
基于TLR4/TAK1/NF-κB途径探讨MCAO大鼠缺血局部脑皮质微血管新生的内在机制
2023-01-18 09:02张春玲梁超王宝爱徐玉婷
中国老年学杂志 2023年2期
张春玲 梁超 王宝爱 徐玉婷
(海口市中医医院,海南 海口 570216)
脑血管疾病在中老年人中较常见,且因其高发病率、高致残率和高死亡率,给人类的健康带来了严重的威胁〔1〕。局灶性脑缺血发生后,其损害程度取决于血管细胞、神经元和胶质细胞之间活跃而复杂的相互作用〔2〕。脑缺血引起的脑微血管内皮细胞(BMECs)损伤是导致血脑屏障破坏的主要原因〔3〕。Toll样受体(TLRs)广泛表达于天然免疫细胞,在早期天然免疫中起着重要作用,其中TLR4被激活后可活化核因子(NF)-κB,进而诱导和调控炎症反应〔4〕。在大脑中动脉栓塞(MCAO)小鼠模型中TLR4表达水平升高,此外,缺乏TLR4的小鼠神经功能缺陷明显减少〔5〕。转化生长因子(TGF)-β激活激酶(TAK)1是一种MAP3激酶,位于与脑缺血损伤有关的几种途径的上游〔6〕。TAK1可被TGF-β、白细胞介素(IL)-1β、肿瘤坏死因子(TNF)-α等缺血性损伤后分泌的细胞因子激活,而 TAK1的激活反过来又会诱导促炎细胞因子和趋化因子的产生,并激活已知在脑缺血病理中起有害作用的免疫细胞,从而参与脑缺血的发病进程〔7〕。也有研究表明,TLR4与肿瘤坏死因子受体相关因子(TRAF)6结合并促进其活化,活化的TRAF6可与TAK1及其TGF-β活化激酶结合蛋白(TAB)1相互作用促使TAK1活化,TAK1被激活后可解除NF-κB的抑制,而NF-κB被活化后可导致炎症因子的广泛表达,从而减轻或加重炎症反应造成的脑缺血损伤〔8,9〕。但TLR4/TAK1/NF-κB途径是否与MCAO大鼠缺血局部脑皮质微血管新生相关尚不清楚。本研究旨在基于TLR4/TAK1/NF-κB途径探讨MCAO大鼠缺血局部脑皮质微血管新生的内在机制。
1 材料与方法
1.1实验动物 40只成年SPF级健康雄性SD大鼠,体重(230±10)g,由珠海百试通生物科技有限公司提供,在温度20~25℃,湿度40%~60%的条件下分笼饲养。
1.2试剂 siRNA-TLR4 纯化腺病毒和阴性对照病毒均购自上海吉凯基因生物技术有限公司;Ki67、血管内皮生长因子(VEGF)、促血管生成素(Ang)-1、Ang-2抗体购自美国Abcam公司;TLR4、TGF、TAK1、NF-κB p65和β-actin鼠单克隆抗体购自美国eBioscience公司。
1.3仪器 荧光显微镜(日本NIKON公司,型号 Eclipse 80i);病理切片机(美国赛默飞公司,型号Thermo HM);包埋机(武汉俊杰电子有限公司,型号JB-P5);PCR仪(美国Bio-Rad公司,型号CFX96 Touch)。
1.4动物分组及造模 40只雄性SD大鼠,适应性喂养1 w,按随机数字表法分为4组,其中10只为假手术(Sham)组,另30只采用线栓法建立MCAO大鼠模型,并将其分为模型(Model)组、阴性病毒(NC)组和siRNA-TLR4转染(siRNA-TLR4)组,具体造模方法如下:所有大鼠于术前禁食12 h,仰卧位固定,采用3.5%异氟烷进行麻醉,分离并暴露一侧颈总动脉、颈外动脉及颈内动脉,结扎颈外动脉,并夹闭右侧颈总动脉,在颈总动脉远端处剪3 mm小口,将0.28 mm的尼龙线从小口处穿入至大脑中动脉,固定线栓,结扎并剪去线栓尾端,缝合切口并消毒。Sham组仅分离血管不插入线栓进行造模。
