
FKBP38可调控多巴胺能神经元的凋亡
2023-01-05 08:11:26谢彩婷封文斌赵文卓李芳红吴晓丽赵子建
中国药理学通报 2023年1期
刘 静,谢彩婷,封文斌,赵文卓,李芳红,吴晓丽,赵子建
(广东工业大学生物医药学院,广东 广州 510006)
帕金森病(Parkinson’s disease PD)又称为震颤麻痹,是一种影响患者活动能力的中枢神经系统慢性疾病,起病隐袭,进展缓慢[1]。其主要病理特征是中脑黑质致密部多巴胺能神经元损伤,神经元内出现以α-突触核蛋白(α-synuclein)为关键成分的路易小体(Lewy body),进而引起神经元变性、凋亡,导致脑部多巴胺水平降低[2]。文献研究表明,减少多巴胺神经元凋亡可一定程度改善PD的症状。目前临床上用于治疗PD的药物(司来吉兰和美多巴)均是通过提高多巴胺水平,从而减少神经元凋亡。然而,这些药物只能改善PD症状,并不能阻止病情的进展,也无法治愈疾病[1-3]。因此,寻找新的靶点,在PD发病早期进行基因修饰,抑制神经元凋亡,进而改善PD症状,是极为迫切的事情。
FK506结合蛋白38 (FKBP38)是FKBP家族蛋白之一,特征在于其相应的 mRNA 在人脑组织中显著表达[4],包含FK506结合域并表现出肽基脯氨酰异构酶活性[5-6]。在小鼠胚胎发育过程中,FKBP38可通过拮抗Shh通路延缓神经管的关闭[7-8]。并且FKBP38的丢失还会导致后神经管细胞凋亡增加和发育异常[9]。此外,还有研究表明FKBP38通过其C端跨膜结构域锚定在线粒体外膜上,招募并帮助抗凋亡蛋白Bcl-2来抑制细胞凋亡[10-11]。这提示FKBP38可能是一种重要的细胞凋亡抑制剂[12]。但是,尚未有研究报道FKBP38在帕金森病中对多巴胺能神经元的作用。
因此,本文旨在探讨在PD体内外模型中FKBP38蛋白表达情况,并选用神经毒素诱导的PD体外模型,研究过表达和敲减FKBP38后,对多巴胺能神经元凋亡的作用。
1 材料与方法
1.1 材料
1.1.1实验动物 8周龄♂的C57BL/6小鼠30只,体质量(20~25) g,购买于广东省医学实验动物中心(广东佛山,编号:44007200085200)。小鼠每笼5只,饲养温度(21~25) ℃,12 h昼夜交替,自由饮水摄食,每周更换3~4次垫料,动物在开展实验前先适应性喂养1周。
