 200个差异表达基因(differential expressed genes, DEGs),其 中 上调 基因6 182个(占 比55.2%),下 调 基 因5 018个(占 比44.8%);穗下节间在突变体和野生型之间共筛选出28 641个DEGs,其中上调基因14 632个(占总DEGs的51.1%),下调基因14 009个(占总DEGs的48.9%)。
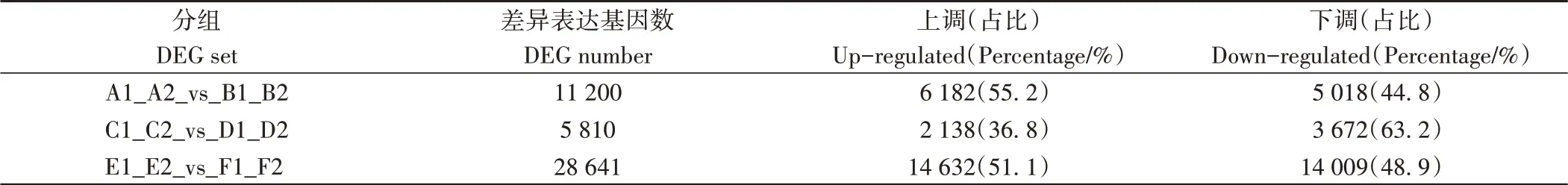
表4 差异表达基因数目Table 4 Number of differentially expressed genes
T2阶段,穗轴节在突变体和野生型之间共筛选出5 810个DEGs,其中上调基因2 138个(占总DEGs的36.8%),下调基因3 672个(占总DEGs的63.2%)。不同器官组织,野生型和突变体穗下节的DEGs多于穗轴节;不同表达时期,T1野生型和突变体穗轴节的DEGs多于T2。将三组差异基因进行维恩图绘制,有差异共表达基因2 526个,其中上调基因890个(图3-A),下调基因1 636个(图3-B)。
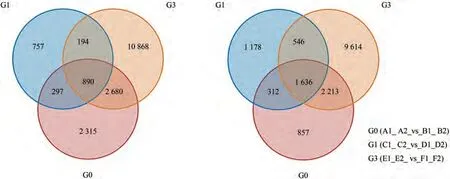
图3 差异表达基因韦恩图Fig.3 Wayn plot of DEGs
2.2.4 差异表达基因GO分类分析 为了解析YFM4和sui1差异表达基因的生物学意义,对上述基因进行GO富集分类。差异表达基因分别注释在生物学过程(biological process)、细胞组分(cellular component)和分子功能(molecular function)的55个分类条目上。对T1突变体和野生型穗轴节11 200个差异表达基因进行GO功能注释,共有7 736个基因获得注释(图4-A);对T2突变体和野生型穗轴节5 810个差异表达基因进行GO功能注释,共有3 344个基因获得注释(图4-B);对T1突变体和野生型穗下节间28 641个差异表达基因进行GO功能注释和分析,共有20 878个基因获得注释(图4-C)。
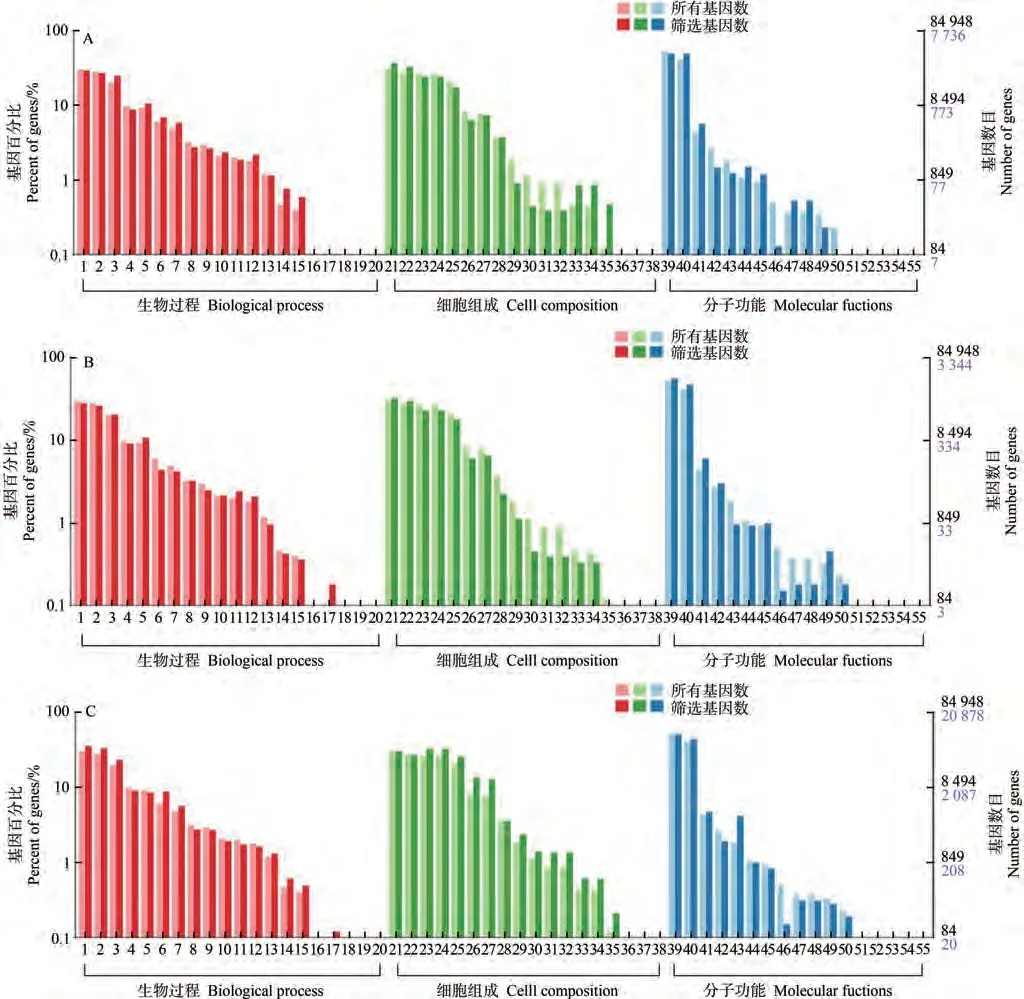
图4 差异表达基因GO注释分类Fig.4 GO classification of differentially expressed genes
分析T1、T2不同时期YFM4和sui1差异表达基因发现(图5-A,B),生物学过程中,T1时期以碳水化合物代谢过程(carbohydrate metabolic process)的基因数最多(7.36%),细胞壁组织(cell wall organization)次之(3.75%);T2时期以防御反应(defence response)的基因数最多(9.73%),碳水化合物代谢过程次之(6.59%)。细胞组分中,T1时期以细胞膜(membrane)功能组拥有的基因数最多(6.72%),质膜(plasma membrane)功能组次之(4.20%);T2时期以叶绿体(chloroplast)功能组拥有的基因数最多(4.80%),叶绿体类囊体膜(chloroplast thylakoid membrane)功能组次之(2.64%)。分子功能注释的基因在T1、T2两个时期富集相同,大部分集中在ATP结合(ATP binding)、蛋白激酶活性(protein kinase activity)上。
