
甘蔗ScNRAMP基因家族的鉴定与生物信息学分析
2022-12-30 07:20:06江姚兰周定港
广西植物 2022年11期
刘 营, 尹 泽, 江姚兰, 周定港,3*
(1. 湖南科技大学 生命科学学院, 湖南 湘潭 411201; 2. 经济作物遗传改良与综合利用湖南省重点实验室, 湖南 湘潭 411201; 3. 福建农林大学农业农村部福建甘蔗生物学与遗传育种重点实验室, 福州 350002 )
甘蔗(Saccharumspp.)是重要的一年生或多年生糖料作物,属于单子叶植物纲(Monocotyledoneae)禾本科(Poaceae)甘蔗属(SaccharumL.)(Irvine, 1999)。甘蔗是一种高光合效率的C4植物,具有生物量大、CO2补偿点低、耐干旱、适应性广和产量高等特点(方静平等, 2014)。甘蔗是异源多倍体,其倍性水平从5×到16×不等,其基因组大小约为10 Gb(陈如凯, 2011)。现代栽培甘蔗是世界上最重要的食糖来源,占全世界食糖总量的80%(Liu et al., 2020)。
NRAMP转运体在离子稳态的维持,尤其是在二价金属离子的转运上发挥着重要作用,并且该蛋白家族在进化过程中高度保守(Nevo & Nelson, 2006)。哺乳动物NRAMP基因的氨基酸序列与酵母、水稻和果蝇NRAMP基因的氨基酸序列相似性分别为46%、58%和73%(Belouchi et al., 1995)。在植物吸收重金属的过程中,NRAMP蛋白等膜转运蛋白发挥着重要作用(Cellier et al., 1995)。Mäser等(2001)对拟南芥的6个NRAMP基因进行克隆和系统进化分析发现,6个家族成员被分为2个亚家族,其中AtNRAMP1和AtNRAMP6位于第一亚家族,AtNRAMP2至AtNRAMP5属于第二亚家族。AtNRAMP1蛋白是模式生物拟南芥中的主要高亲和性锰转运蛋白,定位于质膜,其功能是将重金属转运至细胞质(Cailliatte et al., 2010)。Pottier等(2015)研究发现诱导表达AtNRAMP4的酵母株突变会导致其镉和锌的吸收下降,而对于铁的吸收量没有改变,此研究提示人为诱导NRAMP转运蛋白的突变可用于降低动植物对Cd的吸收和转运。不同植物的NRAMP家族成员具有组织表达特异性,如AtNRAMP1在根中高表达(Castaings et al., 2016),而AtNRAMP2则主要在根表皮和根尖区域表达(Gao et al., 2018);OsNRAMP1在营养生长阶段的根部以及在生殖生长期时的叶片和茎中均为高表达(Takahashi et al., 2011)。
甘蔗是重要的经济作物,与其他植物一样也面临着重金属污染问题。重金属对于植物的生长起着十分重要的作用,一些二价重金属离子如Cu2 +、Zn2 +和Mn2 +等是植物生长必需的微量元素,而有些重金属如Cd2+和Pb2+等的过度吸收则会造成植物的重金属毒害甚至影响产量(Rosa-Santos et al., 2020)。植物NRAMP蛋白的功能研究在拟南芥(Thomine et al., 2000)、水稻(Luo et al., 2018)、土豆(Tian et al., 2021)、大豆(Qin et al., 2017)和油菜(Meng et al., 2017)等植物中虽有广泛报道,但关于甘蔗NRAMP蛋白的研究目前还没有报道。本研究基于甘蔗割手密种的全基因组序列,利用生物信息学的方法筛选甘蔗ScNRAMP基因家族成员,并对其进行进化树构建、染色体定位、基因结构分析和蛋白特性分析等,以期为后续该基因家族的研究提供参考。
1 材料与方法
1.1 材料
AtNRAMP基因家族的蛋白序列来源于拟南芥TAIR数据库,甘蔗全基因组和基因注释文件来源于http://www.life.illinois.edu/ming/downloads/Spontaneum_genome/ (Zhang et al., 2018)。
1.2 方法