1.5给药剂量 造模后1 h,Sham组与Model组均给予尾静脉注射等量生理盐水,NC组给予尾静脉注射阴性对照病毒5×109PFU,siRNA-TLR4组给予尾静脉注射siRNA-TLR4腺病毒5×109PFU,所有大鼠术后正常喂养48 h后进行后续实验。
1.6苏木素-伊红(HE)染色法观察缺血局部脑皮质病理变化 各组大鼠于术后48 h,麻醉断头处死后,迅速取出脑组织,用冷生理盐水清洗脑组织并除去脑组织表面残余的水分,取部分脑组织用4%多聚甲醛溶液固定24 h,脱水并用石蜡包埋,按照步骤处理切片,用HE染色法观察大鼠缺血局部脑皮质病理变化。
1.7氯化三苯基四氮唑染色(TTC)检测大鼠脑梗死面积 各组大鼠处死后取脑组织,并制作冠状脑片,严格按照TTC试剂盒说明书进行染色,分析并计算各组大鼠脑梗死面积。
1.8TUNEL染色检测大鼠缺血局部脑皮质细胞凋亡情况 对各组大鼠的石蜡切片进行脱蜡和水化,用0.01 mol/L柠檬酸缓冲液(pH6.0)处理切片,参照TUNEL试剂盒说明书进行染色,显微镜下观察并计算脑皮质细胞凋亡率。
1.9免疫荧光染色检测微血管内皮细胞数目 石蜡切片脱蜡脱水,用蒸馏水浸泡2 min,微波炉内对抗原进行热修复。10%正常山羊血清封闭,室温孵育30 min,以减少非特异性染色。滴加1∶200浓度的Ki67一抗4℃过夜,滴加异硫氰酸荧光素(FITC)标记山羊抗兔IgG荧光抗体37℃染色30 min。4,6-联脒-2-苯基吲哚(DAPI)复染细胞核,缓冲甘油封固后镜检。其中每一步均需PBS漂洗,5 min/次,共3次。
1.10免疫组化法检测VEGF、Ang-1和Ang-2表达水平 石蜡切片脱蜡后,置于3% H2O2中室温孵育10 min,微波炉内对抗原进行热修复。切片冷却至室温后,用磷酸盐缓冲液(PBS)冲洗5 min×3次。10%正常山羊血清封闭,室温孵育10 min,滴加1∶100浓度的VEGF、Ang-1和Ang-2一抗4℃过夜,PBS冲洗5 min×3次。滴加适量生物素标记二抗,室温孵育1 h,PBS冲洗5 min×3次。滴加适量辣根酶标记的链霉卵白素工作液,37℃孵育10 min,PBS冲洗5 min×3次。滴加二氨基联苯胺(DAB)显色剂显色15 min,自来水充分冲洗、复染、脱水、透明、封片,荧光显微镜下观察并记录。
1.11反转录-聚合酶链式反应(RT-PCR)检测TLR4、TAK1、NF-κB p65 mRNA表达 取部分脑组织放入去酶EP管中,用超纯 RNA 提取试剂盒按产品说明书进行实验操作,提取组织样本中总 RNA,按照逆转录试剂盒说明书进行逆转录反应。反应条件:95℃,45 s,95℃,15 s,60℃,60 s,共进行45个循环。每组进行3次独立测定,采用 2-ΔΔCt法对数据进行分析。RT-PCR引物序列:β-actin正向:5′-CTTGTGCAGTGCCAGCC-3′,反向:5′-GCCCAATACGGC CAAATCC-3′;TLR4正向:5′-AATCTGGTGGCTGTGGAGAC-3′,反向:5′-GCTTGGGCTTGAATGGAGT-3′;TAK1正向:5′-CAAAGCTAAGTGGAGAGCAAAAGA-3′,反向:5′-GATAACTGCCGAAGCTCTACAATAA-3′;NF-κB p65正向:5′-TGTGATGAAAGACGGCAC-AC-3′,反向:5′-CTTCTTCTTTCGGGTATTGTTTGG-3′。
