
Fine root morphology and soil properties under influence of different tree stands along an altitudinal climosequence in the Carpathian mountains
2023-01-05 12:37KarolinaStaszelEwanskaJarosawLasota
Forest Ecosystems 2022年5期
Karolina Staszel,Ewa Bło′nska,Jarosław Lasota
Department of Ecology and Silviculture,Faculty of Forestry,University of Agriculture in Krakow,29 Listopada 46 Str,31-425,Krak′ow,Poland
Keywords:Beech Fir Forest ecosystem Root biomass Soil properties
A B S T R A C T In an era of climate change,understanding the factors that impact root systems can improve our understanding of carbon cycling in forest ecosystems.The study objective was to determine the impact of climatic conditions on the biomass and morphology of roots of different tree species along an elevation gradient,and consequently on properties of montane forest soils.The study plots were established at three different elevations(600,800 and 1000 m a.s.l.)along a slope with an inclination of 15°.The research plots were located in a beech stand(Fagus sylvatica L.)and fir stand(Abies alba Mill.).Soil samples were collected from each study plot,for which basic physical and chemical properties were determined.Additionally,we determined the morphology,production and decomposition rate of fine roots.Our analyses confirmed the significance of climatic conditions in the formation of soil properties,in particular the amount of accumulated carbon and nitrogen content.A decrease of root biomass and reduced root growth were recorded with increasing elevation.The characteristics of roots were linked with the properties of the studied mountain soils,in particular pH,alkaline cation content and content of selected micronutrients.Limitation of root growth in higher elevations affected both study species.Additional research into the formation of tree root morphology is needed,especially in mountainous regions where changes may occur more dynamically.This will provide a better understanding of how stands can cope with different types of environmental stress.
1.Introduction
Montane climate has a specific character,which is linked to the terrain and elevation above sea level(a.s.l.).With increasing elevation,the temperature and humidity change;the climate becomes more harsh,cool and humid.In addition,mountainous areas are characterised by long-lasting snow cover and low annual temperature amplitude.The vegetation affecting soil properties also changes along an elevation gradient(Staszel et al.,2021).Climatic conditions on mountains(temperature,humidity),and the quantities and diversity of microorganisms and organism inputs(plant litter,roots)have a major significance for the creation of soil properties(Bu et al.,2012;Wang et al.,2017;Zhang et al.,2021).These factors subsequently determine the efficiency and stability of ecosystems,which are controlled by precipitation,air temperature and light intensity(Morales et al.,2007).Plants and soil,despite being different components of the biogeochemical cycle,are strictly interlinked and interact(Liu et al.,2021).According to Zhang et al.(2021),vegetation type and soil pH might be the main factors controlling the spatial variability of soil organic carbon(SOC)and total nitrogen in mountain ecosystems.The composition of plant assemblages,in particular the number of plant species,changes as a result of climatic limitations.The number of trees,shrubs and undergrowth plants is reduced with increasing elevation,which translates to the activity of soil microorganisms(Klimek et al.,2020).In addition,the growth and biomass of fine roots decreases with the altitudinal gradient,which could be related to rainfall or nutrient availability(Sierra Cornejo et al.,2020).At lower elevations in montane areas which have favourable climatic conditions with a longer vegetative period plants produce greater amounts of biomass,which supplies the upper soil layer,whereas at higher elevations a decrease in biomass is observed,resulting in a lower influx of detritus to soil(Kotas et al.,2018).