1.2.1 甘蔗ScNRAMP基因家族的鉴定 以拟南芥NRAMP蛋白序列作为种子序列,在TBtools(版本号为v1.0971,Chen et al., 2020)进行第一次Blastp,在NCBI进行第二次Blastp;结合SMART(http://smart.embl-heidelbreg.de/)对候选基因的氨基酸序列结构域进行鉴定,手动剔除不含NRAMP保守结构域的序列,保留下的即为甘蔗ScNRAMP基因家族成员。
1.2.2 编码蛋白理化特性 利用ExPASy在线软件(https://us.expasy.org/tools/protparam.html/)对ScNRAMP基因家族编码蛋白的等电点、分子量、序列长度、总平均疏水性、不稳定系数和脂溶指数等理化性质进行分析预测;利用SignalP v4.1(http://www.cbs.dtu.dk/services/SignalP-4.1/)和CELLO v2.5(http://cello.life.nctu.edu.tw/)分别对29条蛋白序列进行信号肽预测和亚细胞定位分析。
1.2.3 蛋白二级结构和跨膜结构的分析 利用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html)分析NRAMP蛋白的二级结构,利用TMHMM Server v2.0(http://www.cbs.dtu.dk/services/TMHMM/)分析该蛋白家族的跨膜结构。
1.2.4 甘蔗ScNRAMP基因保守基序、结构域及基因结构的分析 首先,利用MEME(https://meme-suite.org/meme/)和NCBI-CDD(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)检测甘蔗ScNRAMP家族基因中所存在相似度较高的基序(motif)和保守结构域(domain);然后,利用基因组注释文件,获得家族成员内含子及外显子分布情况;最后,利用TBtools软件对其进行可视化分析。
1.2.5 甘蔗ScNRAMP基因家族顺式作用元件的分析 提取ScNRAMP基因上游3 000 bp序列作为甘蔗NRAMP基因的启动子;利用PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)对启动子区域顺式作用元件进行分析。
1.2.6 甘蔗ScNRAMP基因的染色体定位 运用TBtools软件,结合甘蔗基因组注释文件和家族成员IDlist,将结果可视化。
1.2.7 甘蔗ScNRAMP家族进化树的构建 使用本地软件MEGAX完成多序列比对并采用邻接法(neighbor joining)构建系统进化树,其中校验参数(bootstrap)设置为1 000次,其余均设置为默认参数;利用在线Evolview软件(https://evolgenius.info//evolview-v2/#login )对进化树进行绘制编辑。
1.2.8 甘蔗ScNRAMP基因的表达分析 利用割手密的RNA-seq转录组表达数据进行甘蔗ScNRAMP基因的表达分析。割手密的转录组RNA-seq表达谱下载自割手密基因组数据库(Li et al., 2020)。筛选得到ScNRAMP基因家族成员在不同时期不同组织中的表达量(FPKM值),并用TBtools进行聚类,绘制热图。
2 结果与分析
2.1 甘蔗NRAMP蛋白的理化特性
从甘蔗割手密基因组中共鉴定到29个甘蔗ScNRAMP基因家族成员,其一级结构和理化性质如表1所示。甘蔗ScNRAMP基因家族成员的氨基酸总数在334~1 272个之间,氨基酸长度平均为563,分子量维持在55 000 Da左右;等电点跨度较大,在4.77~9.43之间;家族成员的不稳定系数大部分低于40%,只有7个成员不稳定系数在40%~48%之间;家族成员平均亲水系数在0.015~0.949之间;29个甘蔗ScNRAMP蛋白均无信号肽,亚细胞定位分析表明成员均定位在质膜上。

表 1 甘蔗NRAMP蛋白理化特性Table 1 Physicochemical properties of sugarcane NRAMP proteins
2.2 蛋白二级结构和跨膜结构分析
