
基于等转化率法和广义主图法的植鞣皮革热解动力学研究
2022-11-19 06:35张铭芮王殷菲王猛王芳刘捷linaGeorgianaAlbuKaya汤克勇
皮革科学与工程 2022年6期
张铭芮,王殷菲,王猛,王芳,刘捷,Mădălina Georgiana Albu Kaya,汤克勇*
(1.郑州大学材料科学与工程学院,河南 郑州450001;2.Leather and Footwear Research Institute,Collagen Department,93 Ion Minulescu,Bucharest 031215,Romania)
前言
皮革文物是传承人类文明、见证历史发展的重要载体。通过研究皮革文物老化的机理,可以了解其的组成、结构、性能与当时的制造工艺。植鞣是最古老、使用最广泛的制革方法,也是皮革文物中最常见的鞣制工艺[1-3]。因此,研究植鞣皮革的老化机理对于文物保护领域具有重要意义。
植鞣皮革是指生皮经植物单宁鞣制而得的皮革。植物单宁中含有大量酚羟基,可与生皮中胶原大分子链形成氢键、发生交联,永久地改变与稳定胶原的结构,从而实现从生皮到皮革的转化[4-5]。单宁是由高等植物合成的复杂且异构的多酚类次生代谢产物,相对分子质量在500至3000 Da之间。根据其化学结构,通常可将其分为水解类单宁和缩合类单宁[6]。水解类单宁由棓酸或由棓酸衍生的多元酚羧酸与糖(主要是d-葡萄糖)或其他物质(如多元醇)通过酯键或糖苷键结合而成的复杂化合物的混合物(如图1a所示),在稀酸、稀碱、酶作用下可水解为多元酚羧酸和糖或多元醇,常见的水解类单宁主要来自于栗木栲胶、塔拉栲胶、橡椀栲胶等;缩合类单宁是分子中的所有芳香核以碳碳键连接的多酚类物质,主要来自于坚木栲胶、落叶松栲胶、杨梅栲胶等,其核心结构化学式如图1b所示。
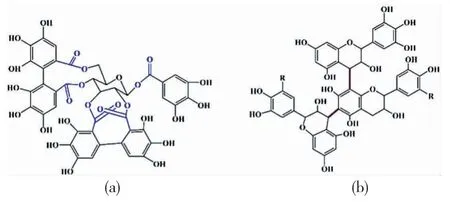
图1 植物单宁的核心结构单元(a)水解型单宁(b)缩合型单宁Fig.1 The core structure units of(a)hydrolysable tannins(b)condensed tannins
热重(TG)分析是研究物质热性能和热解动力学的有力手段,具有样品需求量少、灵敏度高、对样品的物理状态无特殊要求、研究试温度范围宽等优点,符合文物取样的特殊性要求。Budrugeac等通过TG/DTG分析了新制备皮革和皮革文物在静态空气环境中的热降解过程。结果表明,它们的热解过程可分为两个阶段,分别对应水分损失和氧化降解。皮革文物的热氧化降解率低于新制备皮革,因此热氧化速率可用于皮革老化的定性分析[7]。