Soil properties may determine a number of the features of root systems.They significantly affect the possibility of development of root systems,especially by mechanical resistance.The presence of highly rocky or dense soil horizons usually results in less root penetration into the soil.Apart from the graining characteristics,the water and air content in the soil significantly affect the ability to penetrate by the roots(Crow,2005).Among the chemical characteristics of soil,acidification especially the presence of toxic aluminium and the availability of phosphorus exerts the strongest influence on the development of plant root systems(Persson et al.,1995;Lockwood et al.,2003;Robles-Aguilar et al.,2019).Fahey et al.(2005)estimated C input to soil in the Hubbard Brook northern hardwood forest from fine and woody roots to be about 90 and 40 g C⋅m-2⋅yr-1,respectively.Using these estimates,the ratio of aboveground to belowground detrital C input to soil is roughly 1.6.According to Joslin and Henderson(1987),the amount of residue reaching soil from root decay is five times greater than from plant litter fall.The roots are one of the more important components of plant biomass,thanks to which vegetation can control the intake of water and nutrients while also impacting biogeochemical cycles.Despite their minor size,fine roots constitute the main source of carbon in soil due to annual root dieback(Jackson et al.,1997;Jones et al.,2004).In addition,the amount of nutrients that are released by the death of fine roots is much greater than that released by the decomposition of plant litter decay(Yuan and Chen,2010).Depending on the soil type,roots comprise approximately 30%of its biomass,but also 40%–85%of net primary production(Hoffmann and Usoltsev,2001).In times of changing climatic conditions,trees have to adapt to the environment surrounding them.To this end,they use either extensive or adaptive strategies,as appropriate,which differ according to species,climate factors like temperature and precipitation,and also soil properties(Fin′er et al.,2011).In the case of an extensive strategy,biomass and the length of fine roots are increased,and this adaptation enables growth even under difficult conditions(Ostonen et al.,2007).This also impacts the distribution of roots in the soil profile,which consequently greatly influences the cycles of carbon and nitrogen in soil.Stress situations associated with excessive drought or damage can be reflected in the chemical and morphological characters of roots,which include specific root length(SRL),root tissue density(RTD)and root diameter(Weemstra et al.,2016).In response to stress factors,roots become more plastic,allowing them to seek more nutrients more easily and quickly,which will translate into faster growth and productivity of the tree(Hodge,2004;Kembel and Cahill,2005).Detailed knowledge of the factors influencing root biomass and morphology will allow the cultivation of stands that are more stable and more resistant to change.The detailed response of the root systems of various temperate forest species to climate change is currently unknown.
Knowledge on the factors affecting the root systems of different tree species is important for a better understanding of the carbon cycle in forest ecosystems.Our study utilised climatic sequence in a mountain ecosystem for the monitoring of the effect of climatic conditions on the creation of biomass and root morphology.The objective of our study was to determine the biomass and morphology of the roots of beech and fir tree stands growing in different climatic conditions.An additional study objective was to determine the growth of roots and the rate of root decomposition in diverse climatic conditions.The following research hypotheses were tested:i)along with the height along the elevation gradient,the climatic conditions and soil properties change and,as a consequence,the rate of decomposition processes of organic debris in the soil changes;ii)the location along the elevation gradient is important in shaping the morphological features of tree roots;iii)properties of montane forest soils,such as acidity and quality of organic matter,are related to the root characteristics of the studied tree species.
2.Materials and methods
2.1.Study area and soil sampling
The study was conducted in the Jałowiec Massif(49°38′N,19°20′E)in the Western Beskidy Mountains,in the southern part of Poland.The study took place in Magurska Nappe on the sandstone and shales,with domination of Cambisols(WRB,2014).The study plots were established at three different elevations(600,800 and 1,000 m a.s.l.)along a slope with an inclination of 15°.The tested soils were characterised by a similar texture(average sand content was 54%,silt 42% and clay 3%).The average temperature of the growing season for the study plots was 12.4°C at 600 m a.s.l.,11.3°C at 800 m a.s.l.and 10.2°C at an elevation of 1,000 m a.s.l.Average soil moisture at 600,800 and 1,000 m a.s.l.differed,at 22.74%,29.60%and 34.10%,respectively.The research plots were located in a beech stand(Fagus sylvaticaL.)and fir stand(Abies albaMill.),both at the age of 60.In each variant of elevation,3 research plots(0.10 ha)were designated separately for each species.The samples were taken after removing the litter from a depth of 0–15 cm.Three soil samples from different locations were collected on each plot.In total,54 soil samples were collected for the analysis.The field study was realised in 2021.
