
中国家鹅血管紧张素转化酶2(ACE2)基因的克隆、表达及序列解析
2022-10-24 02:46张崇昊李志强谢娜娜曹西月张源淑
中国兽医学报 2022年7期
张崇昊,李志强,谢娜娜,曹西月,张源淑
(南京农业大学 动物医学学院 农业部动物生理生化重点开放实验室,江苏 南京 210095)
在最近暴发的新型冠状病毒肺炎(coronavirus disease 2019,COVID-19)中,已有的研究认为其病原SARS-CoV-2与SARS-CoV相同,均以ACE2作为受体而入侵机体[9]。SARS-CoV-2在进入宿主细胞时,其囊膜上的S蛋白被切割为S1和S2 2个亚基,S2亚基不与受体相互作用,但参与宿主细胞膜融合。S1蛋白含有受体结合域(receptor binding domain,RBD),能够与ACE2的肽酶结构域结合从而进入宿主细胞[10]。并发现ACE2中含有20个氨基酸残基与SARS-CoV-2的S蛋白受体结合域接触,其中8个氨基酸能与S蛋白形成氢键,其他氨基酸主要通过疏水作用与S蛋白相互作用,这些氨基酸残基的改变可能会引起ACE2与SARS-CoV-2的亲和性改变[11]。因此,比较不同动物ACE2中与SARS-CoV-2接触的氨基酸残基变化,可能有助于筛选SARS-CoV-2潜在的中间宿主。
目前,ACE2的研究主要集中在人医临床和哺乳动物,对于家禽特别是鹅的研究仍较为匮乏。本试验成功克隆出中国家鹅ACE2的基因序列,并运用生物信息学分析方法,对ACE2编码氨基酸的同源性及其分子进化、ACE2蛋白质理化性质、二级结构及相关参数进行了预测与分析,并将鹅与人以及其他动物ACE2的关键氨基酸组成进行了比较,对鹅是否易感SARS-CoV-2做出一定的分析,以期为鹅ACE2相关研究提供基础理论资料。
1 材料与方法
1.1 组织样本将16日龄中国家鹅颈部放血处死后,取心脏、肾脏、肺脏、十二指肠、空肠、回肠等组织,经生理盐水漂洗后放入液氮中,后移至-80℃冰箱保存。
1.2 细胞及主要试剂DH5α感受态细胞、TRIozal试剂均购自南京Vazyme Biotech公司;RNA反转录试剂盒购、Master Mix均购自日本TaKaRa公司;DL5000 DNA Maker、pMD-19T(含SolutionⅠ试剂)载体均购自宝生物工程(中国,大连)有限公司;XhoⅠ酶购自南京全式金公司。
1.3 总RNA的提取及反转录随机取80 mg鹅心脏、肺脏、肾脏、十二指肠、空肠、直肠组织,充分匀浆后按TRIzol说明书提取各组织总RNA,检测浓度和纯度后,选取D260 nm/D280 nm比值为1.8~2.0的样品采用二步法反转录,具体操作按反转录酶试剂盒说明书进行。产物cDNA于-20℃冰箱中保存。