分析穗轴节、穗下节不同组织YFM4和sui1差异表达基因发现(图5-B,C),生物学过程中,穗下节以转变(translation)的基因数最多(6.71%)。细胞组分中,穗下节以核小体(nucleosome)功能组拥有基因数最多(3.66%)。分子功能注释基因穗下节大部分基因除集中在ATP结合外,还主要富集在核糖体的结构成分(structural constituent of ribosome)上。
综上分析认为,不同时期sui1较YFM4穗轴节的基因表达差异先来自碳水化合物代谢过程,后因穗轴伸长受到抑制及穗部病害的发生,导致防御反应的差异基因增多;不同组织代谢过程不同,生物学过程和细胞组分差异较大,但分子功能注释大多集中在ATP结合上。
2.2.5 差异表达基因KEGG富集分析 通过对差异表达基因的KEGG富集分析,可以预测基因参与的关键代谢通路,通过对差异表达基因的通路注释分析可以进一步解读基因的功能。对T1突变体和野生型穗轴节注释到的7 736个差异表达基因进行KEGG分析(图5-A),发现这些基因富集到133个代谢通路中,选取富集分析结果中P值最小(富集最显著)的前20条通路途径并绘制散点图。富集因子最高的是光合生物中的碳固定(carbon fixation in photosynthetic organisms)途径,富集因子2.80,富集差异表达基因数量最多的是植物-病原体互作(plant-pathogen interaction)途径,富集到了518个差异表达基因。对T2突变体和野生型穗轴节注释到的3 344个差异表达基因进行KEGG分析(图5-B),发现这些基因富集到124个代谢通路中。富集因子最高的是光合作用-天线蛋白(photosynthesisantenna proteins)途径,富集因子4.85,富集差异表达基因数量最多的是植物-病原体互作途径,富集到了330个差异表达基因。推测上述途径对小麦突变体的穗轴节及穗型变化具有重要作用。
对穗下节注释到的20 879个差异表达基因进行KEGG分析(图5-C),发现这些基因共富集到136个代谢通路中。富集因子最高的是光合作用-天线蛋白途径,富集因子3.54,前20条通路中富集差异表达基因数量最多的是核糖体(ribosome)途径,但所有通路中植物-病原体互作途径富集差异表达基因数量仍为最多,有995个差异表达基因。推测上述途径对小麦突变体的穗下节及株高的变化具有重要作用。穗下节的差异表达基因富集不同于穗轴节,可能与穗轴节是穗型变异直接功能器官,而穗下节间通过茎秆碳水化合物的运输间接影响穗轴的伸长有关。在植物-病原体互作通路中进行差异共表达基因筛选,筛选出相关基因160个,主要含防御反应相关基因63个(39%),蛋白激酶活性相关基因21个(13%),ADP结合相关基因18个(11%),推测上述基因可能与sui1突变性状相关。
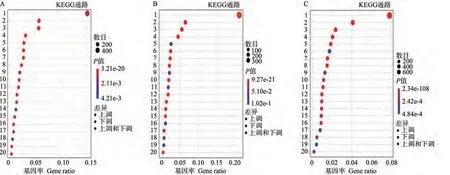
图5 KEGG富集气泡分析图Fig.5 Bubble diagram of the KEGG enrichment
3 讨论
基于RNA-Seq的转录组分析可以在整体转录水平揭示全基因组内基因的表达情况,通过转录组分析可以获得所有基因的表达趋势,从而用于基因功能和调控的深度研究[12-14]。温宏伟等[15]和李玲红等[16]基于转录组测序研究,发现了与蜡质功能相关的差异显著表达基因;范小锋等[17]对小麦直立突变体进行转录组分析,发现差异表达基因主要富集在植物激素信号转导、淀粉和蔗糖的生物合成等通路。本研究通过转录组分析,在突变体和野生型之间筛选出差异表达基因2 526个,KEGG分析发现差异表达基因主要富集在植物-病原体互作-光合生物中的碳固定、光合作用-天线蛋白等通路。基于突变体与野生型相关表型鉴定和转录组分析,推测该通路中的关键基因和小麦穗型突变后赤霉病发病程度加重有关。
