
基于功能性状的外来植物入侵预测模型框架构建
2022-12-30 06:58王世雄何跃军王文颖
广西植物 2022年11期
王世雄, 何跃军, 王文颖
(1. 贵州大学 林学院, 森林生态研究中心, 贵阳 550025; 2. 青海师范大学 生命科学学院, 高原科学与可持续发展研究院, 西宁 810008 )
全球生态系统正经历着前所未有的生物多样性变化,外来物种的入侵是重要的驱动力之一(Hulme, 2012; Simberloff et al., 2013; 杨继和李博,2017; Munro et al., 2019)。预测外来植物的潜在入侵性已成为生物多样性保护的重要研究内容(Mack et al., 2000; 郑珊珊等,2018)。生物拮抗(biotic resistance)是决定外来植物能否成功入侵的重要机制,即乡土物种对外来物种表现的竞争效应(Conti et al., 2018),是广泛存在的对潜在入侵者的一个重要自然屏障。外来物种必须突破生物拮抗,才有可能成功入侵(Nereu et al., 2018)。在所有其他条件相同的情况下,较大的生物拮抗将意味着较低的入侵成功率。达尔文就曾注意到外来植物与乡土物种间的亲缘关系对外来植物成功入侵的重要作用,并提出了达尔文归化假说(Darwin’s naturalization hypothesis, DNH)和预适应假说(Darwin’s preadaptation hypothesis, DPH)(Darwin, 1859)。随着谱系生态学的发展,这两个假说逐渐成为描述外来植物和乡土物种间的亲缘关系与外来植物能否成功入侵的重要理论依据(Strauss et al., 2006; 王坤等,2009; Castro et al., 2014),这为外来植物的入侵预测提供了研究思路。
1 达尔文归化难题
达尔文归化假说(DNH)认为,在含有同属乡土物种较多的地方,外来植物不太容易建立自我维持的可持续种群(Daehler, 2001)。这是因为较近的亲缘关系将表现为较大的物种相似性(Burns & Strauss, 2011),意味着存在剧烈的种间竞争、共同的天敌以及相似的病原菌。种间竞争与物种相似性紧密相关,物种间的相似性越高,生态位越相似,竞争排斥越强烈,物种共存的难度越大。限制相似性假说(limiting similarity hypothesis)认为,共存物种间存在限制相似性以避免激烈的种间竞争(Macarthur & Levins, 1967)。换句话说,共存于同一个生态系统的植物性状应该趋异(trait divergence),拥有与乡土群落不同性状的外来植物将具有较大的入侵潜力,即生态位分化假说(niche differentiation hypothesis)(Lambdon et al., 2008)。同时,达尔文意识到,外来植物在含有较多同属乡土物种的生境中成功入侵也是可能的。特定生境中的关键环境因素决定着哪些物种可以生长、繁殖、扩散,即什么样的生境适合什么样的外来植物入侵,这被称之为生境过滤假说(habitat filtering hypothesis)(Kraft et al., 2015)。相近的亲缘关系意味着相似的生态需求,使外来植物对当地环境反而因有更好的适应性而有利于成功入侵(Procheet al., 2008),即预适应假说(DPH)。