2.2.Root analysis
On each study plot,soil samples with a known volume of 15 cm×15 cm×15 cm were collected in three replications to determine root biomass,from which coarse roots with diameter>2 mm and fine roots with diameter<2 mm were separated.The extracted root system fragments were scanned at 400 dpi resolution and then analysed using a WinRhizo™Pro 2003b image analysis system(Regent Instruments Inc.,Ville de Qu′ebec,QC,Canada)to determine diameter,length and root area.Air-dried roots were further desiccated at 70°C for 24 h to constant weight and then weighed.Root tissue density(RTD)(kg⋅m-3),specific root area(SRA)(m2⋅kg-1),and specific root length(SRL)(m⋅g-1)were calculated according to Ostonen et al.(1999).Following the analysis of the roots,they were used for the determination of the root decomposition rate in the later stage of the experiment.Bags(15 cm×20 cm)containing 10 g of root matter were prepared for the determination of root decomposition.Bags with roots were buried to a depth of 10 cm in each study area for the period from May to October.After this period,the root litter mass loss in the bags was determined.The annual fine root(diameter<2.0 mm)biomass increase(FRBI)was determined with the core method(B¨ohm,1985).The experiment sought to determine the root production between May and October 2021.
2.3.Laboratory analysis
Air-dried soil samples were sieved through a 2-mm mesh.Physical and chemical properties were determined in these prepared samples(Ostrowska et al.,1991).Soil pH was determined by the potentiometric method in water and 1 mol⋅L-1KCl.Hydrolytic acidity(Y)was determined by the Kappen method.Total nitrogen and carbon content was determined using a LECO CNS True Mac Analyser(Leco,St.Joseph,MI,USA).The amount of alkaline cations(Ca2+,Mg2+,K+,Na+)in 1 mol⋅L-1ammonium acetate was determined by an ICP-OES(iCAP 6500 DUO,Thermo Fisher Scientific,Cambridge,UK).The Cd,Cr,Cu,Ni,Pb and Zn contents were determined after mineralisation in a 2:1 mixture of concentrated nitric and perchloric acids by ICP-OES.The gravimetric method was used to determine soil moisture.The soil samples were dried at 105°C for 48 h and weighed after drying.The humidity was determined according to Eq.(1):

where Mw is the weight of moisture,Mn is the weight of the fresh sample,and Md is the weight of the dried sample.
2.4.Statistical analysis
Spearman correlation coefficients were computed to determine the mutual relationship between individual root parameters and soil properties.Principal component analysis(PCA)was used to evaluate relationships between soil and root characteristics.A general linear model(GLM)was used to investigate the effects of elevation and different stands and the interactive effect of these two factors on fine root biomass and morphology.Tukey's test was used to evaluate differences between mean trait values.The results were considered to be statistically significant at α<0.05.All statistical analyses were performed using R statistical software(R Core Team,2021),RStudio(RStudio Team,2020),and Statistica 10 software(2010).
3.Results
3.1.Root analysis
In the case of beech,longer roots were observed compared to fir(Fig.1a).The studied tree species differed statistically significantly in root system length.The length of the beech root systems ranged from 3,957.80 to 6,452.58 cm,and fir from 2,269.63 to 2,475.62 cm at an elevation of 1,000 and 600 m a.s.l.,respectively.A tendency for reduced root length with increasing elevation could be seen for both species.Root diameter in the case of fir increased with elevation from 0.77 to 0.93 mm,but these differences were not statistically significant(Fig.1b).Statistically significant differences in root diameter between species were recorded at elevations of 800 and 1,000 m a.s.l.Root biomass for beech and fir did not change statistically significantly with increasing elevation(Fig.1c).Root biomass differed significantly between the tested tree species only at an elevation of 600 m a.s.l.In the case of SRA,significant differences between the study species were recorded at an elevation of 1,000 m a.s.l.,with an average of 7.34 m2⋅kg-1for fir and 11.70 m2⋅kg-1for beech.Fir SRA did not change significantly along the elevation gradient.The highest beech SRA was recorded at an elevation of 1,000 m a.s.l.(Fig.2a).RTD and SRL of both species did not change significantly along the elevation gradient(Fig.2b and c).At an elevation of 600 m a.s.l.,significant differences of RTD,and at 1,000 m a.s.l.,significant differences of SRL,were recorded between beech and fir.The study species did not differ significantly in root biomass,irrespective of location along the elevation gradient(Fig.3a).In the case of fir,no statistically significant differences in root biomass were determined along the elevation gradient.The lowest beech root biomass was recorded at the highest locations;the average varied from 3.27 to 5.66 g⋅dm-3.No significant differences resulting from the effect of species and elevation could be recorded for the decomposition rate(Fig.3b).At 800 m a.s.l.,the values oscillated from 24.38%to 29.24%for beech and from 19.48%to 31.35% for fir.In the case of root growth,statistically significant differences were recorded at 600 and 1,000 m a.s.l.(Fig.3c).The root growth of beech did not change significantly along the elevation gradient;for fir,significantly greater growth was recorded at 800 m a.s.l.GLM analysis confirmed the significance of location along the elevation gradient on root biomass and root growth of the examined tree species(Table 1).Tree species was significant for formation of length and diameter of roots,SRL and root production.The simultaneous importance of tree species and location along the elevation gradient was recorded for root weight(Table 1).
