
miR-130a-3p调节有氧代谢提高小鼠运动耐力*
2022-11-28 12:30周浩吴佳韩陈婷罗君谊孙加节张永亮习欠云
中山大学学报(自然科学版)(中英文) 2022年6期
周浩,吴佳韩,陈婷,罗君谊,孙加节,张永亮,习欠云
华南农业大学动物科学学院/广东省动物营养调控重点实验室/国家生猪种业工程技术研究中心,广东 广州 510642
MicroRNAs(miRNAs)是一类在转录后水平参与基因调控并发挥重要作用的非编码小RNA,主要通过抑制翻译和降解mRNA 来调节其靶标[1]。30 多年前, 科学家们就在秀丽隐杆线虫Caenorhabditis elegans中发现了第一个miRNA,它最初被认为只是一个传统的蛋白质编码基因,但随着研究的深入,科学家们发现它并不编码蛋白质,而是编码了一个具有调节功能的22 个核苷酸的RNA,从此人类便开启了分子生物学的新纪元[2-4]。现在在人类中已有2 000 多种miRNA 被发现,据研究报道它们共同调节了人类基因组中超过1/3 的基因[5]。miRNAs 作为一个热门研究的调控生长因子,越来越多的研究表明,其在动物机体内能够参与细胞生长、分化、发育和凋亡等代谢活动,是细胞活动的强大调节者[6-8]。有报道称,miR-132 可通过靶向DAPK-1 刺激滋养层细胞的生长和侵袭[9];miR-125b-5p抑制3T3-L1前脂肪细胞增殖并促进成脂分化[10];miR-383-5p 通过靶向癌性PP2A 抑制胃癌细胞增殖并促进其凋亡[11]。同时,miRNAs 还与癌症、糖尿病、代谢性疾病等疾病的发病机制息息相关,可作为各种类型的疾病(尤其是癌症)的预测和诊断的生物标记物[12-14]。研究发现,miR-621 通过靶向TRIM29 通过Wnt/βcatenin信号通路抑制膀胱癌细胞的增殖和转移[15];miR-423 在各种人类肿瘤中异常表达,并参与癌症进展的多个信号通路[16];miR-26a 可通过改善外周胰岛素敏感性和保护β 细胞的功能来缓解2 型糖尿 病[17];miR-125a-5p 可 以 通 过 靶 向STAT3 抑 制肝脏脂肪生成和糖异生并提高糖原合成,从而改善2 型糖尿病肝糖脂代谢紊乱[18]。随着各界学者对miRNAs 功能研究的不断深入发现,miRNAs 与动物机体正常的生长、发育、繁殖等代谢活动密切相关。
近年来,有不少报道称miRNAs参与调节有氧代谢过程。研究表明,miR-181a/b-1 在成骨过程中通过增加线粒体耗氧率和与ATP 相关的呼吸代谢促进成骨分化[19];miR-107 通过促进线粒体β-氧化促进肝脏中脂质积累并降低葡萄糖耐受[20];miR-146a 过表达能促进肝细胞中的线粒体合成与线粒体呼吸[21];miR-21-5p 参与调节H9C2 细胞的线粒体呼吸和脂质含量[22];miR-33a/b 靶向抑制调节脂肪酸氧化调节的关键酶SIRT6,降低肝脏的脂肪酸氧化与胰岛素信号传导[23]。更有研究表示,有氧代谢向动物机体运动时骨骼肌的收缩提供了大量能量,而适度的有氧运动能增加线粒体的数量并减轻胰岛素抵抗[24]。