例如,我国和北美、热带美洲的经纬度、温度等气候因子都比较类似,因此我国北方已分布的入侵植物起源于北美的物种也占比较大,而长江以南地区已分布的入侵植物起源于热带美洲的物种占比较大。近20年来,学者们对这两个对立的假说进行了很多实证研究,但得到了很不一致的结果(Jones et al., 2013; Li et al., 2014; Qian & Sandel, 2017)。因此,达尔文归化假说和预适应假说被合称为达尔文归化难题(Darwin’s naturalization conundrum)(Diez et al., 2008; Park et al., 2020)。由于达尔文的观点是基于物种的系统发育相关性与决定物种共存的功能性状相似性有关这样一个假设,因此亲缘关系可以作为性状相似性的代理,潜在地提供尚未量化的性状信息。然而,也有研究发现,亲缘关系密切相关的物种在影响物种入侵性的关键性状上存在显著差异(Funk & Vitousek, 2007),从功能性状的角度理解生物抗性可能是一个更有前景的方法(Yannelli et al., 2017)。
2 功能性状的重要性
功能性状决定了物种个体的存活、生长和繁殖(Adler et al., 2014),以及物种间相互作用的强度和标志(Kunstler et al., 2016)。越来越多的证据表明,解决物种与物种以及物种与环境间相互作用的方法是利用物种的功能性状,而不是依赖于它们的分类学特性(Violle et al., 2007; Levine, 2016; Pérez-Ramos et al., 2019)。Yannelli等(2017)研究表明,亲缘关系较近物种的优势性状在抵御外来物种的入侵中发挥了最重要的作用。然而,功能性状如何影响物种共存以及决定外来植物入侵,目前知之甚少。建立功能性状与外来植物间的关系可预测外来植物的入侵,为外来植物的入侵防治提供理论依据。
物种的功能性状一方面保持了进化上的保守性,另一方面表现为可塑性。功能性状的种间分化(interspecific differentiation)和种内变异(intraspecific variation)是物种实现对群落环境适应以及对种间竞争响应的重要生态策略。
2.1 功能性状的种间分化
外来植物对乡土群落的入侵过程就是乡土群落被外来植物入侵干扰后的群落构建过程,外来植物的入侵必然伴随着有限资源的争夺。因此,物种间的生态位分化是外来植物实现成功入侵的一般前提。植物的种间性状分化在一定程度上能够反映其生态位分化,导致植物在群落内的适合度差异,进而促进外来植物的成功入侵。物种间的生态分化可体现为与其资源获取和利用策略相关的功能性状分化(Wright et al., 2004)。同时,与资源策略相关的功能性状之间应存在协调性,以优化植物的资源获取和利用效率(Reich et al., 2003; Aubin et al., 2016)。
2.2 功能性状的种内变异