3.2.Soil properties
The conducted analyses indicate differences in the physicochemical properties resulting from the effect of different tree species and location along the elevation gradient(Tables 2 and 3).At an elevation of 600 m a.s.l.,statistically significant differences in nitrogen(N)content were recorded between the study species.At this elevation,soils under the effect of beech were characterised by higher N content compared to soils under fir influence(Table 2).At an elevation of 800 m a.s.l.,the differences between the study species were recorded for pH in KCl,C and N content,C/N ratio,and Mg,K and Na content.At 1,000 m a.s.l.,no differences between the tested species were recorded other than for Ca content(Table 2).Significant differences between the study species were also recorded for micronutrient content(Table 3).At an elevation of 600 m a.s.l.,differences between species in the content of Mn were recorded;at 800 m a.s.l.,in Co,Fe,Mn and Pb content,and at 1,000 m a.s.l.,in Cd,Co,Cu,Mn,Ni and Pb content.Soils at the lowest elevation were characterised by different properties in comparison to soils at elevations of 800 and 1,000 m a.s.l.(Table 2).At 600 m a.s.l.,soils were characterised by significantly higher pH,lower acidity,higher content of alkaline cations and lower C/N ratio in comparison with soils at 800 and 1,000 m a.s.l.,irrespective of species.Significantly higher C and N content was recorded for soils at an elevation of 1,000 m a.s.l.,irrespective of species.In the case of micronutrients,a less pronounced significance of location along the elevation gradient was recorded(Table 3).
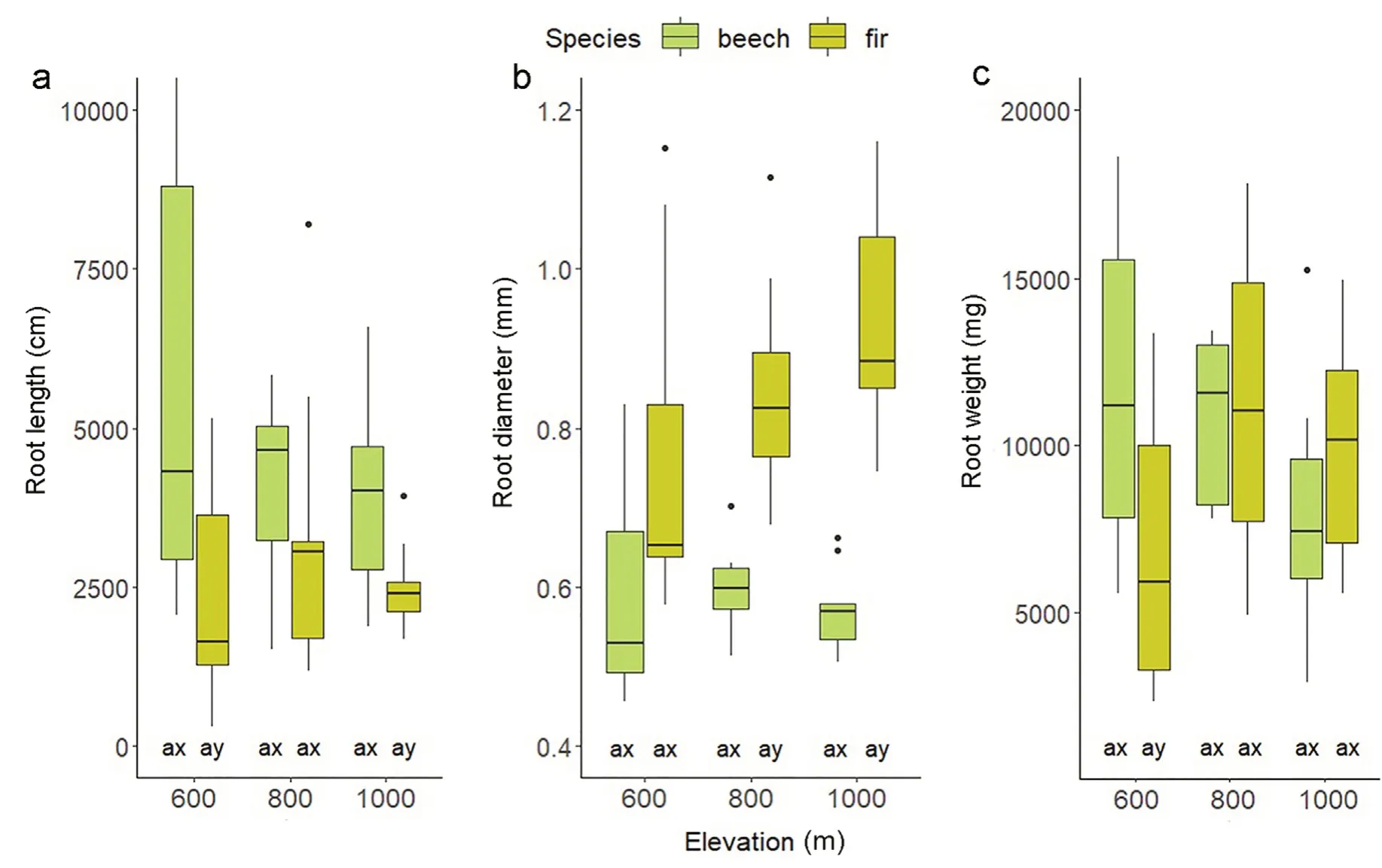
Fig.1.Root length(cm),root diameter(mm)and root weight(mg)at different elevations.Different lowercase letters(a,b,c)indicate significant parameter differences between different heights,(x,y)indicate significant parameter differences between species;TukeyHSD p<0.05(box-whisker plots with median,25-and 75-percentiles,and extremes).

Fig.2.Change specific root area(SRA m2⋅kg-1),root tissue density(RTD kg⋅m-3)and specific root length(SRL m⋅g-1)at different elevations.Different lowercase letters(x,y)indicate significant parameter differences between species,letters(a,b,c)indicate significant parameter differences between different heights;TukeyHSD p<0.05.
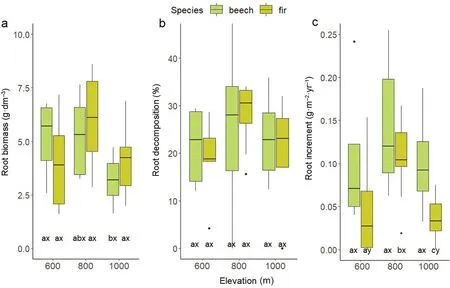
Fig.3.Root biomass(g⋅dm-3),root decomposition(%)and root increment(g⋅m-2⋅yr-1)at different elevations.Different lowercase letters(x,y)indicate significant parameter differences between species,letters(a,b,c)indicate significant parameter differences between different heights;TukeyHSD p<0.05.

Table 1Summary of GLM analysis for root parameters influenced by different tree species in gradient(a.s.l.).