Luo等人通过TG研究了在氮气气氛下,从室温到600℃的不同升温速率下,绵羊毛皮粉末废弃物的热解过程,将其热解分为三个阶段,并通过改进的Kissinger-Akahira-Sunose(MKAS)方法计算,得到主热解阶段的总活化能(E)为275.6 kJ/mol[8]。尽管多加热速率下的TG分析非常适合研究各种材料的热裂解动力学,但单一的技术很难提供完整的信息,因为植物鞣革中含有植物鞣质、胶原蛋白成分和非胶原蛋白成分,使其热解过程更加复杂。Hu等人将TG与傅里叶变换红外光谱相结合分析了醛鞣皮革热解过程中的逸出气体,发现醛鞣不会显著改变热解过程中逸出气体的组成[9]。
本文通过TG及将TG与傅里叶变换红外光谱和质谱仪的分析手段相结合,从分子水平上获得研究了植鞣皮革宏观性能和微观结构之间的关系,并通过分析热解过程中逸出气体的成分,揭示了其老化的机理,可望为皮革文物的保护与修复提供重要的理论依据。
1 实验部分
1.1 主要材料与仪器
浸酸羊皮,购于隆丰皮草企业有限公司(焦作,河南)。用7%的氯化钠溶液和3%的硫代硫酸钠溶液调节羊皮pH至7.0后备用。
塔拉栲胶和坚木栲胶,由郑州恒镕泽商贸有限公司提供。在8 h内,分别将两种栲胶分三次(分别为5%、15%、25%)投入转鼓鞣制皮革。鞣制结束后,用蒸馏水洗涤以去除样品表面和未与胶原结合的鞣剂,得到鞣制后的样品,分别命名为塔拉鞣革和坚木鞣革。
蒸馏水,实验室自制(北京浦肯野通用仪器有限公司,GWB1/1super/2,中国)。
1.2 热重分析实验
采用热重分析仪TGA/DSC-1(瑞士梅特勒托利多集团)进行TG测试。称取未鞣制羊皮和塔拉单宁、坚木单宁鞣制的皮革试样4~5 mg,分别以5、10、20℃/min的升温速率从室温升至600℃。实验气氛为氮气,气体流速为40 mL/min。
1.3 热重-红外光谱-质谱(TG-FTIR-MS)联用分析
采用热重分析仪TGA/DSC(STA499F3 Jupiter,NETZSCH,德国)与红外光谱仪(Bruker Tensor II,德国)和质谱仪(PFEIFFER,OmniStarTM,德国)联用,进行TG-FTIR-MS测试。称取未鞣制羊皮、塔拉鞣革和坚木鞣革试样约8 mg,置于氧化铝坩埚中,在流速为60 mL/min的氮气气氛下,以10℃/min的升温速率从室温加热至800℃。热重分析仪、红外光谱仪和质谱仪连接附件的温度保持在200℃,以防止逸出气体在连接管道中冷凝。
1.4 动力学模型
热分析方法是在程序控制温度的条件下,研究材料的物理性质随着温度变化的技术[10-12]。通常情况下,将处理的反应过程物理性质变化的过程称为转化率α,以质量的变化来表示:

其中,m0是初始质量,mt是时间t的质量,mf是最终质量。反应的速率通过Arrhenius方程参数化,可用式(2)来表示:

其中,E是活化能,A是指前因子,R是气体常数(8.314 J/mol·K)。在非等温条件下,引入升温速率,则对应的固体反应速率可表示为:
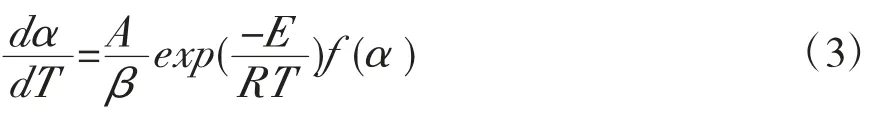
可以采用等转化率法计算活化能,本文采用FR与FWO法,其对应的公式为FR[11]:
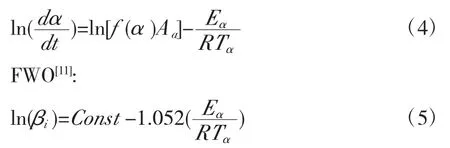
引入广义时间概念θ,广义时间的参数定义为:

以α=0.5为参考点,对式(6)进行微分,将其带入式(2),可得理论模型主曲线方程:

结合式(7),可建立与实验数据的主曲线方程:
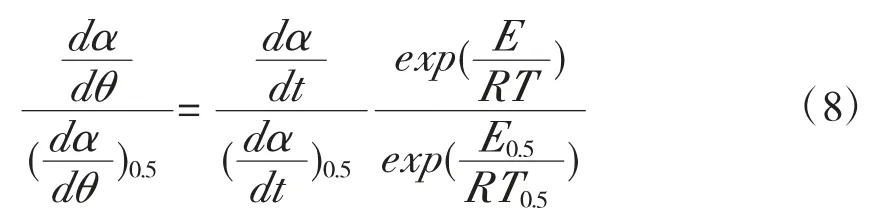
1.5 机理模型函数
计算动力学模型函数的动力学指数和动力学指前因子A,主要用到表1对应的动力学模型:随机成核和核生长机理,An;扩散控制机理,Dn;相边界反应机理,Rn;简单级数反应机理,Fn。其具体对应公式详见表1。
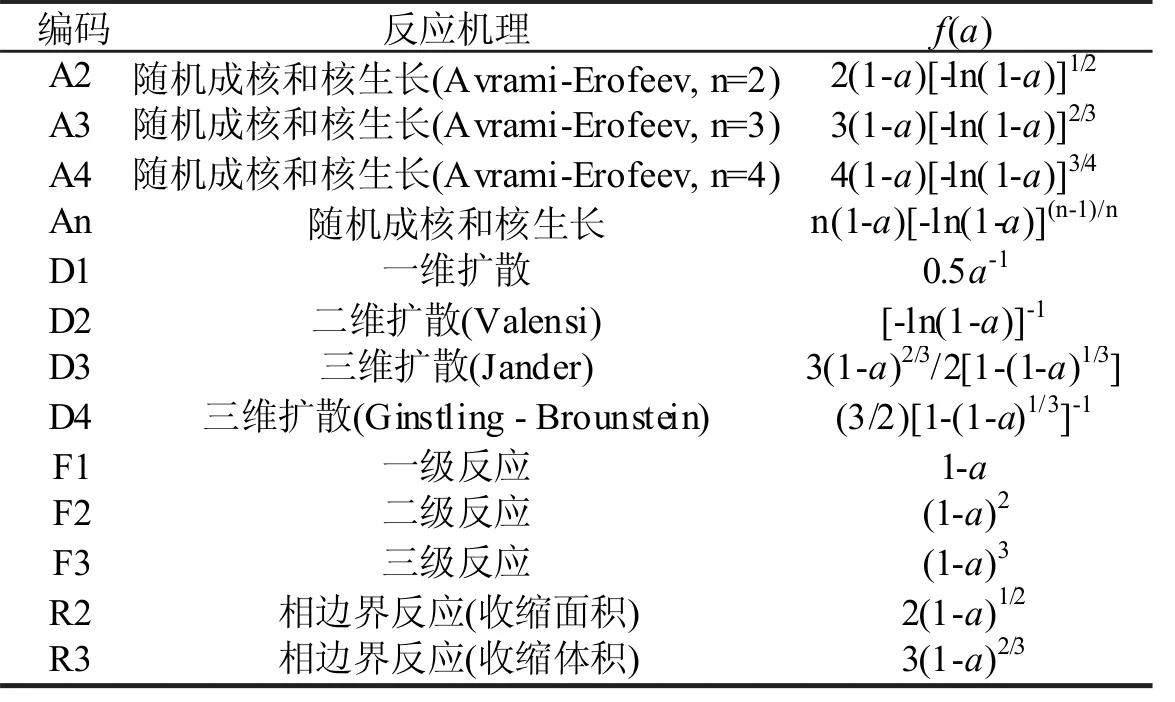
表1 固体材料降解的常用的反应机理模型[11]Tab.1 The commonly used theoretical kinetic models of solid-state reactions[11]
2 结果与讨论
2.1 TG分析
图2为在20℃/min的升温速率下,未鞣制羊皮、塔拉栲胶和塔拉鞣革、坚木栲胶和坚木鞣革的TG和DTG曲线。这三个样品的整个热解过程可分为三个阶段:第一阶段(StageⅠ,35~200℃)的失重主要是其中自由水和结合水等小分子物质的挥发;第二阶段(StageⅡ,200~500℃)发生了显著的质量损失,对应羊皮和皮革中胶原蛋白的降解[13]。未鞣制羊皮、塔拉鞣革和坚木鞣革的最大失重率分别为49.8%、44.6%和43.6%。未鞣制羊皮的最大质量损失率高于其他两种鞣制后的皮革,表明这两种栲胶的鞣制提高了皮革的热稳定性。DTG曲线显示了其两个典型的热解阶段。鞣制皮革的最大热解峰低于未鞣制的羊皮,这可能是因为鞣制过程中,鞣剂的进入破坏了皮革中部分胶原蛋白肽链之间的非共价键,造成了皮革内部鞣制结构的不均匀性。另一方面,塔拉和坚木栲胶的最大降解温度分别为297.7℃和316.6℃,这也会导致鞣制后皮革的最大热解峰向低温方向移动。第三阶段(StageⅢ,500~600℃)的质量损失对应于其热解残渣中含碳物质的缓慢分解与碳化。