1.1.2细胞和慢病毒 MN9D(小鼠中脑多巴胺神经元)细胞购自广州吉尼欧公司。FKBP38慢病毒载体购自广州赛业公司,慢病毒载体为LV-EFS>mFKbP8-PGK>Puro 和LV-CMV>eGFP/T2A/Puro(以下分别简写为OV-FKBP38及OV-NC),LV-U6>mFKbP8[shRNA#2]-PGK>Puro和LV-U6 >Scramble-shRNA-PGK>EGFP/T2A/Puro(以下分别简写为sh-NC及sh-FKBP38),该慢病毒中含有FKBP38全长cDNA,所有结构都经DNA测序验证,转染效果经免疫印迹(Western blot)验证。
1.1.3试剂 RPMI1640(Lot:PM150110A)购自武汉普诺赛公司;胎牛血清购自于美国Gibco公司;胰酶(Lot:2120649)购买于美国Gibco公司;CCK-8(cell counting kit-8,批号:G909FA003)购自上海生物工程有限公司;DMSO购自于美国sigma aldrich公司;FKBP38抗体购自R&D 公司;α-synuclein、Tom20和Bax抗体购自美国CST公司;Bcl-2抗体购自英国Abcam公司;TH抗体购买于美国Gibco公司;鱼藤酮和MPTP购自中国阿拉丁公司;MPP+购买于美国Sigma公司。
1.1.4仪器 二氧化碳培养箱CLM-170B-8-NF型(ESCO,新加坡);二级生物安全柜AC2-4S1型(ESCO,新加坡);EPED-2TF纯水仪(中国);DYY-6C电泳仪(中国);多管架自动平衡离心机TDZ5-WS型(上海卢湘仪离心机仪器有限公司);流式细胞仪(DXP AthenaTM,美国);Leica倒置显微镜(Leica,德国);酶标仪CMAXPLUS+ (美国);化学发光凝胶成像系统ChemiDoc+XRS(Bio-Rad,美国)。
1.2 方法
1.2.1小鼠分组及给药 小鼠随机分为2组,每组 15 只:对照组、模型组。以腹腔注射的方式给予模型组小鼠 10 mg·kg-1的MPTP,给予对照组小鼠相同体积的PBS。3 d注射1次,连续注射7次,在末次注射后的d 2进行脱颈椎处死小鼠,立即取脑,冰上操作,剥离纹状体和黑质,液氮速冻,后转至-80 ℃保存备用。