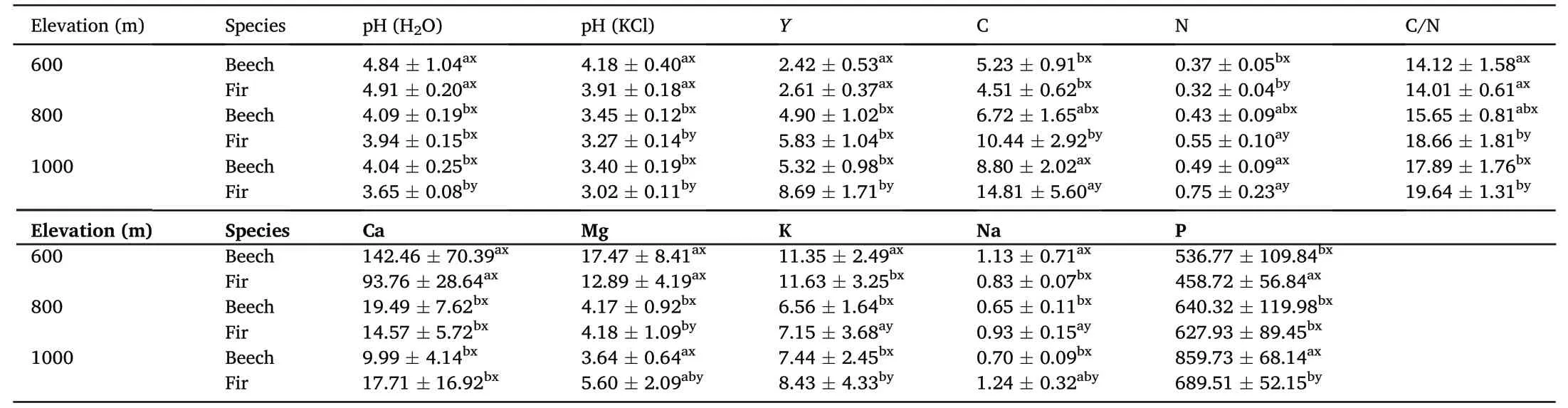
Table 2Chemical properties of study soils under influence of different species in gradient.

Table 3Microelements content(mg⋅kg-1)in study soils under influence of different species in gradient.
3.3.The relationship between root traits and soil properties
The conducted analysis confirmed the relationships between root traits and soil properties along the elevation gradient(Fig.4).Root length correlated positively with pH and content of Ca,Mg,Na,Cd,Fe,Mn,Ni and Zn.Root diameter correlated positively with hydrolytic acidity,content of C and N,and C/N ratio.Root diameter correlated negatively with pH and content of Ca,Cd,Cu,Fe,Mn,Ni and Zn.The content of Cu and Pb correlated positively with SRA and negatively with root biomass.SRL exhibited a positive correlation with pH and content of Ca,Mg,Cd,Cu,Fe,Ni and Zn.In addition,root growth correlated positively with Mn content(Fig.4).The conducted PCA analysis confirmed relationships between the parameters describing roots and soil properties.In addition,the PCA confirmed the distinction between beech and fir tree stands(Fig.5).Factors 1 and 2 explained 46.8% of variance of the tested characteristics.Factor 1 was related to the chemical properties of the tested soils and location along the gradient;Factor 2 was related to root parameters.In addition,PCA analysis confirmed the distinction between the properties of soils located at the elevations of 600 and 1,000 m a.s.l.Soils in these locations differed in acidification,C and N content and the content of alkaline cations.

Fig.4.Correlation between root characteristics and soil parameters.Significance effect*≤0.05.
