
辣椒泛素结合酶变体类似蛋白基因克隆及其与氮吸收利用的相关性分析
2023-01-18 14:39王春萍张世才李怡斐杨小苗段敏杰黄启中黄任中吴红
核农学报 2023年3期
王春萍 张世才 李怡斐 杨小苗 段敏杰 黄启中 黄任中 吴红
(1重庆市农业科学院生物技术研究所逆境农业研究重庆市市级重点实验室, 重庆 401329;2重庆市农业科学院蔬菜花卉研究所, 重庆 401329)
泛素修饰并降解蛋白质是真核生物调节细胞活动与功能的重要途径,在植物细胞生理和对环境条件的反应中具有广泛作用[1]。泛素化-蛋白酶体系统主要由泛素、泛素激活酶(Ub-activating enzyme, UBA 或E1)、泛 素 结 合 酶(Ub-conjugating enzyme, UBC 或E2)、泛素连接酶(Ub ligase, E3)、去泛素化酶和靶蛋白组成[2]。E2在E1与E3之间起协调作用,能将E1激活的泛素分子转移到底物或E3[3-4]。所有E2都含有一个由150~200个氨基酸组成的含有一个半胱氨酸活性位点的UBC结构域。UEVs(ubiquitin E2 variants)在氨基酸序列和结构上与E2相似,但UBC结构域缺乏半胱氨酸活性位点,不能与泛素直接结合,因此其功能与E2不同[5]。已有研究证明UEVS可通过与E2形成复合物催化底物泛素化[6-7]。
关于酵母、动物和人类UEVs的功能已有较多报道,但该蛋白家族在植物中的功能最早只在拟南芥(Arabidopsis thaliana)中有较深入报道,近年来在水稻(Oryza sativa)、番茄(Solanum lycopersicum)、短柄草(Brachypodium distachyon)等植物中也开展了一些研究。如在拟南芥中,COP10在植物的光形态建成中起着重要作用[5-7],AtUEV1D能够增强DNA损伤的耐受性[8];番茄Suv能与Fin3/SI-Ubc13-2协同调控烟草对丁香假单胞菌的免疫[9];在短柄草中,BdUev1B和BdUev1C参与DNA损伤反应,BdUev1A可能参与亚细胞结构定位等生理活动[10];在水稻中鉴定出4个参与DNA损伤修复的OsUEV1[11],其中OsUEV1B参与磷的代谢途径[12]。可见,目前关于UEVs功能的研究较多集中于DNA损伤修复方面,其在非生物逆境方面的作用鲜有报道。
基于重庆市农业科学院辣椒研究团队前期获得的低氮胁迫下辣椒差异转录组数据[13],本研究克隆了辣椒(Capsicum annuum)UEVs类似蛋白基因,命名为CaUev1DL,构建该基因过表达载体,转化至烟草(Nicotiana tabacum)后进一步对T2代转基因烟草进行低氮胁迫后的表型和生理分析,推测该基因在耐低氮中的作用,以期为其功能研究和在基因工程中的运用提供参考。
1 材料与方法
1.1 试验材料
基因克隆所用材料为重庆市农业科学院辣椒研究团队保存的簇生朝天椒自交系750-1。遗传转化材料为本氏烟草。
1.2 CaUev1D-L基因的克隆和生物信息分析