图2 (a)未鞣制羊皮(b)塔拉栲胶和塔拉鞣革(c)坚木栲胶和坚木鞣革的TG和DTG曲线Fig.2 The TG and DTG curves of(a)untanned sheepskin,(b)tara and tara-tanned leather,and(c)quebracho and quebracho-tanned leather
2.2 TG-FTIR-MS分析
图3(a,c,e)所示为对未鞣羊皮、塔拉鞣革和坚木鞣革在氮气气氛中热解产生的逸出气体进行实时检测得到的3D-FTIR光谱图。图3(b,d,f)中,质荷比(m/z)为16、17、18、28、43、44和67的离子碎片分 别 对 应 于CH4、NH3、H2O、CO、HNCO、CO2和吡咯。CO2的逸出主要归因于胶原蛋白中谷氨酸和天冬氨酸中游离羧基的脱羧、肽键之间的缩合反应,以及其他有机基团的内部氧化[14]。在350℃时,可以观察到塔拉鞣革出现CO峰值,而坚木鞣革没有,这可能是由塔拉栲胶鞣制后引入的羰基在热解过程中通过脱羧生成的,可以此作为植鞣皮革鞣剂的种类的判定依据。NH3的生成通常来自于游离氨基和肽链的脱氨反应[15]。与未鞣制的羊皮相比,塔拉鞣革和坚木鞣革中NH3的逸出量均有减少,表明鞣剂与胶原发生交联起到鞣制作用的原理是其与胶原中含氮基团的结合,将其中的含氮基团固定的结果。CH4通常由富含的甲基(-CH3)和亚甲基(-CH2-)分解而成。与未鞣制的羊皮相比,鞣制后皮革热解产生的CH4和NH3的强度均呈下降趋势,因此可将其用于鉴别皮革鞣制的依据。吡咯和HNCO是胶原蛋白热解的特征气体,吡咯主要是其中胶原的脯氨酸和羟脯氨酸的热解产物,在300~500℃范围内检测到微量逸出,说明此时胶原的结构完全破坏了。
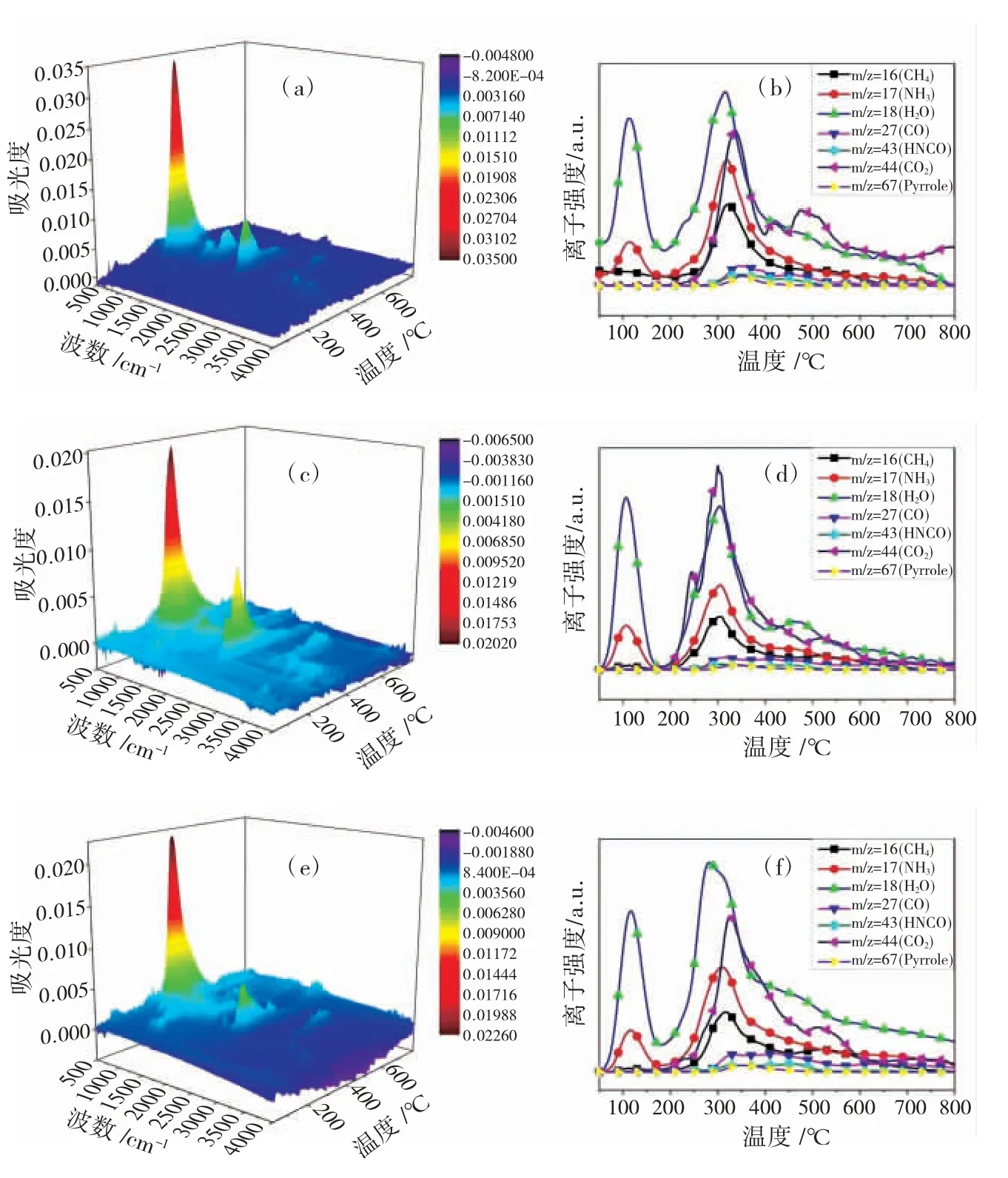
图3 (a,b)未鞣羊皮、(c,d)塔拉鞣革和(e,f)坚木鞣革的3D TG-FTIR谱图和逸出气体离子丰度分布图Fig.3 3D FTIR spectra of gaseous products and ion abundance distributions of gaseous products during pyrolysis of(a,b)untanned sheepskin,(c,d)tara-tanned leather,and(e,f)quebracho-tanned leather
2.3 动力学分析
2.3.1 等转化法分析