4.Discussion
4.1.Soil properties
The conducted research confirmed the validity of the research hypotheses.Our analyses confirmed the significance of climatic conditions in the formation of soil properties,in particular the amount of accumulated carbon and nitrogen content.Carbon and nitrogen resources increased with elevation.Soils at the lowest elevations were characterised by significantly different physicochemical properties as compared with soils from the higher locations.Independently of tree species,at an elevation of 600 m a.s.l.soils were characterised by significantly higher pH,lower hydrolytic acidity,higher content of alkaline cations and lower C/N ratio in comparison with soils at 800 and 1,000 m a.s.l.Our results are congruent with the results of earlier studies,which demonstrated that in granite-based soils,total organic carbon stock increases with elevation a.s.l.and soil acidification increases along an elevation gradient(Bojko and Kabała,2016;Lasota et al.,2016;Staszel et al.,2021).The results obtained in the present study are likely the effect of the increasing precipitation and reduced temperature at higher locations along the elevation gradient.Greater accumulation of acidic organic matter at higher montane locations is the outcome of retardation of its decomposition,which is directly linked to low temperatures inhibiting the biological decomposition of organic residues(Prescott,2010;Klimek et al.,2020).Lower C/N at lower elevations confirmed the higher efficiency of decomposition of soil organic matter,which is directly linked with the biological activity of soils.At lower elevations the climate is milder,which creates better conditions for the growth and activity of microorganisms.In a study by Zhao et al.(2021),climate factors including MAT,MAP and their combined effects controlled the main elevation patterns of soil microorganisms.Acidification of soils located at higher elevations is also linked with the quality of soil organic matter.Soils from lower elevations are accompanied by mor-type humus,which is dominated by poorly decomposed organic residues.Mor humus is formed under the conditions of low biological activity in the soil,where organic matter mineralisation occurs at a slow rate(Lasota et al.,2020).On the other hand,Mull is a type of well-humified organic matter associated with high biological activity.Mull humus is characteristic of soils at lower elevations.More favourable thermal and humidity conditions prevail at lower locations,which affects the decomposition rate of soil organic matter.In our study,we have obtained confirmation of the results of Bojko and Kabała(2017),stating that humus forms reflect the accumulation of carbon in soils of the elevation gradient.The resources of C and N and other soil properties are not affected solely by the climate,but also by vegetation.Along the elevation gradient,differences in the amount of organic matter provided may result from the intensity and differentiation of plant layer development.It is known that the elevation gradient is one of the decisive factors forming the spatial patterns of species diversity(Lomolino,2001).In addition,the availability of light,humidity and soil depth change along the elevation gradient,leading to local differences in the species composition of plants(Cirimwami et al.,2019).An additional cause for soil acidification and lower levels of alkaline cations at higher elevations is the increase of precipitation,which affects the leaching processes(Kocowicz,1998).Our research showed the role of root systems in shaping the accumulation of soil organic carbon in the studied soils.In particular,underground biomass in the form of fine roots not only takes up nutrients but can also,through root exudates or decaying dead roots,contribute to the deposition of about 58%–96%of the total C in the soil(Shahzad et al.,2015).The content of organic acids extracted by the roots not only influences organic carbon,but also contributes to the formation of microbial processes within the rhizosphere(Badri and Vivanco,2009;Erktan et al.,2018).
4.2.Root analysis

Fig.5.Projection of variables onto the plane of the first and second PCA factors.
The conducted study also confirmed the validity of the second hypothesis,concerning the importance of location along the elevation gradient and species on the traits of root systems of trees.In this study,we were able to observe statistically significant differences in the length and diameter of roots,SRL and root growth.Independently of location,beech was characterised by the highest root length compared to fir.Our study confirmed the typical species traits of the analysed trees.The silver fir develops a strong taproot,which can develop independently of the habitat.In addition,fir develops a regular crown comprising of lateral roots,creating anchor-like branching(K¨ostler et al.,1968).The European beech initially develops a taproot,but this alters into a very dense heart-shaped root system,particularly in the direct vicinity of the trunk(K¨ostler et al.,1968).In addition,the present study demonstrated the role of species in SRL determination;higher SRL was recorded for beech.Analysis of morphological characters can provide information on the strategies of plants,their adaptation to climatic conditions and to the environment,elevation above sea level or availability of nutrients(Ostonen et al.,2007;Valverde-Barrantes et al.,2015;Freschet et al.,2017).Better characteristics of root systems in the case of beech predispose it for creating tree stands at higher elevations in mountains.Ryser(1996)noted that the response of a plant to environmental conditions results in changes to root tissue density(RTD)and diameter,which may have a contrasting effect on SRL.Changes in the morphology of roots at different elevations affect SRL and SRA.In addition,the elevation and species have a marked impact on fine root growth during the vegetation season.This relationship was also confirmed by the GLM analysis.The greatest root biomass and growth was recorded at an elevation of 800 m a.s.l.,which may be associated with more favourable humidity conditions at a sufficiently long vegetative season.Greater root growth was recorded for beech than for fir.This may have stemmed from the seasonal fine root production which,depending on the species,occurs in spring—at the beginning of the vegetative season,or at the end of it(Persson,1983;Burton et al.,2000).Research conducted in a beech forest in Italy demonstrated increased biomass and root length in mid-summer and at the beginning of autumn(Montagnoli et al.,2014).Other research on a coniferous species provided evidence that the peak of production occurred between spring and summer(Tingey et al.,2005).On the other hand,another study on the Douglas fir conducted over a period of several years also demonstrated growth in autumn and winter(Santantonio and Hermann,1985).Differences in the growth of roots between the species investigated in the present study can be explained by the study period.Our experiment on root growth was conducted from spring through autumn,with omission of the winter months,which may be of key significance for the growth of coniferous roots.