甘蔗NRAMP蛋白二级结构和跨膜结构分析如表2所示。ScNRAMP基因家族的二级结构主要由α-螺旋、无规则卷曲、延伸链和β-转角组成,其中α-螺旋比例最高,为36.71%~64.32%,β-转角所占比例最小,为1.62%~5.06%;蛋白家族的二级结构较为整齐,除在Sspon.03G0024310-2B和Sspon.03G0024310-1A中所占比例是无规则卷曲>α-螺旋>延伸链>β-转角外,其余家族成员均为α-螺旋>无规则卷曲>延伸链>β-转角;29个甘蔗NRAMP家族成员均有跨膜结构,其数量为6~12个不等。这表明甘蔗NRAMP蛋白为跨膜蛋白,可能与其转运重金属离子的功能相适应。
2.3 甘蔗ScNRAMP基因保守基序、结构域及基因结构分析
对甘蔗ScNRAMP基因的保守基序、结构域及基因结构分析如图1所示。通过对保守基序分析发现motif 1基序高度保守,在割手密29个NARAMP家族成员中均存在; 个别家族成员存在2个motif 9,如Sspon.02G002160-2C、Sspon.02G0057810-1D、Sspon.02G002160-1A和Sspon.02G0021270-1A等;大部分家族成员中有9个保守基序,而成员Sspon.02G0029680-1A、Sspon.05G0038800-1D和Sspon.02G0057180-1D中最少,仅有6个保守基序(图1,表3)。保守基序的差异可能暗示该基因家族成员之间在功能上存在差异。

表 3 甘蔗ScNRAMP基因保守基序logoTable 3 Conservative motif logo of sugarcane ScNRAMP genes

A. ScNRAMP基因的保守基序(motif)分布,10个motif用不同颜色的方框表示; B. ScNRAMP基因保守结构域(domain)分布;C. ScNRAMP基因内含子和外显子分布。绿色方框表示外显子;黑色线条表示内含子;黄色框表示基因的上下游区域。A. Distributions of conserved motifs in ScNRAMP genes, ten putative motifs are indicated in different colored boxes; B. Distributions of domains in ScNRAMP genes; C. Exon and intron organizations of ScNRAMP genes. Green boxes represent exons; Black lines represent introns; The upstream and downstream regions of ScNRAMP genes are indicated in yellow boxes.图 1 甘蔗ScNRAMP基因保守基序、结构域及基因结构Fig. 1 Conserved motif, domain and gene structure analysis of sugarcane ScNRAMP genes
通过对基因结构分析发现ScNRAMP基因家族全都含有内含子,并且内含子数目差异较大(为2~11个不等);部分基因成员在5′末端和3′末端不含非翻译区(untranslated region,UTR)。
通过对保守结构域分析发现,除Sspon.02G0057810-1D、Sspon.05G0038800-1D、Sspon.01G0025840-2B、Sspon.01G0061080-1D和Sspon.01G0025840-1A这5位成员的保守结构域为Nramp superfamily外,其余24位家族成员的结构域均为Nramp。
2.4 甘蔗ScNRAMP基因家族顺式作用元件分析