相同的穗长上容纳更多的小穗数,可使穗密度增加[18];但小穗数相同,穗轴节缩短也会导致穗密度增加。陈树林[19]通过EMS诱变苏麦3号获得矮秆密穗突变体NAU164(Rht23),认为穗型变密是由穗轴节间变短所致,株高变矮是各节间长度变短所致。本试验基于YFM4野生型的突变,研究发现穗轴节长度变短、小穗数不变,穗粒数相差不大,密穗产生的原因也主要在于穗轴节的长度缩短。不同于突变体NAU164,突变体sui1的突变主要来自拔节孕穗期之后,基部节间长度不变,穗下节间长度缩短,株高降低。前人研究还发现,小麦赤霉病抗性与株高及小穗密度具有相关性[20-22],徐晴等[23]研究矮秆基因在我国不同麦区分布及其对赤霉病抗性的影响,发现不同矮秆基因对小麦穗性状的遗传差异可能是导致赤霉病抗性和不同麦区矮秆基因选择利用差异的部分原因。本研究中突变体sui1穗型较野生型穗型变密、株高变矮后,赤霉病抗性减弱,与前人研究结果一致。
小麦的穗密度受多个遗传位点控制[24-26]。其中,基因C的显性作用使得密穗小麦穗部比普通小麦穗部更紧凑。基因C被定位在小麦2D染色体上,通过研究四倍体小麦和六倍体小麦的密穗突变体,发现并定位了4个决定穗密度的位点(C739、C17648、Cpm和Cp)[27]。Q基因是重要的驯化基因,具有多向性,可以同时影响小麦颖壳强度、穗轴韧性、穗长、株高、抽穗期等多个性状[28]。5A染色体上的Q基因是控制驯化相关性状的主效基因。本研究在植物-病原体互作通路中筛选出160个差异共表达基因,其中有8个基因位于2D染色体上,3个基因位于5A染色体上,筛选到的位于2D和5A染色体上的基因位点不同于C基因和Q基因控制穗密度的相关位点,可能为新的突变基因位点。为进一步挖掘和验证突变基因与sui1穗部性状相关性,后续工作还需要建立遗传群体,进行相关基因位点的验证和功能分析。
4 结论
本研究在扬辐麦4号突变体库中获得一个穗轴节和穗下节明显缩短、穗密度增加、株高降低、赤霉病抗性变差的穗型突变体sui1。通过对小麦植株不同时期、不同组织的转录组分析,比较突变体和野生型间的基因表达水平,筛选出差异共表达基因2 526个,其中上调表达基因890个,下调表达基因1 636个。通过功能注释和富集分析发现,差异基因主要涉及植物-病原体互作、光合生物中的碳固定和光合作用-天线蛋白等通路,推测通路中关键基因的表达可能与突变体sui1相较于野生型穗部性状产生的差异有关。
猜你喜欢
四川蚕业(2022年2期)2022-11-19
中国农业大学学报(2022年4期)2022-05-26
中国种业(2022年4期)2022-04-19
农业工程学报(2021年14期)2021-10-12
作物学报(2021年7期)2021-04-27
世界科学技术-中医药现代化(2020年2期)2020-07-25
临床医药文献杂志(电子版)(2017年11期)2017-05-17
上海农业学报(2017年3期)2017-04-10
安徽医科大学学报(2016年12期)2017-01-15
天津医科大学学报(2015年2期)2015-12-22