4.3.The relationship between root traits and soil properties
The conducted study confirmed the relationship between root characteristics and properties of montane forest soils.Root length correlated positively with pH,alkaline cation content and selected micronutrient content in the studied soils.Soil acidification,and its resultingly increased Al content,may directly alter root characteristics,disturb root growth and limit root functions(Vanguelova et al.,2007).′Swiątek and Pietrzykowski(2021)identified chemical properties of soils as factors affecting the biomass of fine roots.The pH values between 3.5 and 4.1 increased the growth of fine roots,whereas availability of exchangeable Mg inhibited such growth.In the present study,a reversed relationship between fine root growth and Mg content was observed.Soil richness in alkaline cations(Ca,Mg and Na)stimulated the root growth of the tested tree species(′Swiątek and Pietrzykowski,2021).According to Meier et al.(2020),on fertile soils rich in alkaline cations,the growth of fine roots of beech is intensive.Root growth can also be associated with the microbial activity of soils.Acidic soils with low pH are characterised by lower microorganism activity.Earlier research demonstrated that the combination of arbuscular mycorrhizal fungi(AMF)and green algae resulted in carbon decomposition and formation of soil aggregates through pH reduction and increased C bacteria biomass,and consequently root density(Al-Maliki and Breesam,2020).A clear correlation between upper soil pH values and ectomycorrhizal fungi investigated in five different forest tree species was not observed by Shirkonyer et al.(2013).
5.Conclusions
The conducted study confirmed the significance of climatic conditions for the properties of montane forest soils and the biomass and morphology of beech and fir roots.With decreasing temperature shortening the vegetative period at higher positions along the elevation gradient,the organic residue decomposition process is limited,soil pH is reduced and the conditions for the development of root system deteriorate.At the highest elevations,characterised by a harsher climate,lower root biomass and poorer root production were observed.Limitation of root growth occurred in both study species;however,in the case of beech,greater adaptive capacity to the changing environmental conditions was observed.It can be expected that,in the event of climate warming,beech will be predisposed for expanding its distribution range.
Authors’contributions
K.S.,E.B.and J.L.:conceived and designed the investigation;analysed and visualized the data;E.B.and J.L.:concepts research methodology;K.S.,E.B.and J.L.:preparation of manuscript.
Availability of data and material
The data that support the findings of this study are available from the corresponding author on reasonable request.
Declaration of competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
This work was supported by the Ministry of Science and Higher Education of the Republic of Poland(AD11)and were additionally supported by project No.2021/41/N/NZ9/00264,PRELUDIUM-20,National Science Centre,Poland.

- Forest Ecosystems的其它文章
- Multiple forest structural elements are needed to promote beetle biomass,diversity and abundance
- Diversity of click beetles in managed nonnative coniferous and native beech stands:Consequences of changes in the structural and species composition of tree stands in Central Europe
- Environmental and canopy conditions regulate the forest floor evapotranspiration of larch plantations
- Allometry-based estimation of forest aboveground biomass combining LiDAR canopy height attributes and optical spectral indexes
- Examining approaches for modeling individual tree growth response to thinning in Norway spruce
- Influence of soil and elevation on roadside cryptogam diversity in the tropical Andes