骨骼肌作为动物机体参与运动、氧化代谢等活动的重要代谢器官,其自身的发育及其对体内能量物质的利用受到外部环境因素以及内部调节因子的共同作用,包括miRNAs。早在2006年,研究显示miR-1 能促进成肌细胞分化,而miR-133 刺激成肌细胞增殖[25]。近来研究也发现,miR-664-5p 通过靶向血清反应因子SRF 和Wnt1 促进成肌细胞增殖并抑制成肌细胞分化[26];miR-17-92 促进C2C12 成肌细胞增殖但抑制分化[27];小鼠骨骼肌中miR-133a 的缺失会导致线粒体合成减少,运动耐力下降[28];miR-696 通过抑制PGC-1α 降低小鼠骨骼肌线粒体活性,并降低成肌细胞耗氧率[29];miR-494-3p 可以通过靶向E1A 结合蛋白P300 抑制快速氧化肌纤维的形成,并降低人骨骼肌的基础耗 氧 率[30];C2C12 肌 管 中miR-204-5p 的 沉 默 可通过调节PGC-1α 增强线粒体生物合成,同时miR-204-5p 可作为一种人类骨骼肌线粒体的功能调节剂[31];另有研究发现,miR-499 通过直接靶向Fnip1 激活肌细胞中的AMPK/PGC1α 信号,改变小鼠骨骼肌纤维类型,从而增强运动能力和线粒体活性[32]。上述资料表明,miRNAs 参与了骨骼肌发育以及机体氧化代谢的调节作用。miR-130a 作为miRNAs 的一员,其存在与功能在2006 年首次报道[33]。近年来,有关miR-130a 的研究不断深入,有报道表明miR-130a-3p 在非酒精性脂肪性肝炎纤维化的发展过程中可直接靶向TGFBR1和TGFBR2负性调节肝星状细胞活化和增殖[34];还可通过PTEN/PI3K/Akt 信号通路促进软骨细胞增殖和减轻骨关节炎[35],以及通过靶向SnoN 通过TGF-β1 通路抵抗肾脏纤维化[36]等。本实验室前期研究发现,miR-130a-3p 可以通过靶向PHLPP2 抑制小鼠脂肪合成代谢,并促进葡萄糖转运蛋白-4(GLUT4)转位,增加细胞对葡萄糖的吸收作用[37],同时还可靶向雌激素受体α(ERα)抑制催乳素(PRL)的表达[38]。研究提示,miR-130a 同样参与了物质代谢的调节,但其是否通过提高营养物质有氧代谢,进而影响骨骼肌的发育及其功能还未见报道。因此,本研究拟通过miR-130a-3p全身性敲除与过表达小鼠为对象,初步探究miR-130a-3p 对小鼠有氧呼吸及其运动能力的作用,为深入研究miR-130a-3p 对骨骼肌能量代谢和运动机能的调节机制提供前期基础。
1 材料与方法
1.1 试验动物
miR-130a-3p 全身性敲除与过表达FVB 小鼠由赛业(广州)生物科技有限公司通过CRISPR/CAS9基因编辑技术制备[37]。WT 小鼠为普通无特定病原体(SPF,specific pathogen free)级FVB 小鼠。小鼠料为普通SPF 级,符合国家标准,高脂饲料(HFD,60%的能量来自脂质),普通饲料(5%的能量来自脂质),购自于广东省医学实验动物中心(广东佛山)。试验动物的饲养和应用遵照《美国国立卫生研究院实验动物应用指南》进行。所有程序均获得华南农业大学实验动物伦理委员会的批准,并按照协议(SCAU-AEC-2016-0714)进行。
1.2 试验仪器