根据R型因子分析及R型聚类分析结果,结合全区地质、地球化学特征以及研究区现有矿化线索,将分析的20种元素进行划分,确定组合异常5个分别为Cr,Ni,Co组合,La,Nb,Th,U,Y组合,Ag,Cd,Cu,Pb,Zn组合,W,Sn,Mo,Bi组合及Au,As,Sb组合。
1.2.2细胞培养及药物处理 MN9D细胞用添加10%胎牛血清的RPMI 1640培养基,于37 ℃、5%二氧化碳细胞培养箱中进行常规培养,每隔1天进行换液,2~3 d进行传代,待细胞密度为85%左右,根据不同条件进行刺激。鱼藤酮及MPP+采用完全培养基溶解,刺激细胞时采取全换液的方式进行。
1.2.3细胞分组及感染 将MN9D细胞分为FKBP38过表达组(OV-FKBP38)及对照组(OV-NC)、敲减FKBP38组(sh-FKBP38)及对照组(sh-NC),将细胞铺板接种于6孔板,每孔接种细胞数2×105个,放置于培养箱常规培养。待细胞密度长于30%~50%时进行感染慢病毒实验,收集细胞提蛋白进行Western blot检验慢病毒感染效率。
1.2.4CCK-8法检测细胞增殖活性 将生长状态良好的MN9D细胞及转染慢病毒成功后的细胞制备成单细胞悬液并铺板接种于96孔板,每孔100 μL细胞悬液,每孔接种细胞数2×104个,放置于培养箱常规培养。实验分组为不同浓度(8、4、2、1、0.5、0.25、0.125、0)μmol·L-1的鱼藤酮处理MN9D组,及同浓度鱼藤酮分别处理OV-NC、OV-FKBP38、sh-NC、sh-FKBP38组。过夜培养待细胞贴壁后,吸出原培养基,更换为提前配好不同浓度鱼藤酮药物的培养基;继续放置培养箱培养不同时间后,向每孔添加10 μL CCK-8溶液,贴壁加入以免产生气泡,避光孵育,1~2 h后取出,用酶标仪测定在450 nm处的吸光度(A值)。