1.12Western印迹检测TLR4、TAK1和NF-κB p65蛋白表达 取部分脑组织在研钵中充分研磨,加入裂解液置冰上充分裂解,4℃下离心30 min,吸取上清;采用二喹啉甲酸(BCA)试剂盒测量蛋白质含量,确定每孔上样蛋白量相等。取少量蛋白质配制成上样缓冲液,电泳、转膜、封闭,加入一抗TLR4、TAK1、NF-κB p65和β-actin,4℃过夜;TBST洗涤,加辣根过氧化物酶标记的二抗,室温轻摇1 h,TBST充分洗涤,用 ECL 化学发光试剂显色后,化学发光仪成像。
1.13统计学方法 采用SPSS21.0软件进行方差分析、t检验。
2 结 果
2.1各组缺血局部脑皮质病理变化 Sham组细胞质分布均匀,核膜完整,核仁清晰,神经元排列整齐,且形态完整;Model组和NC毒组可观察到大量不规则的细胞和空泡,细胞核固缩和迁移,胞质和细胞核模糊不清;siRNA-TLR4组缺血半影区细胞形态比模型组和阴性病毒组规则,且脑组织损伤程度降低。见图1。

图1 各组缺血局部脑皮质病理变化(HE染色,×400)
2.2各组脑梗死面积比较 与Sham组相比,Model组、NC组和siRNA-TLR4组脑梗死面积明显增大(P<0.01);与Model组相比,siRNA-TLR4组脑梗死面积明显减小(P<0.01)。见表1。
2.3各组脑皮质细胞凋亡情况 与Sham组相比,Model组、NC组和siRNA-TLR4组脑皮质细胞凋亡率明显升高(P<0.05);与Model组相比,siRNA-TLR4组脑皮质细胞凋亡率明显降低(P<0.05)。见表1、图2。
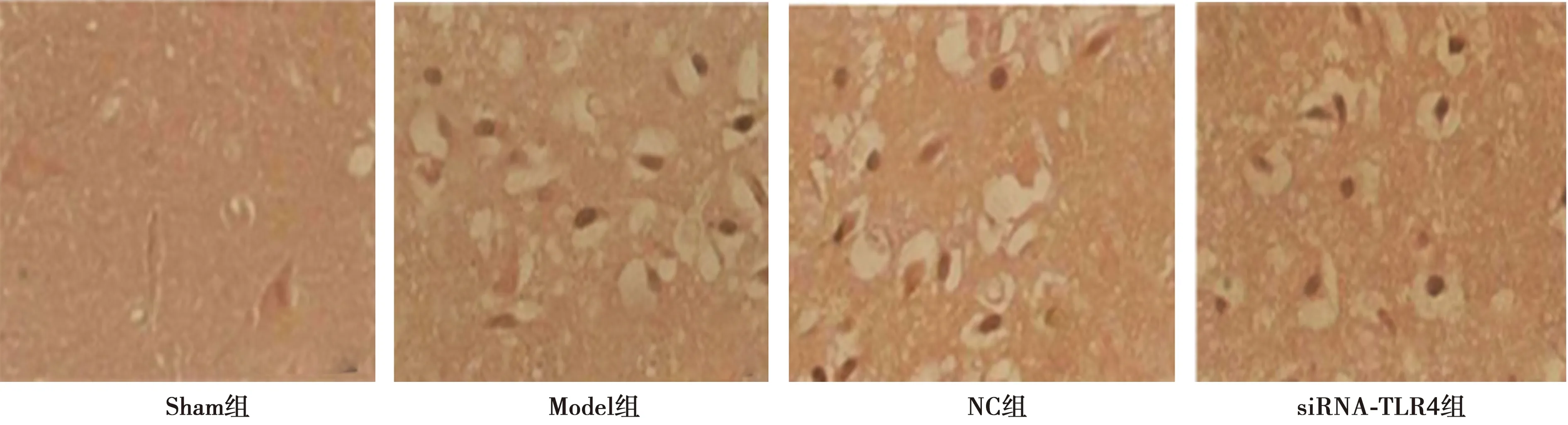
图2 各组脑皮质细胞凋亡情况(TUNEL染色,×400)
2.4各组BMECs增殖数目比较 与Sham组相比,Model组、NC组和siRNA-TLR4组BMECs明显增加(P<0.05);与Model组比较,siRNA-TLR4组BMECs明显增加(P<0.05)。见表1、图3。
表1 各组脑梗死面积和脑皮质细胞凋亡情况、BMECs增殖和脑组织VEGF、Ang-1和Ang-2表达
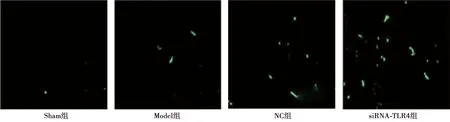
图3 各组脑皮质微血管内皮细胞增殖数目(免疫荧光染色,×200)
2.5各组脑组织VEGF、Ang-1和Ang-2表达比较 与Sham组相比,Model组、NC组和siRNA-TLR4组VEGF、Ang-1和Ang-2表达水平明显升高(P<0.05);与Model组相比,siRNA-TLR4组VEGF、Ang-1和Ang-2表达水平明显升高(P<0.05)。见表1。