1.4 引物设计利用Primer Premier 5软件,根据GenBank预测鹅ACE2基因序列自行设计引物,引物由南京擎科生物科技有限公司合成。具体引物序列如下:ACE2-F 5′-CGGAATTCATGTTGGCTCACGTCTTGCTTC-3′,ACE2-R 5′-CCCTCGAGAAAACAGAAACCCTTATTGTCCCATGT-3′。
汽车发动机电控系统维修、汽车自动变速器维修、汽车电气系统维修等课程的核心知识和操作技能,如果仅凭教师的讲授和演示,学生很难真正理解和掌握。教师普遍采用单一的课堂讲授的形式,有的会配合多媒体及案例进行讲授,但学生反映还是理解困难,学习积极性差,教学效果很难得到保证。
1.5 PCR扩增及扩增产物回收以cDNA为模板进行PCR扩增,反应体系(50 μL):3dH2O 22 μL,Master Mix 25 μL,上、下游引物各1 μL,cDNA 1 μL。反应条件:94℃预变性3 min;95℃变性30 s,60℃退火30 s,72℃延伸2 min 30 s,72℃彻底延伸5 min,33个循环。PCR扩增结束后,取10 μL产物进行琼脂糖凝胶电泳检测。依据DNA胶回收试剂盒说明书回收DNA琼脂糖凝胶并进行纯化,纯化后产物于-20℃冰箱中保存。
1.6 pMD-19T-ACE2重组质粒的构建及感受态转导参照文献[12]的方法进行。重组条件:16℃运行14 h获得重组质粒。按照DH5α感受态细胞说明书将重组质粒导入DH5α感受态细胞,重悬后涂布在含氨苄青霉素的LB固态培养基上,生长出的单克隆菌落用于菌液PCR鉴定。
1.7 目的基因的鉴定随机挑取单克隆菌落加入到含500 μL LB液态培养基的EP管中,37℃震荡培养5 h。扩大培养后取菌液进行PCR扩增和琼脂糖凝胶电泳验证,步骤同1.3。用小型质粒提取试剂盒提取质粒,用XhoⅠ酶对提取质粒进行单酶切验证,1%琼脂糖凝胶电泳。将鉴定阳性的质粒送上海英俊生物技术有限公司测序。
1.8 中国家鹅ACE2基因生物信息学分析使用NCBI网站中的ORF Finder功能查找开放阅读框;利用NCBI网站Blast软件进行核苷酸和氨基酸的同源性比对分析;利用ClustalX和MEGA6.0软件的邻接法(neighboe-joiningmethod,NJ)构建系统发育树;利用在线软件(http://web.expasy.org/protparam/)以及DNA Star里的Protean分析蛋白质理化性质;利用JPred4在线软件预测二级结构;利用TMHMM在线软件(http://www.cbs.dtu.dk/services/TMHMM/)进行鹅ACE2蛋白质跨膜结构分析;利用SignalP在线软件(http://www.cbs.dtu.dk/services/SignalP-4.0/)进行蛋白质信号肽分析;利用NetPhos2.0Server(http://www.cbs.dtu.dk/services/NetPhos/)对ACE2基因编码蛋白质进行磷酸化位点预测。
2 结果