实时荧光定量PCR 仪(Bio-Rad C1000 Touch,广州市昊洋贸易有限公司);小鼠跑步机(瑞沃德生物科技有限公司);抓握力测定仪器(BIO-GS3,Bioseb,France);代谢测量系统(Sable Systems International,美国);分光光度计(Nanodrop 2000,Thermo Fisher,美国)。
1.3 试验设计与分组处理
选用30 只3 周龄生长性能相近的FVB 雄性小鼠,其中野生型(WT)、miR-130a-3p 过表达(130OE)与miR-130a-3p 敲除(130KO)小鼠各10只。在整个试验中,使用高脂饲料(HFD)喂养小鼠。所有小鼠均单笼饲养,每天光照和黑暗各12 h,自由采食和饮水。环境相对湿度控制在60%左右,温度控制在(25±1)℃左右。每3 d 记录1 次采食量,每7 d记录1次体质量。
1.4 小鼠肌肉力量与运动耐力检测
1.4.1 肌肉力量检测在试验的第5 周通过小鼠砝码抓握试验与抓握力测定试验检测肌肉力量。小鼠砝码抓握试验方法为:用手抓住小鼠尾巴把小鼠拎起来,迫使小鼠抓住一定质量的砝码,当小鼠能抓住砝码并坚持3 s 以上时则认为小鼠可以承受该质量,进入下一质量的砝码检测。每一只小鼠均轮流进行检测,测试结束后统计每组小鼠的得分情况[39]。抓握力测定试验通过将小鼠置于抓握力测定仪器中,每只小鼠轮流进行一共10 次的拉力试验,试验结束后去掉每只小鼠所测得的10 个数据中的最大值与最小值,取剩下数据的平均值用于统计分析。
1.4.2 运动耐力测试测试开始前将跑步机倾斜角度调为10°,每条跑道放入1 只小鼠,先以11 m/min 的速度运动,适应10 min 后休息10 min。然后开始试验:给予小鼠一个11 m/min 的初速度,每5 min加速4 m/min,直至小鼠静坐在电网上5 s,即判定为小鼠力竭,记录每只小鼠的运动时间与距离[40]。
1.5 活体有氧呼吸代谢测量
在饲喂高脂饲料的第6周,通过使用代谢测量系统得到小鼠的O2消耗量(VO2)、CO2产生量(VCO2)和产热量。
1.6 RNA提取与qRT-PCR
使用Trizol(Invitrogen)从组织中提取总RNA,使用分光光度计对RNA 的浓度进行检测。使用EZB 4 × EZscript Reverse Transcription Mix Ⅱ逆转录试剂盒(海方生物,上海)将总量1 μg的RNA进行逆转录,其中miR-130a-3p 使用特异性颈环引物逆转录,mRNA 使用OligodT18 逆转录。将得到的产物cDNA稀释5倍使用。
qRT-PCR 通 过 EZB 2× Color SYBR Green qPCR Master Mix(A0012)在实时荧光定量PCR 仪中进行检测,miRNA 使用U6作为内参,将得到的结果通过2-ΔCt方法进行统计。
1.7 数据统计分析
采用SPSS 24.0 进行单因素方差分析和独立样本t检验分析。所有统计分析均用Graghpad 7.0 统计软件完成,结果用平均值±标准误(mean±S.E.M)表示,以P<0.05作为差异显著性判断标准。
2 实验结果
2.1 miR-130a-3p 过表达和基因敲除转基因小鼠的鉴定