性状的种内变异是群落构建的重要贡献者(唐青青等,2016; He et al., 2018)。同一物种的不同个体间存在明显的功能性状变异,即性状可塑性(trait plasticity),这种变异可以有效避免或者缓冲种内竞争,是物种生存策略的权衡,具有十分重要的进化和生态学意义(Mackay et al., 2009)。此外,性状可塑性使植物能够应对环境异质性,并可能在更广泛的环境下生存(Palma et al., 2021)。性状可塑性也可能使外来植物和乡土物种的生态位重叠最小化,从而提高外来植物成功入侵的概率(Bennett et al., 2016)。将性状的种内变异引入外来植物入侵中,不仅可以极大提高物种入侵机制检测的灵敏度(Jung et al., 2010; Paine et al., 2011),而且可以有效揭示物种入侵机制(Albert et al., 2011; Clark et al., 2011)。有研究表明,在资源有限的环境中,性状可塑性能够提高外来植物对资源的获取能力及利用效率,加速外来植物的生长和繁殖,从而增强其种群竞争能力(Leicht-Young et al., 2007; Funk, 2008)。与乡土植物相比,入侵植物常常具有较大的性状可塑性(Daehler, 2003; Mozdzer et al., 2012; 潘玉梅等,2017),约有50%入侵植物的入侵能力与其性状可塑性相关(Ren & Zhang, 2009; 刘建等,2010;郑欣颖和薛立,2018)。然而,群落生态学家似乎更重视性状的种间分化,却忽视了种内变异的重要性,而性状的种间分化与种内变异是外来植物适应异质生境和实现成功入侵的两种不同生态策略(Helsen et al., 2020)。因此,在入侵生态学中,真正值得关注的不仅仅是性状的种间分化或者种内变异的绝对大小,而是二者的相对重要性及其权衡关系。
3 外来植物入侵预测的理论基础
达尔文归化难题通过功能性状的引入就可以描述为如下两个不同的假说。其一,亲缘关系相近的一些物种集合,它们占据相同或相似生态位、资源利用方式相同或相近,物种间存在着强烈竞争,在含有相似功能性状的群落中,被引入的外来植物建立自我维持的野生种群的可能性将减少(Daehler, 2001),即达尔文归化假说。其二,外来植物和乡土物种具有相似的功能性状特征,对资源需求基本一致(Hooper & Vitousek, 1997; Naeem et al., 2000; Symstad, 2000; Dukes, 2001; Young et al., 2009),使其对当地环境有着更好的适应性而有利于成功归化或者入侵,即预适应假说(Daehler, 2001; Diez et al., 2008)。达尔文归化假说强调的是物种间的相互作用,而预适应假说强调的是物种对生境的共同需求(Skóra et al., 2015),外来植物能否成功入侵依赖于生境需求与种间互作的权衡,而这种权衡关系又依赖于外来植物与乡土群落间的相似性。
近年来,基于物种间关系的入侵研究取得了一些重要的成果,但这些研究主要集中在外来植物与乡土物种二者间的亲缘关系方面(Carmona et al., 2019; van Klink et al., 2019; Dong et al., 2020),并且基于控制实验的研究结果是否与自然条件下的结论一致仍是一个悬而未决的问题。自然界中的物种关系并不是简单的两两间的物种关系,而是外来植物与乡土群落所有物种间的一对多的关系。同时,一对多的种间关系并不等于两两间物种关系的平均。不同的乡土群落意味着不同的物种多样性、不同的功能群组成、不同的物种多度格局等,这些不同决定了外来植物的入侵预测必须从群落水平上整体考虑。因此,研究外来植物和乡土物种亲缘相似性与入侵结果间的关系,应该从外来植物与乡土物种二者间的亲缘关系转变为外来植物与乡土群落间的相似性关系(Skóra et al., 2015)。通过构建外来植物与乡土群落间的相似性关系来进行外来植物入侵预测,该法从整体上考虑外来植物与乡土群落间的关系,排除了特定种间关系(如乡土群落中的优势物种)对入侵的主导性,这使得预测结果更加准确和全面。