2.1 中国家鹅ACE2基因表达及扩增电泳结果将鹅不同组织中提取的cDNA进行PCR扩增,后经1%琼脂糖凝胶电泳,结果如图1所示。在鹅十二指肠、直肠和肾脏组织中出现单一条带(泳道4、8、14),条带在2 000~3 000 bp之间,与预期(2 473 bp)大小一致。

图1 鹅不同组织中ACE2基因的表达电泳图
2.2 中国家鹅ACE2基因克隆结果
2.2.1菌液PCR验证结果 随机挑取5个阳性菌落,对其菌液进行PCR扩增和琼脂糖凝胶电泳,结果如图2所示,泳道6在2 000~3 000 bp之间出现单一条带,插入载体的目的基因片段与PCR扩增结果一致,提示阳性菌落中目的基因片段插入成功。
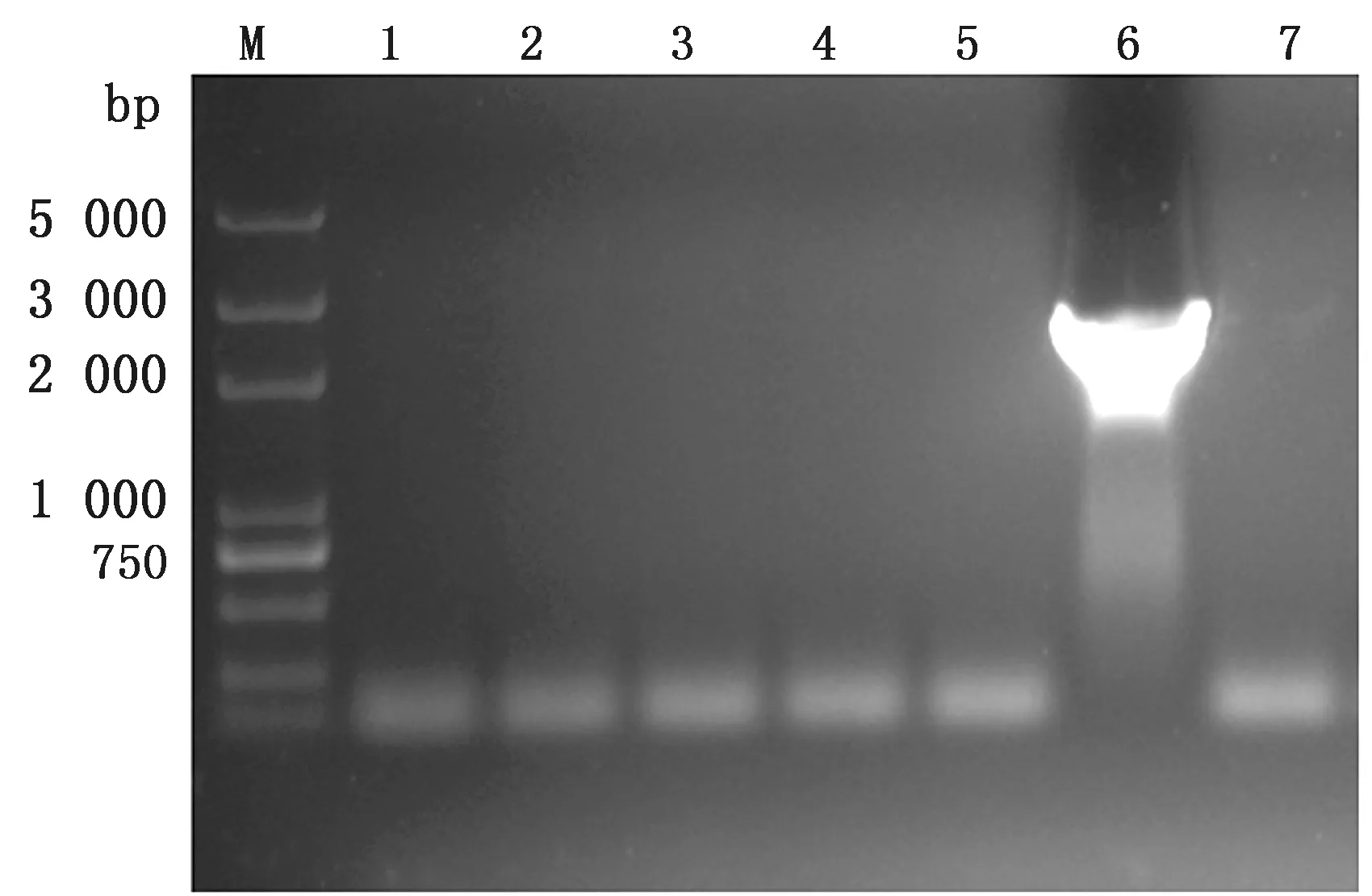
图2 菌液ACE2基因PCR扩增产物
2.2.2酶切验证结果 对pMD-19T-ACE2进行酶切验证,结果如图3所示。XhoⅠ单酶切后发现有1条约5 165 bp的条带,大小与pMD-19T载体(2 692 bp)和目的基因(2 473bp)总和一致。进一步说明重组质粒构建成功。

图3 pMD-19T-ACE2重组质粒单酶切鉴定结果
2.2.3家鹅ACE2基因测序 利用Sanger测序法对阳性质粒测序,结果显示本试验所克隆鹅ACE2基因核苷酸序列全长为2 473 bp,与预期结果(2 473 bp)一致,表明克隆成功。测序结果已提交到GenBank,序列号为MW355441。
2.3 中国家鹅ACE2基因序列的生物信息学解析
2.3.1ACE2开放阅读框(ORF)及编码氨基酸组成分析 利用ORF Finder进行分析,结果显示本试验所得中国家鹅ACE2基因ORF长度为2 473 bp,共编码808个氨基酸(图4A)。与GenBank上提供的白羽肉鸡ACE2氨基酸数(808个氨基酸)相同,比人、褐家鼠(805个氨基酸)多编码3个氨基酸,比牛、羊(804个氨基酸)多编码4个氨基酸。
对其基因编码氨基酸组成进一步分析,结果如图4B所示,该中国家鹅ACE2共含有20种氨基酸,其中Leu含量最高,占氨基酸总量的8.8%,Glu、Ala含量次之,分别为7.9%和7.7%,Cys含量最少,仅占1.2%。

A.鹅ACE2基因编码氨基酸序列;B.鹅ACE2基因编码氨基酸组成
2.3.2氨基酸同源性及分子进化分析 通过DNAStar软件中的MegAlign 模块,将所得中国家鹅 ACE2 氨基酸序列与 GenBank 提供的其他动物的序列进行同源性分析(图5A),并绘制遗传进化树(图5B)。结果显示,该鹅ACE2氨基酸序列与中国麻鸭的同源性最高为93.1%,并处于同一进化分支。与白羽肉鸡的同源性为88.6%。与山羊、牛的同源性最低,分别为42.3%和38.5%,处于完全不同的进化分支。
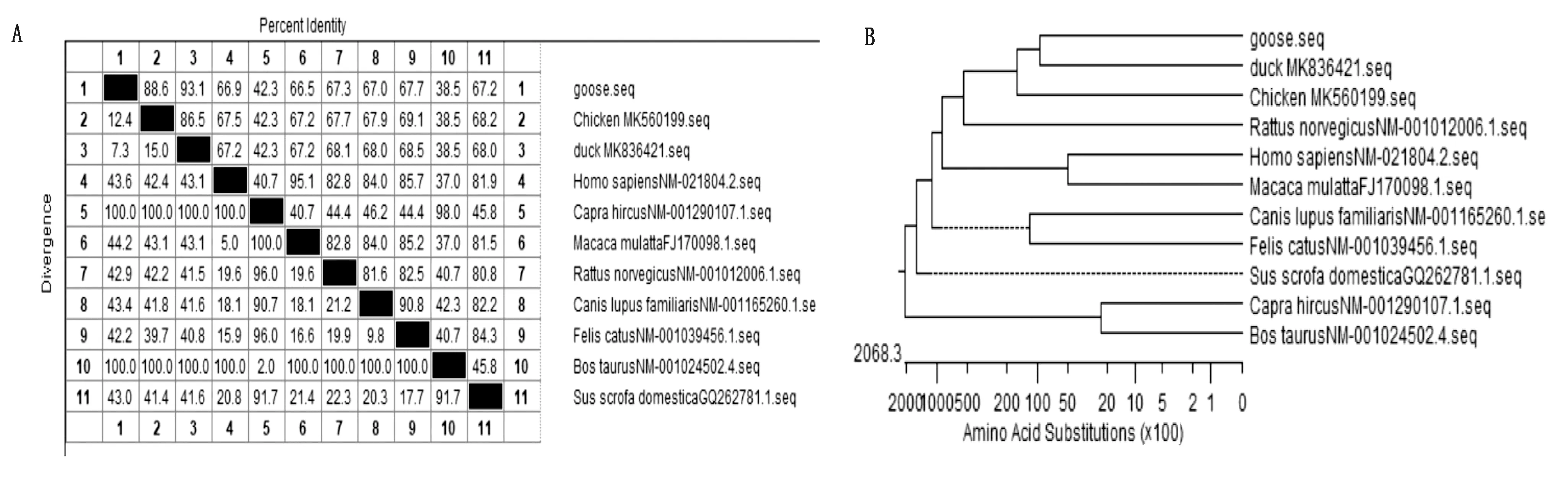
A.ACE2氨基酸序列同源性分析结果;B.ACE2基因序列分子进化分析结果