为了探究miR-130a-3p 是否参与调节骨骼肌发育以及机体的氧化代谢,我们建立了miR-130a-3p过表达(130OE)和miR-130a-3p 基因敲除(130KO)小鼠模型。在琼脂糖凝胶电泳中,130OE小鼠中可以检测到插入的外源基因(图1a);测序结果表明,与WT小鼠相比,130KO小鼠中缺失了miR-130a-3p的一段种子序列(图1b);通过qRT-PCR 检测小鼠体内的miR-130a-3p 发现,相对于WT 小鼠,130OE 小鼠的miR-130a-3p 表达量显著性升高(P<0.01),130KO 小鼠表达量显著下降(P<0.01)(图1c)。以上结果表明,miR-130a-3p 基因敲除与过表达小鼠模型已成功建立。

图1 转基因小鼠模型的构建Fig.1 Construction of transgenic mouse model
2.2 miR-130a-3p 过表达和敲除对小鼠采食量与体质量的影响
在高脂饲喂条件下,每3 d 测量1 次小鼠采食量发现,与WT 小鼠相比,130OE、130KO 小鼠的采食量均无显著差异(图2a),表明miR-130a-3p不影响小鼠的采食量;同时,从饲喂高脂饲料第2周开始,130KO 小鼠体质量显著大于WT 小鼠(P<0.05)(图2b);另外,在整个饲养期间,与WT 小鼠相比,130OE 小鼠的体增量显著降低(P<0.05),而130KO 小鼠的体增量显著升高(P<0.05)(图2c)。
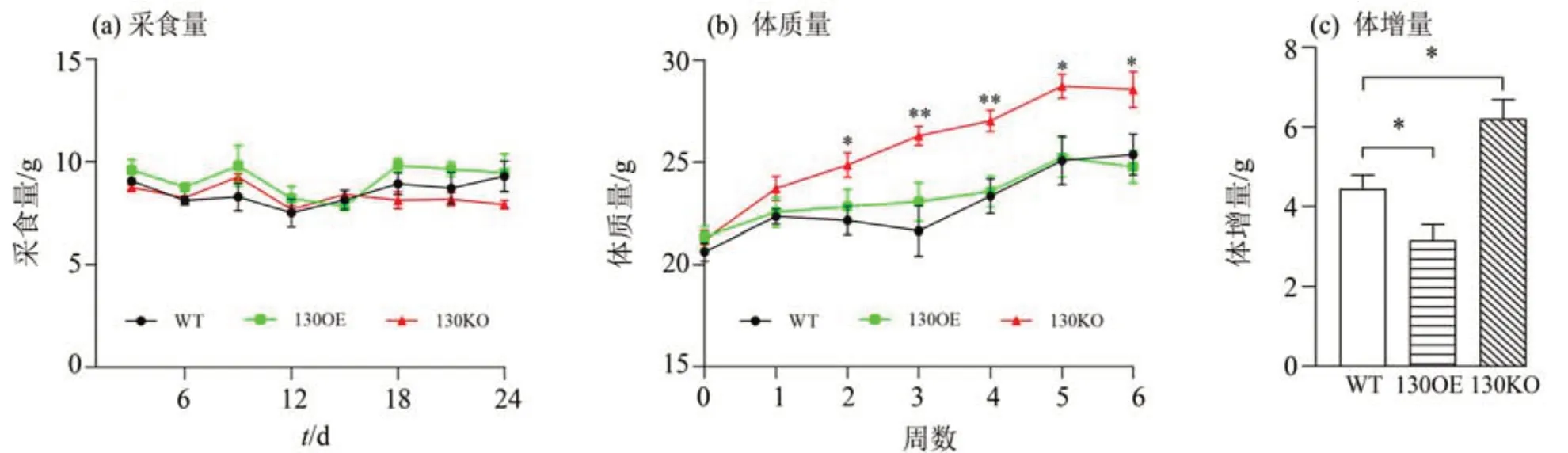
图2 过表达和miR-130a-3p敲除对小鼠采食量与体质量的影响Fig.2 Effects of overexpression and miR-130a-3p knockout on feed intake and body mass in mice
2.3 miR-130a-3p 过表达和敲除对小鼠有氧代谢的影响
随后我们在代谢笼中检测3种不同基因型小鼠的O2消耗量、CO2产生量以及产热量,由图3 可以得出,130KO 小鼠的O2消耗量显著低于WT 小鼠(P<0.01)(图3a);且相对于WT小鼠,130OE小鼠CO2产生量有显著上升(P<0.01),而130KO 小鼠的CO2产 生 量 显 著 下 降(P<0.01)(图3b);除此之外,130KO 小鼠的产热量显著低于WT 小鼠(P<0.01)(图3c)。以上结果表明,miR-130a-3p敲除时会降低小鼠的有氧呼吸代谢,而miR-130a-3p 过表达时也能在一定程度上提高小鼠的有氧呼吸代谢,说明miR-130a-3p 参与机体的有氧代谢活动。
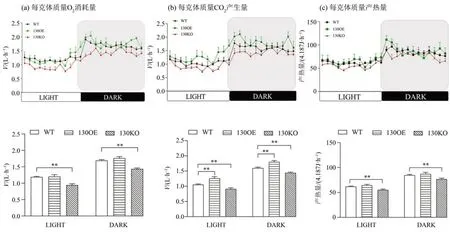
图3 miR-130a-3p敲除或过表达对小鼠有氧代谢的影响Fig.3 Effects of miR-130a-3p knockout or overexpression on aerobic metabolism in mice
2.4 miR-130a-3p 过表达和敲除对小鼠肌肉力量与运动耐力的影响