为了评价外来植物与乡土群落间的相似性,应该引入基于功能性状的功能相似性指数(functional similarity index, FSI)。根据外来植物与乡土群落间的相似性,理论上有两种理想的关系模式(Skóra et al., 2015),即指数模式和对数模式(图1)。指数模式表示的外来植物与乡土群落间的相似性较大,即外来植物与绝大多数乡土物种功能性状相似,在该模式下,如果外来植物成功入侵,其可能是因为达尔文预适应假说(DPH),而入侵失败则可能是由于达尔文归化假说(DNH);对数模式表示的外来植物与乡土群落间的相似性较小,即外来植物与绝大多数乡土物种功能性状不相似,在该模式下,如果外来植物成功入侵可能是因为达尔文归化假说(DNH),而入侵失败则可能是由于预适应假说(DPH)。
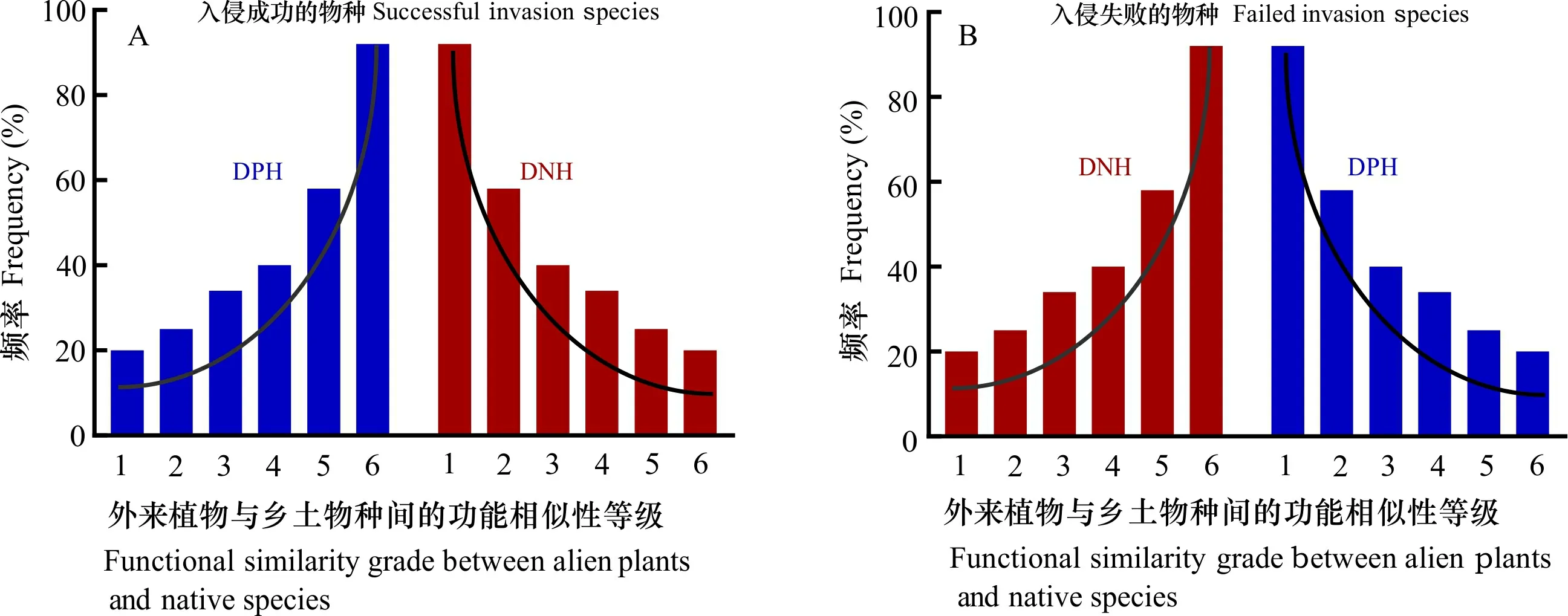
DNH表示达尔文归化假说; DPH表示达尔文预适应假说。下同。引自参考文献(Skóra et al., 2015),有修改。DNH indicates Darwin’s naturalization hypothesis; DPH indicates Darwin’s preadaptation hypothesis. The same below. Figure modified from the reference (Skóra et al., 2015).图 1 基于功能性状的外来植物与乡土群落间相似性及其入侵结果Fig. 1 Chart of similarity between alien plants and native communities based on functional traits and its invasive results
建立基于功能性状的入侵植物与乡土群落间的相似性,不仅可以有效预测外来植物能否成功入侵,而且可以通过群落功能性状的变化揭示外来植物入侵机制。对数模式下的成功入侵,种间互作是主要作用,反映在物种性状上可能表现为种间分化的主导性,生态位空缺或者生态位分化是外来植物成功入侵的可能机制(Lambdon et al., 2008; Dong et al., 2020);指数模式下的成功入侵,反映了物种间的共同生境需求,在该模式下要实现成功入侵,性状的种内变异应该是主要贡献者。在性状趋同的前提下,性状的种内变异能够提高外来植物对资源的获取能力及利用效率,加速外来植物的生长和繁殖,从而增强其种群竞争能力,内禀优势应该是外来植物成功入侵的可能机制(Sax & Brown, 2000; Leicht-Young et al., 2007; Funk, 2008)。
4 外来植物入侵预测的流程
具体来说,建立基于功能性状的外来植物入侵预测,大体上需要以下3个基本过程(图2):一是功能性状的测定与筛选;二是基于功能性状计算外来植物与乡土群落间的相似性;三是基于功能性状种间分化和种内变异的分解探究外来植物的可能入侵机制。