ScNRAMP基因起始密码子上游3 000 bp启动子区域顺式作用元件分析结果如图2所示。在ScNRAMP基因启动子上含有与生长发育有关的作用元件,即CAT-box(分生组织调控元件)、RY-element(种子特异调控元件);与激素响应相关的顺式作用元件,即ABRE(脱落酸响应元件)、TGACG-motif(茉莉酸响应元件)、TCA-element(水杨酸响应元件)、TGA-element(生长素响应元件)和TATC-box/P-box(赤霉素响应元件);与非生物胁迫相关的作用元件,即TC-rich(防御和应激响应元件)、MBS(干旱响应元件)、 LTR (低温响应元件)、GT1-motif/G-box、MRE和I-box(光响应元件)和ARE(厌氧反应元件);与蛋白相关元件,即MBSI和MYBHv1(MYB转录因子结合位点)及O2-site(玉米醇溶蛋白代谢调节)。进一步分析发现,G-box、MRE、TGACG-motif、ABRE等元件存在于所有成员。以上结果表明,ScNRAMP基因家族很有可能通过TC-rich motif 和MRE motif等元件参与甘蔗的激素代谢,进而影响其对重金属胁迫的响应。
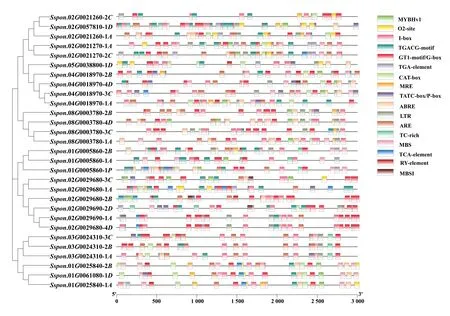
图 2 甘蔗ScNRAMP基因启动子区域顺式作用元件预测结果Fig. 2 Prediction result of cis-acting elements in the promoters of sugarcane ScNRAMP genes
2.5 甘蔗ScNRAMP基因的染色体定位
染色体定位结果如图3所示。甘蔗ScNRAMP基因的29个家族成员不均地分布在19条染色体上;染色体Chr2A上有4个基因,染色体Chr1B、染色体Chr2C和染色体Chr2D上各有3个基因,染色体Chr1A上有2个基因,其余14条染色体上各含有1个基因。

图 3 甘蔗ScNRAMP基因在染色体上的位置Fig. 3 Positions of sugarcane ScNRAMP genes on chromosomes
2.6 甘蔗ScNRAMP家族的系统发育树
为了解甘蔗割手密和其他禾本科植物NRAMP基因家族的进化关系,构建了水稻、拟南芥、割手密、玉米和高粱5个物种NRAMP家族成员的进化树(图4)。由图4可知,进化树有3个分支,分为3个亚家族。亚家族Ⅰ中,甘蔗ScNRAMP有14位成员(Sspon.05G0038800-1D、Sspon.04G0018970-3C、Sspon.04G0018970-1A、Sspon.04G0018970-2B、Sspon.04G0018970-4D、Sspon.02G0021260-1A、Sspon.02G0021260-2C、Sspon.02G0057810-1D、Sspon.02G0021270-1A、Sspon.02G0021270-2C、Sspon.08G0003780-2B、Sspon.08G0003780-1A、Sspon.08G0003780-3C和Sspon.08G0003780-4D),与高粱14位成员、玉米15位成员、拟南芥2位成员和水稻4位成员聚为一支。亚家族Ⅱ中,甘蔗ScNRAMP有9位成员(Sspon.01G0005860-2B、Sspon.01G0005860-1A、Sspon.02G0029680-2B、Sspon.02G0029680-1A、Sspon.02G0029680-3C、Sspon.02G0029680-4D、Sspon.02G0029690-1A、Sspon.02G0029690-2D和Sspon.01G0005860-1P),与高粱3位成员、玉米5位成员、拟南芥4位成员和水稻2位成员聚为一支。亚家族Ⅲ中,甘蔗ScNRAMP有6位成员(Sspon.03G0024310-3C、Sspon.03G0024310-2B、Sspon.03G0024310-1A、Sspon.01G0025840-2B、Sspon.01G0061080-1D和Sspon.01G0025840-1A),与高粱5位成员和玉米7位成员聚为一支,并且在此分枝上没有拟南芥和水稻的家族成员。水稻、拟南芥、高粱、玉米和甘蔗NRAMP家族成员之间均有同源基因,相比之下玉米与甘蔗的NRAMP基因家族成员之间亲缘关系更近。