由图4 结果可知,与WT 小鼠相比,130OE、130KO 小鼠在砝码抓握试验的抓力得分(图4a)和抓握力测定试验(图4b)中均无显著差异,表明miR-130a-3p 不影响小鼠的肌肉力量。随后,我们通过跑步机测量小鼠的运动耐力发现,与WT小鼠相比,130OE 小鼠的运动时间与距离均显著上升(P<0.05),而130KO 小鼠的运动时间与距离均极显著下降(P<0.01)(图4c~d),表明miR-130a-3p可以提高小鼠的运动耐力。
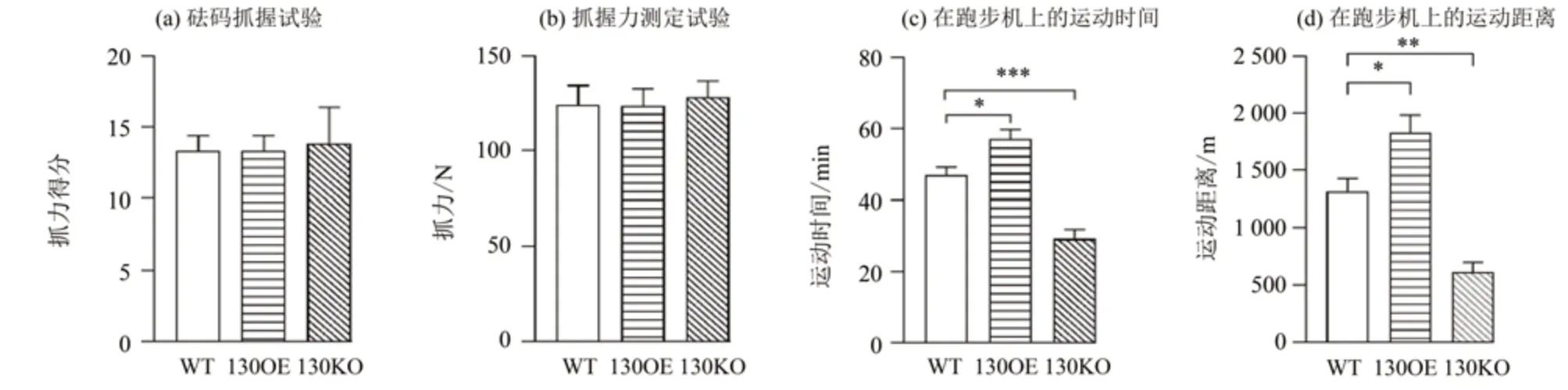
图4 miR-130a-3p过表达和敲除对小鼠肌肉力量与运动耐力的影响Fig.4 Effects of miR-130a-3p overexpression and knockout on muscle strength and exercise endurance in mice
3 讨论与结论
运动与有氧氧化代谢是研究肌肉相关功能的重要课题。骨骼肌作为机体运动的重要器官,其收缩的能量主要由无氧代谢途径和有氧代谢途径提供,而有氧代谢途径所产生的能量远高于无氧代谢途径,是骨骼肌最有效的ATP 来源[41]。骨骼肌由数以千计的具有收缩功能的肌细胞组成,被称为肌纤维[42]。在正常情况下,骨骼肌的肌纤维数目在动物出生之前就已经确定,出生后肌肉的生长发育主要体现在肌纤维类型之间的转变和大小变化,机体在整个生长发育过程中甚至发育成熟后都可能发生肌纤维类型的转变[43-44]。随着生物学的发展,肌纤维类型的分类方法也在不断演变,不同的肌纤维类型之间线粒体含量、ATP酶活性与有氧代谢酶活性均不相同[45]。目前根据肌纤维的代谢类型、代谢酶活性含量等,可将肌纤维分为氧化型和酵解型肌纤维。氧化型肌纤维具有更多的毛细血管与肌红蛋白,外观呈红色,因此也被称为“红肌”,含有较多的细胞色素、氧化代谢酶和线粒体,收缩速度较慢,属于慢收缩肌纤维[46-47]。而酵解型肌纤维的肌红蛋白和毛细血管较少,被称为“白肌”,细胞色素、氧化代谢酶和线粒体含量较少,收缩速度较慢,属于慢收缩肌纤维[48]。由于氧化型与酵解型肌纤维的种种不同,氧化型肌纤维比酵解型肌纤维表现出更强的抗疲劳能力。先前已有研究表明,骨骼肌纤维可以影响身体的耐力水平,优秀的长跑运动员的骨骼肌中含有较高水平的氧化型肌纤维比例[49]。其次,肌纤维的大小数量以及氧化型、酵解型的比例都与生理性衰老和病理性肌萎缩有关[50]。有报告指出,肌营养不良症患者的氧化型纤维含量较低,出现氧化型向酵解型转化的现象[51]。这些发现表明,肌纤维类型的比例与肌肉疾病密切相关。而在畜禽生产中,氧化型纤维比例更高的肉类由于肌红蛋白与血红蛋白的含量较高,具有更高的肉色评分与嫩度,肉品质更高。因此提高氧化型纤维的比例对于肌肉疾病的防控与改善畜禽肉品质都具有重要意义。