根据前期转录组数据注释到的辣椒ubiquitinconjugating enzyme E2 variant 1D-like基因,从NCBI数据库(https:∕∕www.ncbi.nlm.nih.gov∕)中搜索其全基因序列(LOC107877293),设计该基因全长PCR扩增引物CaUev1D-L-F (5´-ATGACTCTCGATTCAGGA-3´)和CaUev1D-L-R (5´-TCACCCATTTTCTTCAAC-3´),以辣椒自交系750-1的叶片cDNA为模板进行PCR扩增。总反应体系20 µL:DNA模板1 µL、2×TaqMaster Mix (上海Novoprotein) 10 µL、上下游引物各1 µL、ddH2O 7 µL。PCR反应条件:94 ℃预变性5 min;94 ℃变性30 s,57 ℃退火30 s,72 ℃延伸60 s,30个循环;72 ℃终延伸10 min。将扩增出的相应基因片段利用同源重组的方式无缝连接到改造后的表达载体pBI121上(在原pBI121上的T-DNA插入区域增加35S∶GUS∶NPTII融合基因,用于卡纳霉素筛选和GUS染色鉴定)。连接产物通过热激发转入大肠杆菌DH5α中,并通过PCR菌落鉴定,筛选阳性克隆子进行测序。
利用Protparam (http:∕∕web.expasy.org∕protparam) 在线分析基因编码蛋白氨基酸的理化性质,利用SOPMA (https:∕∕www.expasy.org∕proteomics_sopma) 和Phyre2 (http:∕∕www.sbg.bio.ic.ac.uk∕phyre2)进行蛋白质二级、三级结构预测。
1.3 CaUev1D-L基因烟草转化
提取阳性克隆子菌落的表达载体pBI121质粒,转化农杆菌EHA105感受态。将筛选出的农杆菌EHA105菌株浸染烟草无菌苗叶片(约1 cm2)伤口,经共培养、抗性筛选、生根和移栽等过程获得T0代转基因植株,经2次繁殖后获得T2代种子。
1.4 转基因烟草T2代植株低氮处理
挑选饱满一致且无病虫害的烟草种子,均匀放入盛有滤纸的培养皿中,加入蒸馏水,保持培养皿中滤纸湿润,置于人工气候箱,25~28 ℃黑暗条件下发芽。将发芽的种子用蒸馏水清洗后分单株移到装有基质的穴盘中培养,基质配比椰糠∶珍珠岩∶蛭石体积比为5∶3∶2,用营养液定量浇灌,待植株长至两叶一心时进行低氮处理,营养液配方参照辣椒低氮处理方法[14]。各处理营养液pH值为6.2~6.4,每5 d更换一次营养液,培养温度为25~28 ℃,每天光照12 h,光照强度为200 µmol·m-2·s-1,空气相对湿度为70%~80%。每个处理设3次重复,每个重复10株植株。
1.5 低氮胁迫下转基因烟草植株CaUev1D-L基因的表达分析
低氮处理28 d后分别取对照和处理材料的功能叶,液氮处理后于-80 ℃保存,或直接提取RNA。用反转录试剂盒(美国Bio-Rad)对RNA进行去基因组DNA和反转录得到cDNA。用Bio-Rad CFX96 Real-Time System荧光定量PCR仪进行实时荧光定量PCR,反应体系20 µL:iQ SYBR Green Supermix (美国Bio-Rad) 10 µL、上下游引物各0.5 µL、cDNA模板1.0 µL,ddH2O补足体积。每个样品重复3次。内参基因为烟草Tubulin基因(LOC107800889)。
1.6 低氮胁迫下转基因烟草植株形态和植株干物质积累分析
处理28 d后,分别测定每棵植株的株高、茎粗、叶片数、叶绿素相对含量(soil and plant analyzer development,SPAD)值、根系形态、茎叶干重、根干重,并根据以下公式计算单株干重、根冠比:
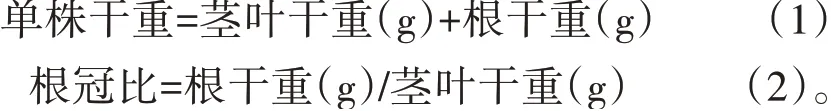
2 结果与分析
2.1 CaUev1D-L基因的克隆及序列分析
根据前期辣椒转录组数据注释到的低氮胁迫下差异表达基因(ubiquitin-conjugating enzyme E2 variant 1D-like)为参考序列设计引物,以辣椒材料750-1的叶片cDNA为模板对该基因进行PCR扩增,获得全长为402 bp的编码序列,命名为CaUev1D-L(图1)。该基因编码133个氨基酸,相对分子质量为15.09 kDa,等电点为5.35。利用NCBI的CD search(Conserved Domain Search Service)对CaUev1D-L蛋白的结构功能域进行预测,发现其18~102氨基酸区间为UBC结构域。

图1 CaUev1D-L基因的核苷酸序列和对应的氨基酸序列Fig.1 Nucleotide sequence and corresponding amino acid sequence of CaUev1D-L
2.2 CaUev1D-L蛋白的结构特征分析
利用SOPMH在线预测CaUev1D-L蛋白的二级结构,其中无规则卷曲所占比例最多(43.61%),其次是延展链(29.32%),α螺旋(19.55%),β转角(7.52%)。利用Phyre2进一步预测H蛋白的三级结构,参照模板为c2hlwA,一致性和覆盖率分别为100%和96%,从结构组成上看,以无规则卷曲为主,与二级结构预测结果一致(图2)。
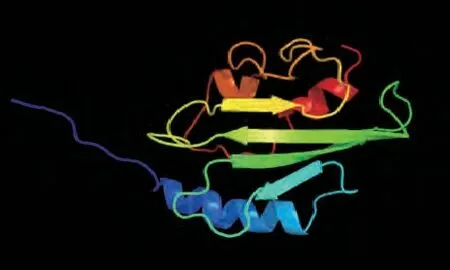
图2 CaUev1D-L蛋白的三级结构预测模型Fig.2 Tertiary structure prediction model of CaUev1D-L
2.3 CaUev1D-L蛋白的多序列比对和系统进化分析
将CaUev1D-L的氨基酸序列与辣椒(XP_016546430.1)、烟草(XP_016434792.1)、番茄(NP_001306880.1)和马铃薯(XP_006339155.1)等4种茄科作物的Uev1D(ubiquitin-conjugating enzyme E2 variant 1D)氨基酸进行比对。结果表明,在氨基酸序列末端CaUev1D-L比4个Uev1D少14个氨基酸,在UBC结构域与4个Uev1D有6个氨基酸的差异,而4个Uev1D的序列十分保守(图3)。进一步利用MEGH7.0软件中的Neighbor-joining方法分析CaUev1D-L蛋白在不同植物物种之间的进化关系。结果显示,CaUev1D-L蛋白与辣椒的几个同源蛋白同处于同一进化分支上,与其他物种相距较远,与辣椒Uev1D不在同一进化分支上(图4)。

图3 CaUev1D-L与部分茄科作物Uev1D氨基酸序列比对结果Fig.3 Amino acid sequence comparison between CaUev1D-L and Uev1D of some Solanaceae crops

图4 CaUev1D-L氨基酸序列的系统进化分析Fig.4 Phylogenetic analysis of amino acid sequence of CaUev1D-L