在动力学分析中,选取与分析了试样在200~500℃之间的热解行为。根据FWO和FR方法,绘制lnβ与1000/T和ln(dT/dα)与1000/T的曲线,如图4所示。在α=0.1~0.8的范围内,采用最小二乘法获得的线性拟合直线的相关系数(R2)均大于0.95,因此计算所得的活化能(Eα)具有准确性。然后,通过FWO法和FR法分别计算得到未鞣制羊皮、塔拉鞣革和坚木鞣革的平均活化能分别为209.4、222.4、238.7 kJ/mol以 及239.5、254.9、270.2 kJ/mol。根据计算两种方法的平均活化能偏差(εE,εE=|E-E0|×100/E0,%)如图5所示。Vyazovkin等人[11]指出,当活化能的εE小于20%~30%时,尽管热解过程实际可能包含多步反应,仍可视为单一反应过程进行动力学分析,因此选用FWO法进行后续拟合。
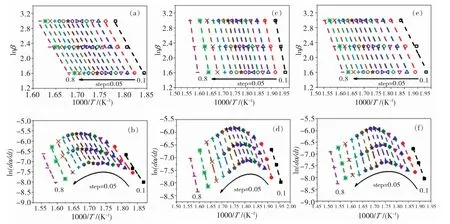
图4 (a,d)羊皮、(b,e)塔拉鞣革和(c,f)坚木鞣革的FWO和FR法对应的lnβ与1000/T和ln(dα/dt)与1000/T的关系曲线Fig.4 Plots of lnβ vs.1000/T and ln(dα/dt)vs.1000/T by methods of FWO and FR of(a,d)untanned sheepskin,(b,e)tara-tanned leather,and(c,f)quebracho-tanned leather

图5 (a)羊皮、(b)塔拉鞣革和(c)坚木鞣革的FWO和FR法对应的Eα和εE Fig.5 Eα和εE by methods of FWO and FR of(a)untanned sheepskin,(b)tara-tanned leather,and(c)quebracho-tanned leather
2.3.2 动力学模型分析
由式(8)计算所得到的未鞣制的羊皮、塔拉鞣革与坚木鞣革对应的实验主图不与任何一个理论主曲线模型相重叠,如图6所示。考虑到皮革中胶原蛋白交联的不均匀性以及皮革中鞣剂的存在,本文为简化起见,假定未鞣制的羊皮中有两种假组分,塔拉鞣革与坚木鞣革中有三种假组分,并将其DTG曲线进行接卷积处理,处理前后如图7所示。根据解卷积处理后所得到的假组分数据,计算其动力学参数,绘制对应的实验主图,如图8所示。通过主图的研究,发现假组分均符合随机成核和核生长模型,且曲线位于A3与A4之间。选取0.1间隔,拟合最佳的反应级数。其对应的动力学参数以及反应函数如表2所示。