对于肌纤维之间转换的研究一直是近几年来对于骨骼肌相关研究的热点。研究表明,运动与否与不同的运动形式都能使肌纤维代谢类型之间发生转变,长时间的耐力运动能增强有氧氧化代谢相关酶活、增加肌纤维的线粒体含量,促进肌纤维的有氧氧化代谢[52]。也有研究报道,高脂饮食能够提高小鼠骨骼肌中线粒体含量从而显著提高跑步期间的运动耐力表现[53]。而近年来,随着对miRNAs的深入研究,肌纤维类型之间的转变同样有miRNAs 的参与,miR-152 通过靶向解偶联蛋白3(UCP3)基因促进慢肌纤维形成[54];miR-151-3p通过靶向骨骼肌细胞中的ATP2a2 调节慢肌基因表达[55];miR-22-3p 通过抑制AMPK/SIRT1/PGC-1ɑ信号通路调节骨骼肌纤维类型的转换[56]。目前对miR-130a 的研究报道主要围绕疾病与免疫、癌症等方面[57],研究表明,外泌体miR-130a-3p可以通过介导SIRT7/Wnt/β-catenin轴调节人脂肪干细胞的成 骨 分 化[58];Bta-miR-130a/b 通 过 靶 向 肉 牛 中 的PPARG 和CYP2U1 来 调 节 前 脂 肪 细 胞 分 化[59];miR-130a 和miR-27b 可以通过靶向PPARγ 促进人间充质干细胞的成骨分化[60];miR-130a 直接靶向抑制PPARγ减少肝脏脂肪的生成[61]。
已有报道表明miR-130a 参与了机体的发育与代谢调节,但其是否参与骨骼肌的发育还未见报道。碳水化合物作为骨骼肌在长时间(耐力型)运动中氧化的主要燃料,其来源在很大程度上取决了运动强度和持续时间,随着运动强度的增加,碳水化合物的贡献更大,通过摄入碳水化合物来提高长时间运动中碳水化合物的利用率一直是运动营养研究领域的主导[62]。有研究报道,氧化型肌纤维比酵解型肌纤维具有更高的葡萄糖敏感性与GLUT-4 蛋白表达,在基础和胰岛素刺激条件下能够摄取更多葡萄糖[63]。前期研究表明miR-130a-3p能够促进GLUT4 的转位并增加细胞对葡萄糖的摄取[37]。本研究结果显示miR-130a-3p过表达小鼠具有更高的运动耐力,敲除小鼠则反之。代谢测量结果也显示,miR-130a-3p 过表达小鼠具有更强的有氧代谢,而敲除小鼠则反之。研究结果提示,miR-130a-3p 能够增强小鼠的有氧代谢,从而提高其运动耐力,但具体的调控机制尚不清楚。我们推测miR-130a-3p 可能通过促进骨骼肌对葡萄糖的利用,调节肌纤维类型转换,提高小鼠骨骼肌氧化型纤维的比例,增强运动耐力。
本文以miR-130a-3p 全身性敲除与过表达小鼠为对象,通过分子生物学、在体生理指标测量等技术,初步探究了miR-130a-3p 对小鼠有氧呼吸及其运动能力的作用。研究显示miR-130a-3p 具有提高小鼠有氧代谢、降低体质量、增强运动耐力的作用,其结果为深入研究miR-130a-3p 对骨骼肌能量代谢和运动机能的调节机制提供了前期基础。
猜你喜欢
中老年保健(2022年3期)2022-11-21
中老年保健(2022年2期)2022-08-24
中老年保健(2022年4期)2022-08-22
中国临床医学影像杂志(2022年6期)2022-07-26
中国临床医学影像杂志(2022年5期)2022-07-26
国际放射医学核医学杂志(2021年10期)2021-02-28
食品安全导刊(2018年36期)2018-05-25
天然产物研究与开发(2018年2期)2018-04-04
中国运动医学杂志(2016年3期)2016-07-10
中国运动医学杂志(2016年3期)2016-07-10