2.4 CaUev1D-L基因功能分析
2.4.1 不同氮素水平下CaUev1D-L基因的表达特点 分别对正常氮水平和20%氮水平下CaUev1D-L在转基因烟草和相应野生型烟草叶片的表达情况进行分析。结果表明,在两种氮水平下CaUev1D-L在野生型植株的表达量都非常低,而在转基因植株的表达量显著提高,说明目标基因在转基因植株中能够稳定高表达(图5)。
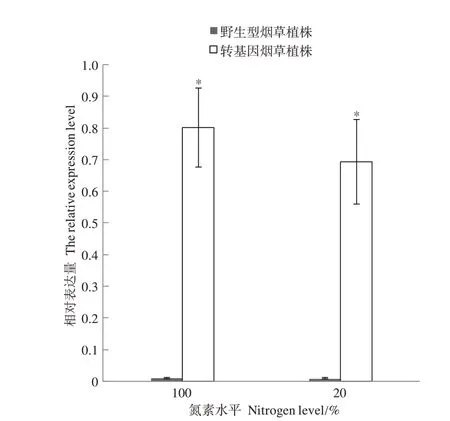
图5 不同氮素水平下CaUev1D-L表达情况Fig.5 Expression of CaUev1D-L under different nitrogen levels
2.4.2 不同氮素水平下转CaUev1D-L基因烟草植株形态特点 在正常施氮素水平下,与野生型植株相比,转CaUev1D-L基因植株的株高、茎粗、叶片数、SPAD值和根长等5个形态指标值均与野生型植株差异不显著(图6)。在低氮胁迫下,转基因烟草植株的根长显著大于野生型植株,其余4个形态指标在两植株之间差异不显著(图7)。
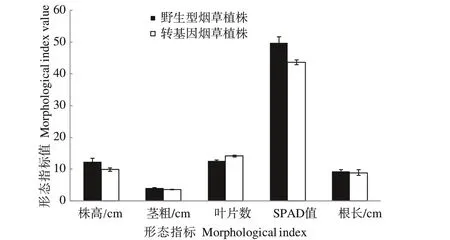
图6 正常氮水平下烟草植株的形态特点Fig.6 Morphological characteristics of tobacco plants under normal nitrogen level
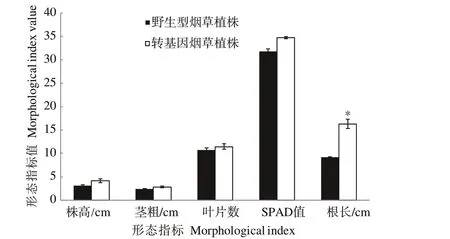
图7 低氮水平下烟草植株的形态特点Fig.7 Morphological characteristics of tobacco plants under low nitrogen level
2.4.3 不同氮素水平下转CaUev1D-L基因烟草干物质积累特点 在正常氮素水平下,转CaUev1D-L基因烟草植株茎叶干重显著小于野生型烟草植株(图8)。但在低氮水平下,转基因植株的茎叶干重和根干重显著大于野生型植株(图9),说明在低氮水平下CaUev1D-L的过量表达能够促进氮的吸收利用。
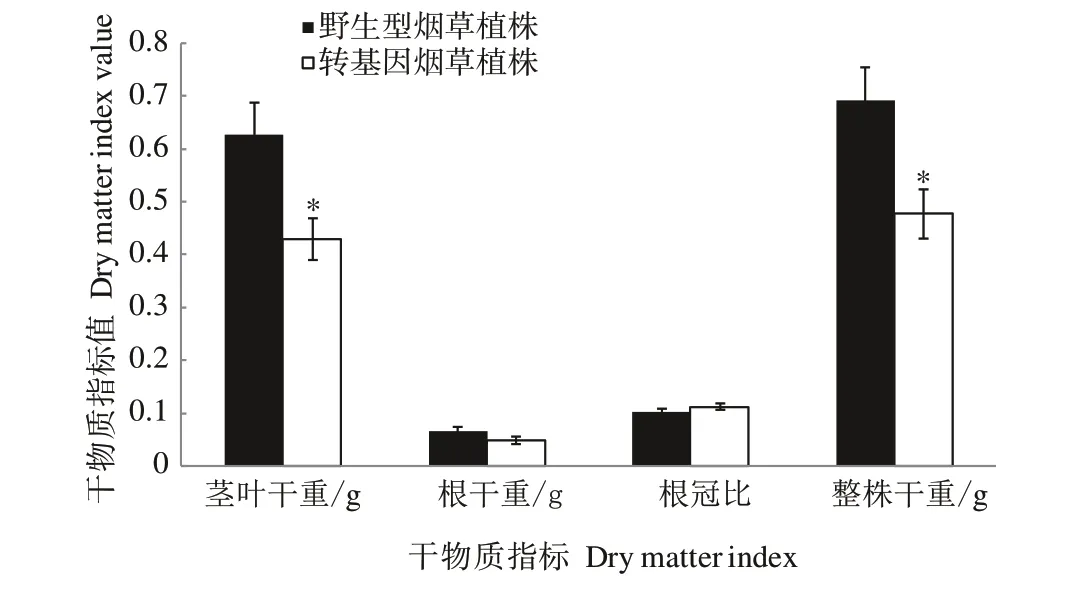
图8 正常氮水平下转基因烟草植株干物质积累特点Fig.8 Dry matter accumulation characteristics of transgenic tobacco plants under normal nitrogen level
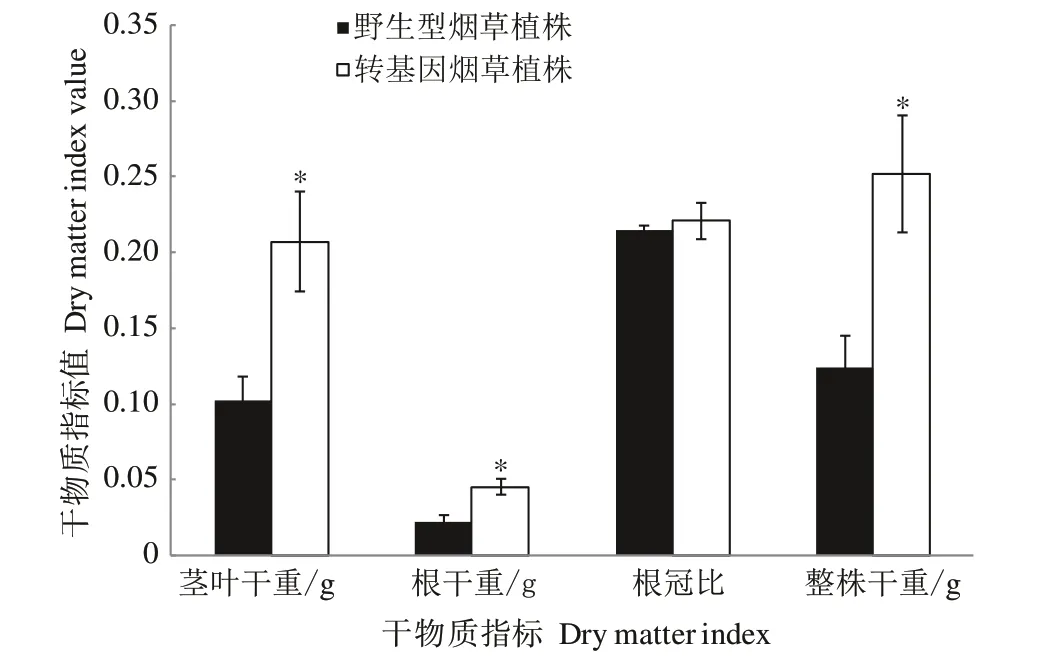
图9 低氮水平下转基因烟草植株干物质积累特点Fig.9 Dry matter accumulation characteristics of transgenic tobacco plants under low nitrogen level
3 讨论