2.6各组脑组织TLR4、TAK1、NF-κB p65 mRNA表达 与Sham组相比,Model组、NC组和siRNA-TLR4组TLR4、TAK1、NF-κB p65 mRNA及蛋白表达水平明显升高(P<0.05);与Model组相比,siRNA-TLR4组TLR4、TAK1、NF-κB p65 mRNA及蛋白表达水平明显降低(P<0.05)。见表2、图4、表3。
表2 各组脑组织TLR4、TAK1、NF-κB p65 mRNA表达比较

图4 各组脑组织TLR4、TAK1、NF-κB p65 蛋白表达
表3 各组脑组织TLR4、TAK1、NF-κB p65蛋白表达比较
3 讨 论
脑缺血又称为缺血性脑卒中,占脑卒中的80%以上,且脑缺血是全球死亡率和发病率的主要原因〔10〕。脑血管阻塞导致脑缺血、缺氧,并对大脑造成一系列的病理损害,且脑缺血生理过程通常被认为是一个多环节、多通道的快速级联反应〔11〕。脑缺血可引起神经细胞的水肿、变形、凋亡及坏死等,而该病理过程与炎症反应密切相关,脑组织炎症因子的产生不仅可引起脑胶质细胞的增生,也可参与缺血局部脑皮质微血管新生,且缺血区微血管新生程度对脑缺血患者的神经功能恢复具有重要作用〔12,13〕。血管新生是指血管内皮细胞发生分裂、分化和增殖形成新的毛细血管网的过程〔14〕。据报道,微血管新生和脑缺血后再灌注血流可通过保护血脑屏障改善脑组织能量代谢障碍,促进脑缺血后神经祖细胞的分化、增殖及神经元再生,从而改善脑缺血患者神经功能、认知功能、语言功能及运动功能等〔15〕。
TLRs介导炎性脑缺血损伤,而TLR4是一种在神经细胞中表达的膜蛋白受体,主要负责炎症反应〔16〕。多项研究表明,TLR4参与脑缺血损伤的病理过程〔17〕。本研究结果显示,siRNA-TLR4转染组缺血半影区细胞形态比模型组和阴性病毒组规则,且脑组织损伤程度降低。此外,沉默TLR4表达后,MCAO大鼠脑梗死面积及脑皮质细胞凋亡率明显降低。BMECs是脑组织微血管的组成单元,其损伤程度决定了缺血局部脑皮质微血管新生的可能性〔18〕,本研究结果表明,沉默TLR4表达可促使微血管内皮细胞增殖。VEGF和Ang是血管新生的两种相关因子,VEGF可通过促进血管内皮细胞增殖,加速新生血管的形成〔19〕;Ang具有诱导血管生成的作用,其中Ang-1能促使血管成熟,Ang-2能促使血管出芽及生长,VEGF、Ang-1和Ang-2三者相互协调共同促进血管新生〔20〕。本研究结果表明沉默TLR4表达可通过上调VEGF、Ang-1和Ang-2水平促进微血管新生。脑缺血后抑制TLR4表达,可进一步抑制TAK1和NF-κB活化,从而抑制炎症反应对脑缺血后的继发性损伤。
综上,TLR4/TAK1/NF-κB途径通过增加缺血局部脑皮质新生血管形成,从而改善脑缺血对脑组织的损伤。
猜你喜欢
昆明医科大学学报(2021年3期)2021-07-22
世界最新医学信息文摘(2021年12期)2021-06-09
世界科学技术-中医药现代化(2021年10期)2021-03-02
小资CHIC!ELEGANCE(2021年46期)2021-01-11
昆明医科大学学报(2020年11期)2020-12-28
睿士(2020年11期)2020-11-16
中成药(2018年4期)2018-04-26
中国卫生标准管理(2015年2期)2016-01-14
中国康复理论与实践(2015年10期)2015-12-24
中国体外循环杂志(2015年3期)2015-12-08