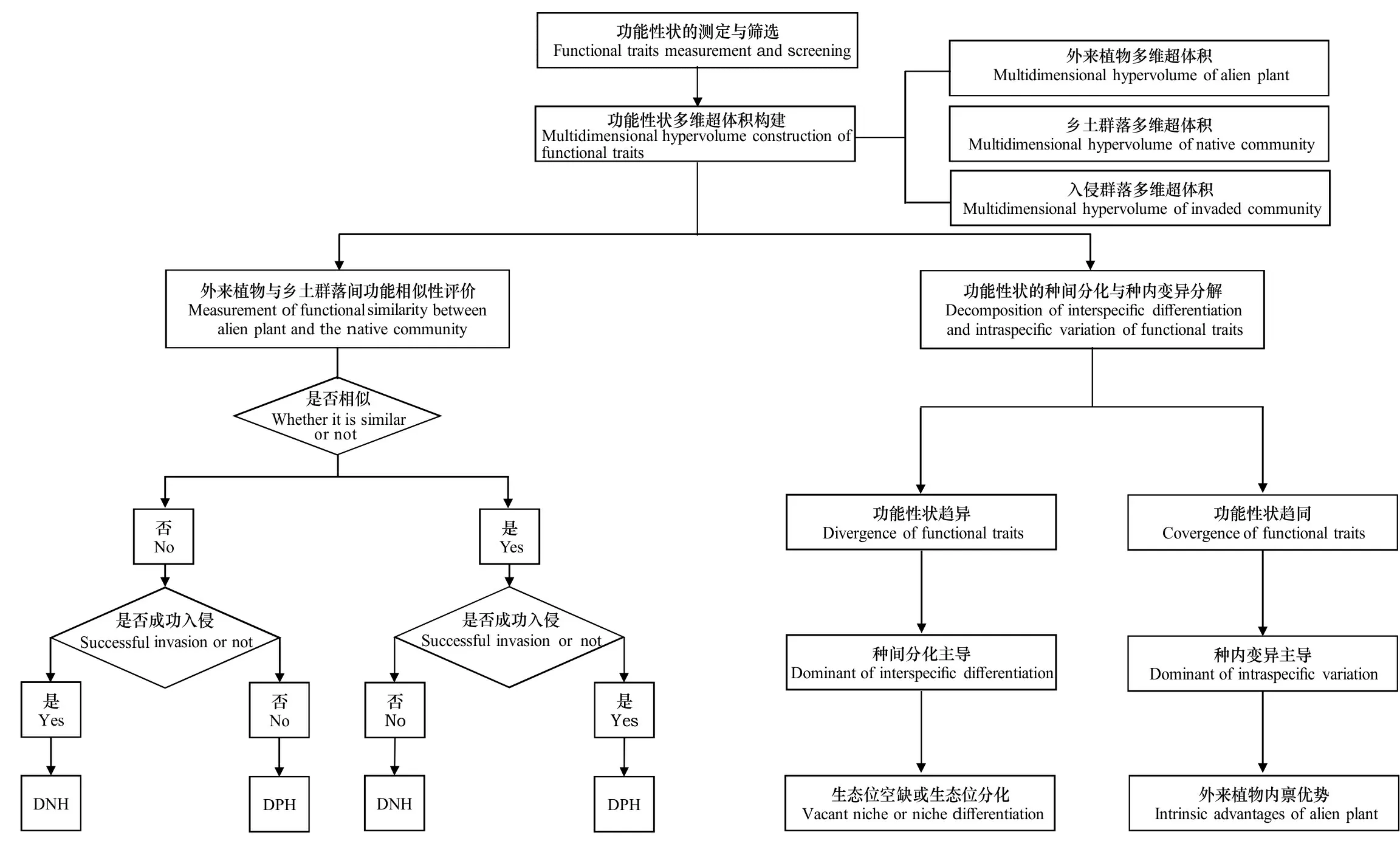
图 2 基于功能性状的外来植物入侵预测流程示意图Fig. 2 Flow chart of invasion prediction of alien plants based on functional traits
4.1 功能性状的测定与筛选
有研究表明,并不是所有的性状都适合用来预测特定的生态功能,性状的选择必须基于其作用于特定生态功能下的潜在机制。“合理的性状选择—考虑性状种间分化和种内变异—功能多样性指数的选择—功能性状与生态功能的尺度匹配”是提高物种功能性状对生态功能预测的有效手段(Liu et al., 2021)。某些性状的大量测量和广泛的研究并不意味着它们在外来植物入侵中发挥重要作用,可能仅仅是因为它们更易测量。外来植物具有不同的入侵方式,并依赖于不同的性状组合。例如, 一些性状 (如种子质量)对入侵的不同阶段表现了促进、阻碍或没有显著影响的复杂作用(Palma et al., 2021)。因此,功能性状的选择和筛选成为了外来植物预测的重要基础。今后,物候特征、植物发育阶段、共存植物生长的异步性以及一些难以测量的“硬”性状都应引起生态学家的足够重视(Liu et al., 2021)。
4.2 外来植物与乡土群落间的相似性分析
目前,外来植物与乡土群落间的功能相似性构建主要有两种思路:一种是基于外来植物与乡土群落中每一物种的相似性而形成的相似性等级谱;另一种是基于超体积生态位的n维性状空间及其超体积质心之间的距离大小。由于基于性状的生态位差异是多维的,外来植物和乡土群落间的相似性或者差异性应该同时跨越多个性状进行探索(Geange et al., 2011; Blonder et al., 2014; Bittebiere et al., 2019)。因此,本研究推荐使用第二种方法进行外来植物与乡土群落间的相似性评价。
Hutchinson的n维超体积概念将生态位解释为几何形状,为跨生态学和进化学的不同领域的研究提供了基础(Hutchinson, 1957)。这些形状可以被解释为生态位、生态或进化策略空间或群落结构的代表。该方法通过评估表征一组物种所占据的功能性状的超体积来量化生态位空间,同时,性状超体积算法可以度量共享相同功能空间的超体积比例以及每个物种的特异性体积分数。因此,n维性状超体积能够预测物种间沿着环境梯度生态位转移的群落内产生的功能冗余(重叠),并且能推断相关生态过程,如竞争排斥和生态位分化(de la Riva et al., 2018)。目前,性状超体积概念的应用范围不断扩大,有越来越多的统计方法可用来进行运算。例如,基于多元核密度估计的方法(Mouillot et al., 2005; Mason et al., 2008; Blonder et al., 2014)构建性状超体积结构来评估外来植物入侵过程中的多维生态位重叠,并表现出了很好的效果和优势。
功能性状超体积构建后,可以通过计算多维性状空间中超体积质心之间的欧氏距离、每个超体积的唯一体积分数以及超体积结构之间的Jaccard相似性来评估每个数据集的超体积结构之间的功能相似性(functional similarity)和生态位重叠情况。其中,Jaccard相似性定义为两个超体积结构交集的体积除以两个超体积结构并集的体积。
4.3 功能性状的种间分化与种内变异分解