1.2.5Western blot检测蛋白水平 在10 cm细胞培养皿中加入200 μL RIPA裂解液,裂解液含(RIPA、PMSF、蛋白酶抑制剂及磷酸酶抑制剂),用无菌细胞刮缓慢刮下,置于冰上裂解10 min,超声破碎1~2次,每次30 s,4 ℃离心机12 000 r·min-1离心20 min后,取上清于另一干净Ep管中,蛋白定量 BCA 法检测待测样品浓度。待测样品与5×上样缓冲液稀释混合,金属浴10 min。取同等质量蛋白样品在质量分数10%或12%聚丙烯酰胺凝胶电泳(SDS-PAGE)进行电泳,恒压转膜至PVDF膜,室温下用5%脱脂奶粉封闭1 h或4 ℃过夜封闭。分别加一抗α-syn(1 ∶1 000)、FKBP38(1 ∶500)、Bax(1 ∶1 000)、TH(1 ∶1 000)、Tom20(1 ∶1 000)、Bcl-2(1 ∶2000) 、 β-actin(1 ∶5 000),4 ℃孵育过夜。过夜孵育后TBST洗膜3次,二抗室温孵育1 h,再次TBST洗膜3次,ECL发光成像仪曝光。并应用ImageJ软件对目标蛋白和内参蛋白灰度值进行分析,相对表达量=目的蛋白灰度值/内参灰度值。
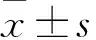
2 结果
2.1 PD体内模型中FKBP38表达情况如Fig 1所示,通过Western blot检测MPTP诱导的PD体内模型。结果显示,在小鼠脑纹状体和黑质区域,α-synuclein蛋白的表达量水平明显升高(P<0.01),TH表达量水平明显下降(P<0.01)(Fig 1A)。而在MPTP诱导的PD小鼠脑纹状体区域中FKBP38表达水平明显下降(P<0.01),在黑质区域中FKBP38表达量无明显变化(Fig 1B),说明在MPTP诱导的PD体内模型中FKBP38在纹状体中特异性下调,提示FKBP38与PD模型中的特异性蛋白(α-synuclein和TH)存在相关性。
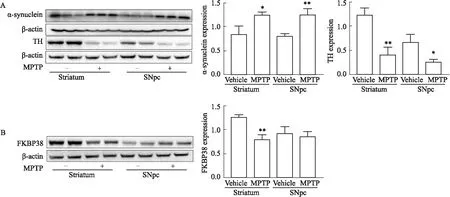
Fig 1 Expression of FKBP38 protein in striatum and substantia nigra of PD model

Fig 2 Expression of FKBP38 in PD cell model A:α-synuclein/TH expression in PD cells; B:Tom20/FKBP38 expression in PD cells. *P<0.05,**P<0.01 vs control