表2 羊皮、塔拉鞣革和坚木鞣革假组分的动力学参数与反应函数Tab.2 Kinetic parameters and reaction functions of pseudocomponents of sheepskin,tara-tanned leather,and quebracho-tanned leather

图6 (a)羊皮、(b)塔拉鞣革和(c)坚木鞣革的实验和理论模型广义主曲线图Fig.6 Experimental curves and theoretical curves of(a)untanned sheepskin,(b)tara-tanned leather,and(c)quebracho-tanned leather

图7 (a)羊皮、(b)塔拉鞣革和(c)坚木鞣革的DTG曲线及解卷积分析图Fig.7 The deconvoluted DTG curves of(a)untanned sheepskin,(b)tara-tanned leather,and(c)quebracho-tanned leather

图8 (a,b)羊皮、(c,d,e)塔拉鞣革和(f,g,h)坚木鞣革假组分的实验和理论模型广义主曲线图Fig.8 Experimental curves and theoretical curves for pseudocomponents of(a,b)untanned sheepskin,(c,d,e)tara-tanned leather,and(f,g,h)quebracho-tanned leather
3 结论
本文对未鞣制的羊皮、塔拉单宁鞣制皮革和坚木单宁鞣制皮革,以不同升温速率进行了热重分析,研究与对比了其热解特性及其动力学反应模型,计算得到了其热解活化能。研究结果表明,未鞣制羊皮、塔拉单宁鞣制皮革和坚木单宁鞣制皮革均符合An模型,n分布于3.1~3.8之间,说明其热解是一个复杂的热化学反应过程。基于等转化率法和广义主图法的热解分析方法有望为皮革文物的鉴定和保护提供理论基础,热解过程中CO的逸出信息及CH4和NH3强度的变化为皮革鞣制方法和单宁类型的鉴定开辟了一条新的途径。
猜你喜欢
酿酒科技(2022年5期)2022-05-24
散文诗(2021年22期)2022-01-12
林业科技(2020年3期)2021-01-21
中国酿造(2020年12期)2021-01-10
小学阅读指南·低年级版(2020年11期)2020-11-16
中国果菜(2019年11期)2019-12-14
小学阅读指南·低年级版(2016年11期)2017-02-06
中南林业科技大学学报(2016年6期)2016-12-19
小学教学研究·新小读者(2016年9期)2016-09-21
中华家教(2016年5期)2016-05-14