为了评估性状的种间分化和种内变异对功能性状空间的相对贡献可以通过性状变异分解的方式来实现(Lepš et al., 2011),目前已有诸如“cati”等多个R语言的软件包可以轻松实现(Taudiere & Violle, 2016)。其基本过程如下:首先,计算包含样方间种内变异和物种周转的性状总变异T1。T1既包括不同样方(或环境)中的物种周转,也包括不同样方(或环境)中同一物种的性状值的种内变异。然后,固定所有样方中同一物种的性状值,计算一次性状变异T2。因为去除了同一物种的种内变异,所以T2只包括由样方间物种周转所引起的性状变异。最后,通过计算T1和T2之差可以估计种内性状变异T3的相对大小。计算公式如下:
T3=T1-T2。
式中:pi为第i个物种的比例;xi为固定了所有样方中第i个物种的性状平均值;xi_habitat为第i个物种在不同生境中的特定性状值;S为群落中的物种总数。
5 研究展望
功能性状超体积的引入为建立外来植物与乡土群落间的相似性奠定了重要基础,同时,性状的种间分化和种内变异的分解也为揭示外来植物入侵机制提供可能途径。尽管这些均是进行外来植物入侵预测的重要基础,但是,这些美好的前景也面临着一些挑战。
5.1 入侵的生境依赖性
尽管功能性状在物种入侵中扮演了十分重要的作用,但是外来植物要能成功入侵,首先要突破区域环境的限制。生境条件不仅影响外来植物的入侵过程,而且影响乡土物种的组成(Richardson & Pyšek, 2012; Pearson et al., 2018)。环境过滤将导致性状和功能相似的物种被筛选进入相近的生态位,使群落内各物种的性状趋同(Bazzaz, 1991; Lavorel & Garnier, 2002)。相反,竞争排斥会使得生态位相似的物种无法共存于同一环境,而群落内物种的亲缘关系则较远,表现为各物种的性状趋异发散(Webb et al., 2002; Ackerly & Cornwell, 2007)。环境过滤和生物拮抗共同作用决定了入侵植物性状特征的分布格局。一般来说,环境中主要资源(如光照、水分等)梯度决定了物种功能性状的分布范围(Fargione & Tilman, 2005; Kraft et al., 2015),而生物拮抗决定了物种性状变异的方向和程度。外来植物的入侵过程就是环境过滤、种间竞争和种内性状变异耦合作用的结果。例如,对北非干旱带的研究表明,在资源极端限制的环境中,外来植物和乡土物种在性状上相似,这支持了达尔文预适应假说(DPH)。然而,在资源相对丰富的环境中,外来植物和乡土植物在性状上不相似,外来植物表现出更具入侵性的性状(更高的比叶面积、地上生物量和高度),这为达尔文归化假说(DNH)提供了支持(El-Barougy et al., 2020)。在对中欧温带地区的6种不同生境类型中的入侵研究表明,外来物种在植物群落中成功建立的主要决定因素是环境过滤。然而,已建立的外来植物如果想要成为入侵物种,就必须有足够的差异性来占据新的生态位空间,即性状空间的边缘(Divíšek et al., 2018)。因此,进一步了解环境条件如何决定植物群落的功能结构和生态位超体积以及检验不同环境梯度上的外来植物入侵过程是十分必要的。综上认为,基于功能性状的预测模型应该充分考虑生境条件的重要性。
5.2 空间尺度的重要性