2.3 稳定过表达FKBP38的MN9D细胞株的建立如Fig 3所示,利用携带FKBP38的慢病毒感染MN9D细胞。随后,我们利用Western blot检测FKBP38 的表达水平。与MN9D组相比,OV-NC组和sh-NC组的FKBP38表达量均没有明显变化,说明选择的载体合适。与OV-NC组相比,OV-FKBP38组的FKBP38表达量明显上升(P<0.05),说明FKBP38过表达细胞构建成功。与sh-NC组相比,sh-FKBP38组的FKBP38表达量明显下降(P<0.05),说明FKBP38敲减细胞构建成功。
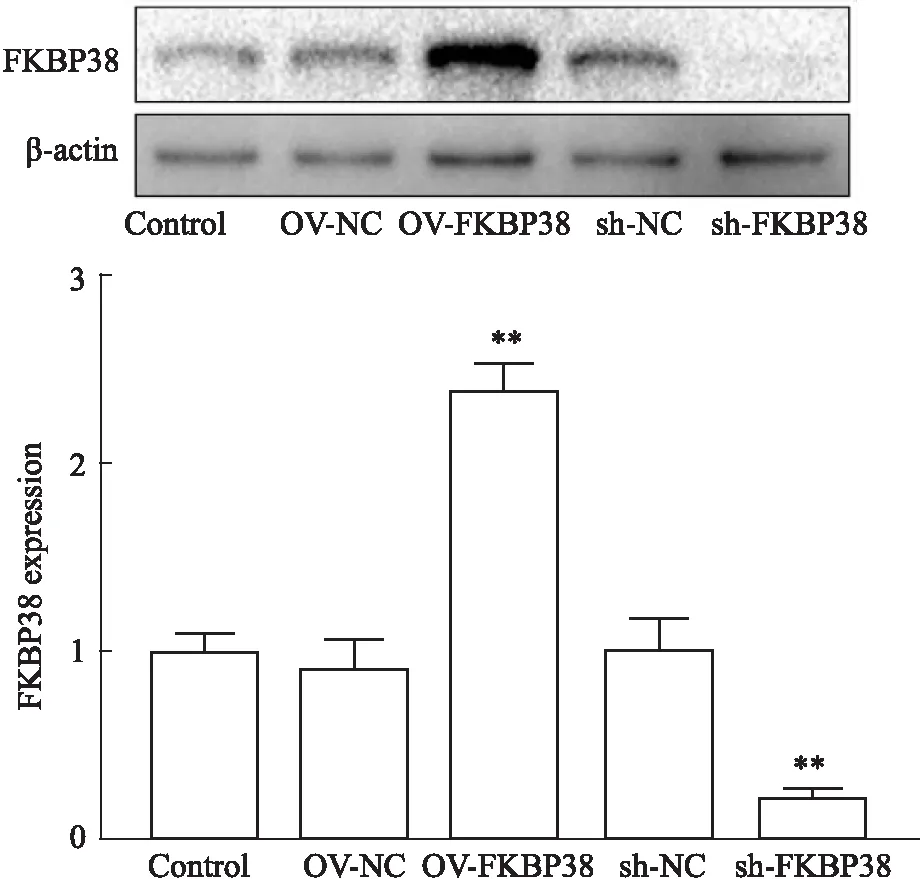
Fig 3 FKBP38 lentiviral transfection of MN9D cells
2.4 鱼藤酮抑制MN9D细胞活力如Fig 4所示,CCK-8检测不同浓度 (8、4、2、1、0.5、0.25、0.125、0)μmol·L-1的鱼藤酮处理MN9D细胞8 h后,结果显示,显著抑制细胞增殖(Fig 4A)(P<0.01),4 μmol·L-1浓度时,细胞活力下降到对照组的49.52%,且用鱼藤酮(4 μmol·L-1)处理MN9D细胞不同时间,发现随着处理时间的增加,细胞活力降低加剧(Fig 4B)(P<0.01)处理8 h时,细胞活力为对照组50.08%。因此,选用鱼藤酮(4 μmol·L-1)处理MN9D细胞8 h进行后续研究。
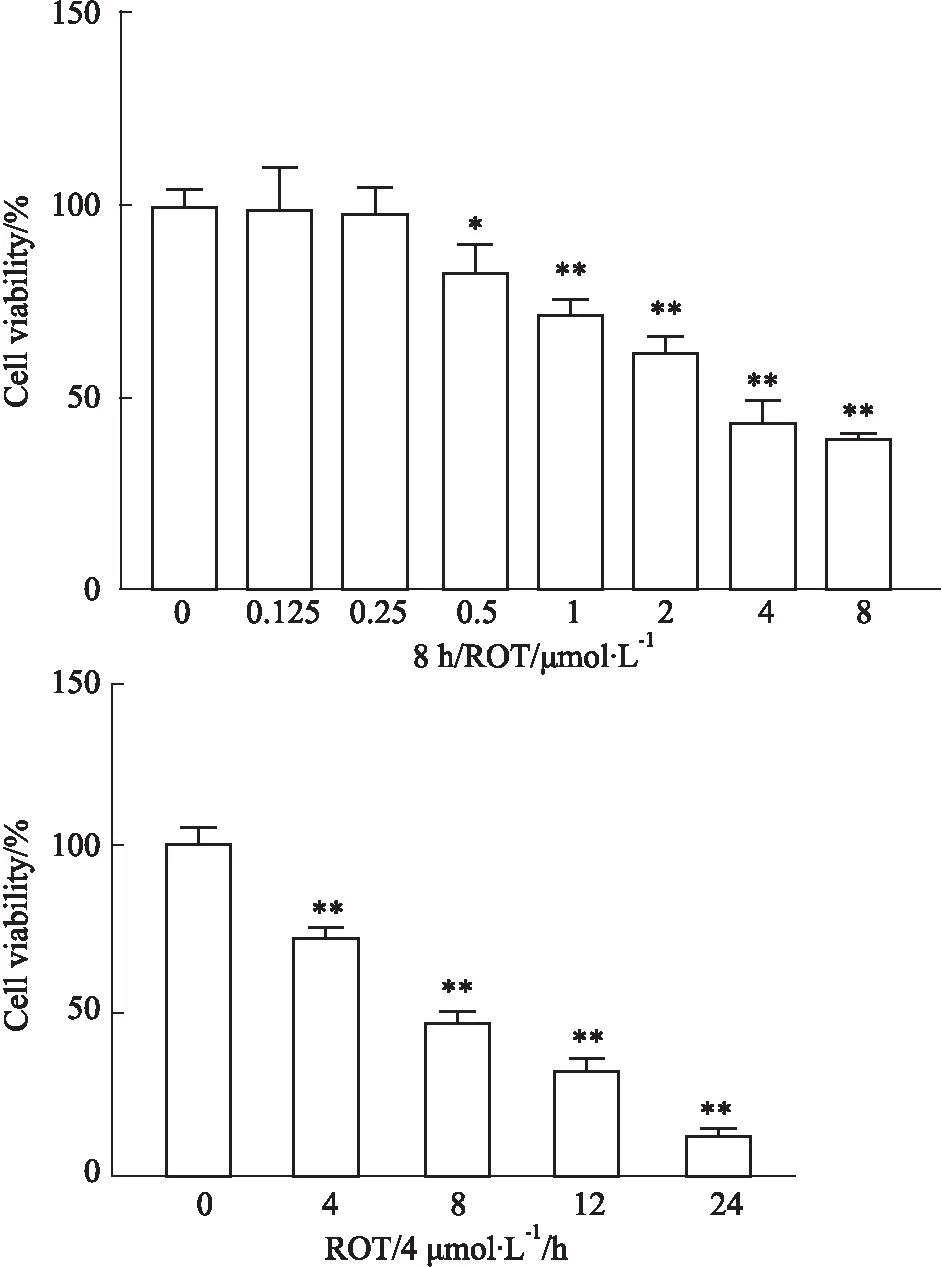
Fig 4 Effect of rotenone on viability of MN9D cells
2.5 FKBP38对PD细胞活力下降的影响如Fig 5所示,鱼藤酮处理MN9D细胞8 h后,与sh-NC组模型组相比,sh-FKBP38组模型组的细胞活力明显下降(Fig 5A)(P<0.01),表明FKBP38敲减后加剧PD细胞活力下降。而与OV-NC组模型组相比,OV- FKBP38组模型组的细胞活力明显增强(Fig 5B)(P<0.01),表明过表达FKBP38可以明显改善PD细胞活力下降。