黑色星号表示甘蔗ScNRAMP; 绿色方框表示玉米ZmNRAMP; 黄色圆圈表示水稻OsNRAMP; 红色三角形表示拟南芥AtNRAMP; 粉色对号表示高粱SbNRAMP。Black asterisks represent sugarcane ScNRAMP; Green boxes represent corn ZmNRAMP; Yellow circles represent rice OsNRAMP; Red triangles represent Arabidopsis AtNRAMP; Pink ticks represent sorghum SbNRAMP.图 4 甘蔗ScNRAMP的系统发育树Fig. 4 Phylogenetic tree of sugarcane ScNRAMP
2.7 甘蔗ScNRAMP基因组织表达
为了解甘蔗ScNRAMP基因家族成员的功能与时空表达模式,本文以公开发表的割手密基因表达数据分析了ScNRAMP基因在不同组织和不同发育时期的表达情况(图5)。图5结果显示,甘蔗ScNRAMP基因的表达呈现组织特异性。其中,Sspon.04G0018970-2B、Sspon.04G0018970-4D、Sspon.04G0018970-1A、Sspon.04G0018970-3C等基因在茎成熟期(M-s-3、6、9)表达量较高,在前成熟期和发芽期表达量较低;Sspon.03G0024310-3C、Sspon.03G0024310-2B、Sspon.03G0024310-1A、Sspon.01G0025840-2B和Sspon.01G0061080-1D5个成员在发育期叶中的表达量高于前成熟期和成熟期。另外,29个家族成员中仅有1个成员Sspon.05G0038800-1D的表达量FPKM值为0,表明其在各发育时期和组织部位均未表达。

S. 实生苗期; P. 成熟前期; M. 成熟期; s. 茎; l. 叶; 3. 甘蔗茎第3节; 6. 甘蔗茎第6节; 9. 甘蔗茎第9节。S. Seedling stage; P. Early maturity stage; M. Mature stage; s. Steam; l. Leaf; 3. Section 3 of sugarcane stem; 6. Section 6 of sugarcane stem; 9. Section 9 of sugarcane stem.图 5 甘蔗ScNRAMP基因表达模式分析Fig. 5 Expression pattern analysis of sugarcane ScNRAMP genes
3 讨论与结论
本研究对甘蔗ScNRAMP基因家族成员进行理化特性分析发现,不同的NRAMP蛋白序列有较大的差异,氨基酸长度为334~1 272 aa,等电点跨度较大(4.77~9.43),表明其编码蛋白能适应不同的酸碱环境。本研究中,甘蔗NRAMP蛋白不稳定系数均小于40%,表明其稳定性较好;蛋白的平均亲水系数介于0.015~0.949之间,表明其是一类相对疏水的蛋白;29个家族成员分布在19条染色体上,染色体Chr2A上最多,有4个家族成员;割手密NRAMP蛋白所有成员均有6~12个跨膜结构域,均分布在质膜上,可能与其具有转运重金属离子的功能相适应。基因结构分析显示,所有家族成员均具有内含子。
甘蔗割手密基因组中共鉴定出29个ScNRAMP基因,与水稻(6个成员)和拟南芥(6个成员)相比,ScNRAMP基因家族成员的数量明显要多,推测可能与甘蔗是多倍体且基因组庞大有关,尤其是甘蔗作为多倍体植物,其在多倍化进程中的全基因组复制促进了甘蔗基因组含量的提高及其基因家族的扩张,这与相关研究茄科植物高度变异的基因家族在全基因组复制和基因的串联重复引起基因组的含量增加和基因家族扩张的观点一致(Mäser et al., 2001;Zhang et al., 2018;Tian et al., 2021;Wang et al., 2021)。拟南芥和水稻的基因组大小分别为125 Mb和466 Mb,而甘蔗割手密的基因组大小则达到了3.36 Gb。此外,拟南芥和水稻均为二倍体,甘蔗为八倍体,而测序的甘蔗品种割手密AP85-441则是甘蔗八倍体SES208单花粉培育得到的整倍体(四倍体),理论上现代甘蔗栽培种(多为异源八倍体且为非整倍体)具有的ScNRAMP家族成员应多于29个(陈如凯, 2011; Zhang et al., 2018)。