越来越多的研究表明,达尔文的两个对立的假说并不一定是相互排斥的(Carboni et al., 2013; Park et al., 2020)。相反,在不同环境或空间尺度下,每种假说都可以正确地预测生物入侵的结果。例如,Meta分析发现,引进的外来物种在局域尺度上与乡土物种的亲缘关系更密切,但在更大的区域尺度上与乡土物种的亲缘关系则不那么密切(Li et al., 2016)。在小空间尺度(如地块或生境尺度)上,近缘物种的相互排斥格局可能更加严重(Swenson et al., 2007)。在更大的空间尺度上,由于广泛的共同环境偏好和较少的种间竞争,近缘物种更有可能共存(Procheet al., 2008)。美国植物区系的研究表明,随着空间尺度的增大,环境过滤特性超过竞争特征的概率增大,达尔文的两个对立假说在不同的空间尺度上得到了合理解释(Park et al., 2020)。
5.3 乡土群落的可入侵性
外来植物能否最后成功入侵,不仅取决于其与乡土群落的相似性或差异性,同时还依赖于乡土群落的可入侵性。从功能性状的角度看,物种丰富的群落表现出不同的功能性状和获取资源的策略,这往往会导致较高的功能多样性。相应地,物种较少的群落可能意味着生态位的空余,一些建立早、生长快的外来植物可能通过快速抢占资源产生优先效应,从而抑制生长缓慢的乡土物种在群落中的聚集。同时,物种多样性对生物拮抗的影响可以进一步划分为选择效应和互补效应(Loreau, 1998; Loreau & Hector, 2001)。选择效应是指具有特定性状的物种优势将决定多样性效应的情况,而互补效应描述的是物种之间的资源分配或物种间的积极相互作用对多样性效应贡献最大的情况(Loreau & Hector, 2001)。有研究表明,空生态位抢占(优先效应)和生态位分化(多样性效应)均是芦苇(Phragmitesaustralis)成功入侵的潜在机制(Byun et al., 2013)。因此,结合外来植物的功能性状以及乡土群落的可入侵性可能在外来植物入侵预测方面提供新的见解。
本研究解析了达尔文归化难题的内涵,从物种功能性状的保守性、种间分化及其种内变异进行了分析,提出功能性状的种间分化与种内变异是外来植物实现成功入侵的两种不同生态策略。在此基础上,本研究通过物种功能性状的多维超体积构建了外来植物和乡土群落间的相似性,提出了基于这种相似性的外来植物入侵预测的研究框架和基本流程。该模型框架对理解外来植物的入侵机制提供了理论依据,对外来植物的入侵预测提供了实践指导。当然,要实现外来植物成功入侵的准确预测,不仅依赖于功能性状的选择和测定,还要重点考虑入侵的生境依赖性、空间尺度的重要性以及乡土群落的可入侵性等,未来的研究重点是通过控制实验对该机制模型进行验证和进一步完善。
猜你喜欢
数学物理学报(2022年5期)2022-10-09
当代水产(2022年8期)2022-09-20
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
中国农业科学(2022年10期)2022-06-28
昆明医科大学学报(2022年2期)2022-03-29
广东蚕业(2021年12期)2022-01-18
中南林业科技大学学报(2021年11期)2021-12-17
食品安全导刊(2021年20期)2021-08-30
河北画报(2020年8期)2020-10-27
热带农业科学(2019年8期)2019-09-19