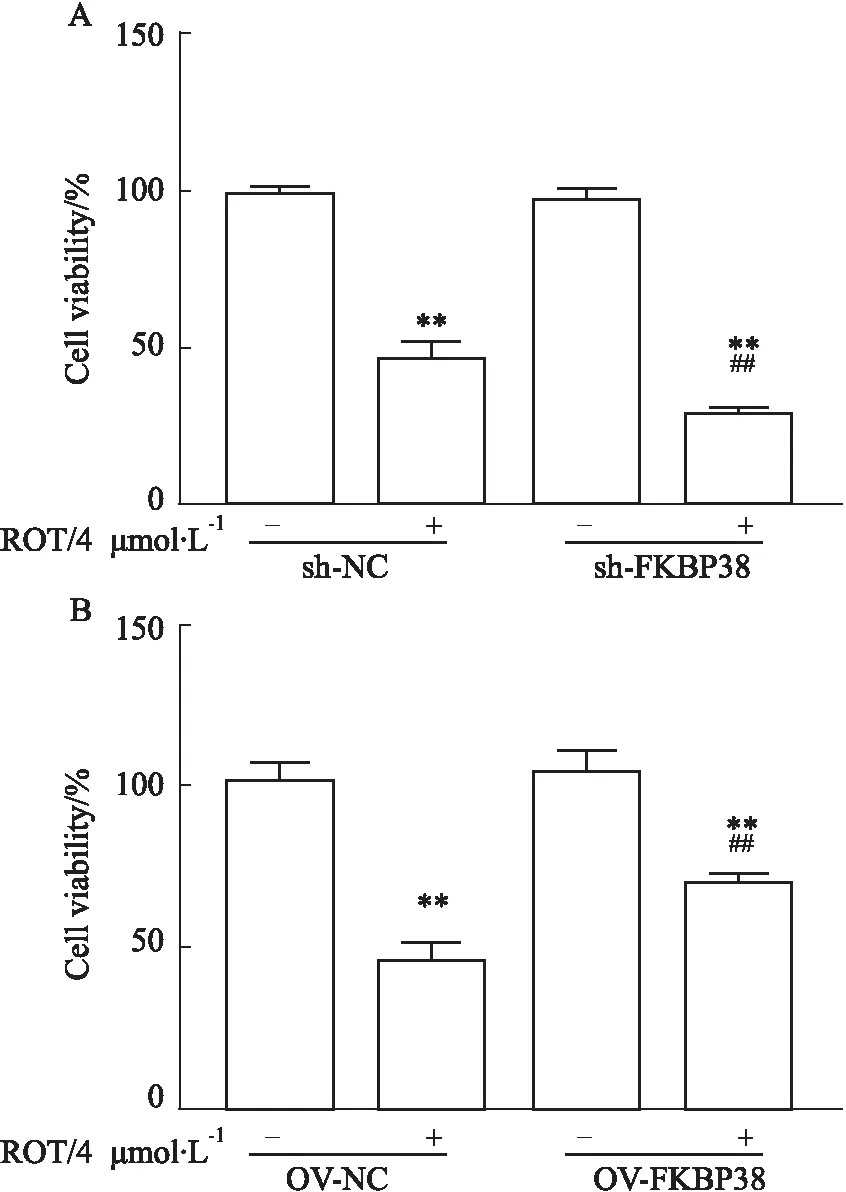
Fig 5 Effect of FKBP38 on decreased viability
2.6 FKBP38对PD细胞凋亡的影响如Fig 6所示,Western blot 检测结果显示,与对照组相比,模型组经鱼藤酮处理MN9D细胞8 h后,抗凋亡蛋白Bcl-2的表达明显下调(P<0.05),凋亡蛋白Bax的表达没有差异,但Bcl-2/Bax比值降低(P<0.05),表明鱼藤酮引起了多巴胺能神经元的凋亡。与OV-NC组模型组相比,OV-FKBP38组模型组的Bcl-2蛋白表达上升(P<0.01),Bax的表达量没有差异,Bcl-2/Bax比值上升(P<0.01),说明过表达FKBP38可以明显改善鱼藤酮引起的多巴胺能神经元凋亡。与sh-NC组模型组相比,sh-FKBP38组模型组的Bcl-2和Bax的表达量均无明显差异,表明敲减FKBP38无明显加剧鱼藤酮引起的多巴胺能神经元凋亡。
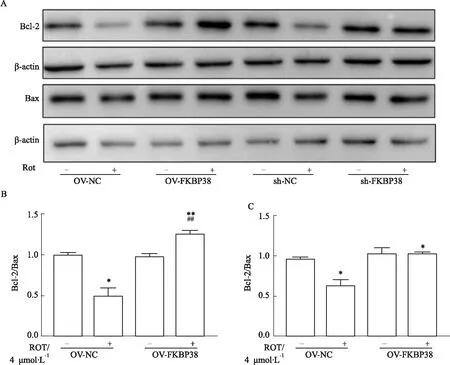
Fig 6 Effect of FKBP38 over expression on expression of Bcl-2,Bax protein in PD
3 讨论
帕金森病是一种进展非常缓慢的疾病,发病因素复杂,且不同患者病情轻重也存在很大的差异[13]。多巴胺是一种神经递质,将信息从一个神经细胞传至另一个神经细胞,保证神经的传导性,由多巴胺能神经元合成并储存在囊泡中,而后通过胞裂外排的方式由神经元释放,多巴胺能神经元的病变会导致帕金森病。这也是临床上导致PD的主要病理特征,减少神经元凋亡对缓解PD进程有重要作用[14]。目前,关于帕金森病治疗手段只能延缓但无法阻止疾病的进展[15]。基因治疗是帕金森病治疗的新兴领域,由于其发病机制复杂,其中线粒体功能障碍在帕金森病发生中具有重要作用[16-17]。因此以线粒体相关基因为靶点寻找新的治疗方法具有重要意义。同时,应深入研究线粒体功能障碍分子机制,明确线粒体相关基因对帕金森病的影响。