基因结构和基序组成的分析,可为基因家族的进化关系提供重要依据(Boudet et al., 2001)。对ScNRAMP基因结构分析发现的同一亚家族的大多数基因在外显子、基序或非翻译区具有类似的结构特征,这一现象与其他物种的NRAMP蛋白家族相似(Belouchi et al., 1997;Lanquar et al., 2005),如AtNRAMP3和AtNRAMP4的基因结构相似,并且对Fe2+都有极高运输能力(Lanquar et al., 2005);OsNramp1、OsNramp2和OsNramp3序列的相似性为64%~75%(Belouchi et al., 1997),推测甘蔗NRAMP蛋白同一亚家族的成员在功能上可能相似。通过对甘蔗割手密、水稻、高粱、玉米4种单子叶植物和拟南芥1种双子叶植物的系统进化分析,可将29个ScNRAMP家族成员分为3个亚家族。在亚家族Ⅲ中,进化树分支上只有高粱(5位成员)、玉米 (7位成员) 和割手密(6位成员)的家族成员,没有水稻和拟南芥的成员,推测甘蔗割手密NRAMP亚家族Ⅲ中的这6位成员可能存在着与水稻和拟南芥的NRAMP蛋白家族不一样的特殊功能,而这个亚家族的特殊功能在高粱和玉米中却可能存在。
植物启动子作为植物功能基因表达调控的最重要顺式元件之一,启动子分析有助于阐明基因表达的调控和响应机制。通过对启动子区域的顺式作用元件分析,可以得到植物关于响应特定生物胁迫或非生物胁迫的重要元件,并由此推测甘蔗割手密种可能通过光响应因子、 激素响应元件和抗逆响应因子(TC-rich motif)等元件间的相互作用形成复杂的代谢调控网络来响应和应对重金属胁迫。这与水稻NRAMP基因受到JA、ABA等激素的调控来应对病原菌侵染、外界防御信号和金属离子,从而参与植物的防御反应一致(Zhou et al., 2004)。在对甘蔗ScNRAMP基因上游3 000 bp区域研究发现,成员中大部分均含有茉莉酸响应元件(TGACG-motif)和生长素响应元件(TGA-element),推断该基因家族在进化过程中可能具有与植物生长和逆境胁迫相关的功能。甘蔗ScNRAMP基因的表达具有组织特异性,甘蔗ScNRAMP基因主要在特定组织和器官中表达,Sspon.01G0025840-2B和Sspon.01G0061080-1D2位成员在种子发芽期的叶和茎中的表达量明显高于其他时期。对各时期和各组织高表达的基因分析发现,家族成员均具有MBS、ABRE和TCA-element等与激素响应相关的作用元件,推测其参与激素代谢过程进而影响甘蔗对重金属胁迫的响应。
目前,NRAMP基因家族在菜豆(Ishida et al., 2018)、大豆(Qin et al., 2017)和甘蓝型油菜(Meng et al., 2017)等经济作物上已被广泛研究。由于甘蔗作为重要的糖料作物和能源作物,关系着我国乃至世界人民的食糖安全,因此降低重金属胁迫对甘蔗产量及含糖量的影响,了解ScNRAMP等基因家族转运重金属离子的分子机制尤其必要。本文通过对甘蔗割手密中29个ScNRAMP基因家族成员的全基因组分析,有利于阐明甘蔗ScNRAMP基因家族的功能,并为甘蔗的分子育种尤其是应对重金属胁迫的研究提供了重要的候选基因。
猜你喜欢
新医学(2023年10期)2023-12-09 15:04:51
中学生天地(A版)(2023年1期)2023-02-17 00:33:04
儿童故事画报·自然探秘(2022年12期)2022-11-24 10:10:12
南方医科大学学报(2022年3期)2022-04-13 01:51:26
基层中医药(2021年3期)2021-11-22 08:08:04
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10 07:07:36
特别健康(2018年3期)2018-07-04 00:40:08
生命科学研究(2018年1期)2018-05-29 01:12:47
上海农业学报(2017年3期)2017-04-10 12:39:14
中国糖料(2016年1期)2016-12-01 06:48:57