不同的泛素链具有不同的拓扑结构和功能,如Lys48-泛素链导致蛋白质降解,Lys63-泛素链通常参与信号转导[15-16]。Ubc13基因参与了多种生命活动,如Ubc13在拟南芥的DNA损伤反应[8]、顶端优势[17]、铁代谢[18]、生长素信号转导[19]、低温胁迫反应[20]以及免疫反应[21]等过程都发挥着重要作用。鉴于在植物中发现了多个UEV基因,且这些基因的表达水平和功能都存在差异,有学者推测,在Ubc13-Uev复合体中真正起调节作用的是UEVs蛋白。UEVs蛋白可能参与多种调节过程[10-11,22],UEVs在氮素代谢途径中的研究还未见报道。重庆市农业科学院辣椒研究团队前期研究发现,Uev1D类似蛋白基因(LOC107877293)能够响应辣椒苗期低氮胁迫反应[13],因此本研究克隆了该基因,命名为CaUev1D-L,并对其功能进行了研究。
本研究结果表明,在正常氮水平下,过表达辣椒CaUev1D-L导致烟草的干物质积累显著下降,而在低氮水平下,转基因烟草的干物质量积累显著增加,初步判断CaUev1D-L参与了氮代谢调控途径,且在低氮胁迫下促进了氮素的吸收利用。氮代谢途径可分为三个过程:吸收、同化和再分配[23-24]。氮的再分配过程要经历蛋白质的降解,泛素化途径是蛋白质降解的主要途径,因此,蛋白质的泛素化与氮代谢途径具有紧密联系。Peng等[25]在拟南芥中发现了一个低氮适应性调节蛋白NLA,该蛋白属于RING型E3连接酶[26]。与野生型植株相比,nla基因突变体植株表现出对低氮十分敏感且早衰,这是由于nla基因突变改变了NLA蛋白的亚细胞定位,导致其不能与泛素结合酶8(AtUBC8)结合,从而影响了其正常行使功能。后续研究表明,nla基因突变体早衰是由于植物体中磷(Pi)过量积累所致,且nla基因突变体表型特征与PHO2突变体表型相同,而PHO2是一种E2结合酶[27-30],说明氮代谢与磷代谢具有密切相关性,同时反映了E2与氮代谢的相关性。本研究通过超表达方法证明CaUev1D-L参与了氮代谢途径,再次表明了E2家族成员与氮代谢的相关性。但CaUev1D-L参与氮代谢的机理尚不清楚,后期还需对其作用机理和调控网络进行深入研究。
另外,本研究对比了辣椒、烟草、番茄和马铃薯等品种的Uev1D,其UBC结构域的氨基酸序列完全一致,只在蛋白的氨基酸末端有较低的多态性。然而CaUev1D-L在上游、UBC结构域以及末端与Uev1D都存在差异,且比Uev1D末端少了14个氨基酸差异,由此推测CaUev1D-L与Uev1D的功能可能存在较大差异。
4 结论
本研究从辣椒中克隆了CaUev1D-L基因,该基因编码的蛋白与茄科作物的Uev1D蛋白氨基酸序列末端和UBC结构域都存在差异,与辣椒的Uev1D蛋白不在同一进化分支,推测CaUev1D-L与Uev1D的功能可能存在差异。转CaUev1D-L基因烟草植株在正常氮水平下茎叶干重显著小于野生型植株,在低氮水平下茎叶干重和根干重显著大于野生型植株,初步判断该基因参与了氮代谢调节途径,且在低氮水平下能够促进植物对氮的吸收利用。
猜你喜欢
学与玩(2022年10期)2022-11-23
今日农业(2022年3期)2022-06-05
小哥白尼(神奇星球)(2021年6期)2021-07-28
快乐作文(1.2年级)(2020年8期)2020-09-10
中外文摘(2020年9期)2020-06-01
医学综述(2020年11期)2020-02-16
创新科技(2015年1期)2015-12-24
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15
中国医学科学院学报(2015年5期)2015-03-01
现代检验医学杂志(2015年2期)2015-02-06