为研究FKBP38对神经毒素诱导的帕金森病的影响,本课题组检测在PD体内外模型中FKBP38表达水平,实验结果发现,在PD体内模型中,与正常组相比模型组小鼠脑纹状体区域中FKBP38表达水平明显降低。同样,在PD细胞模型中,与正常组相比FKBP38表达水平明显特异性降低。猜测高表达FKBP38可能会延缓多巴胺神经元凋亡从而缓解PD进程。为进一步探究FKBP38对帕金森病的影响,我们在体外实验中通过使用FKBP38慢病毒转染MN9D细胞来检测其表型变化。实验结果显示,鱼藤酮可明显降低MN9D细胞活力,并通过降低Bcl-2的表达量引起MN9D细胞凋亡。而过表达FKBP38后能明显改善鱼藤酮引起的细胞活力下降,表明过表达FKBP38可明显提高PD细胞活力。过表达FKBP38后,抗凋亡蛋白Bcl-2的表达量明显升高,Bcl-2/Bax比值明显升高,表明过表达FKBP38改善鱼藤酮引起的多巴胺能神经元凋亡可能是通过线粒体内源途径 Bcl-2/Bax 通路诱导的。说明过表达FKBP38在帕金森病的发展过程中发挥着抑制多巴胺能神经元凋亡的作用。这与先前研究中FKBP38锚定在线粒体外膜招募Bcl-2从而抑制凋亡作用相一致[11]。
综上所述,FKBP38表达在鱼藤酮诱导的PD细胞模型被特异性下调,敲减FKBP38加剧鱼藤酮诱导的细胞损伤,通过过表达FKBP38能够明显恢复鱼藤酮诱导的细胞活力下降,并抑制鱼藤酮引起的细胞凋亡。提示FKBP38与多巴胺神经元凋亡相关,有望成为治疗或者减缓帕金森病进程的重要靶点。
猜你喜欢
欣漾(2024年2期)2024-04-27 12:03:09
疯狂英语·新悦读(2023年2期)2023-10-12 14:42:47
保健医苑(2022年6期)2022-07-08 01:25:28
疯狂英语·新读写(2020年3期)2020-06-06 09:05:58
时代英语·高一(2019年5期)2019-09-03 02:09:34
活力(2019年22期)2019-03-16 12:47:04
深圳职业技术学院学报(2018年3期)2018-07-23 06:42:18
中国野生植物资源(2018年1期)2018-04-23 07:09:01
医学研究杂志(2015年5期)2015-06-10 06:43:26
中国中医药现代远程教育(2014年11期)2014-08-08 13:23:44