2.4 中国家鹅ACE2基因编码蛋白质分析
2.4.1理化性质分析 利用在线软件(http://web.expasy.org/protparam/)以及DNA Star里的Protean分析,该鹅ACE2基因编码蛋白质由808个氨基酸残基组成,其分子式为C4170H6357N1087O1241S37,原子总数为12 892,相对分子质量为92 760.11。理论等电点(pI)为5.14。脂溶指数为79.44,总平均亲水指数-0.346,属于亲水蛋白质。带负电荷的氨基酸残基(Asp+Glu=108)大于带正电荷的氨基酸残基(Arg+Lys=78),其摩尔消光系数为175 725 L/(mol·cm),1A(280)=1.894 g/L。
2.4.2二级结构及亲水/疏水性分析 ACE2蛋白质二级结构及亲水/疏水性分析结果如图6所示。该鹅ACE2蛋白质有32处α螺旋,在第Ala617~Asp669处出现最大α螺旋,分布均匀,且绝大部分由多个氨基酸组成,同时有18处β折叠存在,在第Ile436~Glu467、Pro737~Gly764处有最大β折叠,在各β折叠单元之间存在长短不一的转角,β转角区域分布较为分散,可见较多单个氨基酸出现。组成ACE2蛋白质二级结构的α螺旋(HELIX,H)、β折叠(STRAND,E)、无规卷曲(LOOP,L)的百分比为39.51%∶22.22%∶38.27%。

图6 ACE2蛋白质二级结构及亲水/疏水性分析
此外,该蛋白质存在较多的亲水性区域,较少的疏水性区域,其中亲水性较高的区域主要集中在Ile155~Tyr251和Gly764~Phe808,疏水性较高的区域主要集中在Met1~Gln18和Thr740~Thr763。
2.4.3跨膜结构及信号肽分析 使用TMHMM在线软件和SignalP在线软件分别对ACE2蛋白质的跨膜结构和信号肽进行分析。跨膜结构分析结果(图7A)显示该鹅ACE2氨基端第1~739氨基酸位于胞外,第740~762氨基酸为跨膜部分,靠近羧基端第763~808氨基酸位于胞内,仅有1次跨膜,符合Ⅰ型跨膜蛋白。信号肽分析结果(图7B)显示图框处C值最大,S值陡峭,Y值最高峰,预测此处第17~18氨基酸之间存在潜在的裂解位点,且该蛋白为分泌蛋白(分泌蛋白由D值来判断,D值是S值和Y值的平均值,该值对区分是否为分泌蛋白具有重要作用,该蛋白D值为0.711,大于0.450,此为分泌蛋白的判断标准),此信号肽区域在第1~17氨基酸之间,这与跨膜区分析预测的结果一致。

A.ACE2蛋白质跨膜结构分析结果;B.ACE2蛋白质信号肽分析结果
2.4.4磷酸化位点分析 使用NetPhos2.0Server对ACE2蛋白质的磷酸化位点进行分析,结果如图8所示。该鹅ACE2蛋白质有较多的苏氨酸Thr(27处)以及丝氨酸Ser(42处)磷酸化位点和相对较少的酪氨酸Tyr(21处)磷酸化位点。
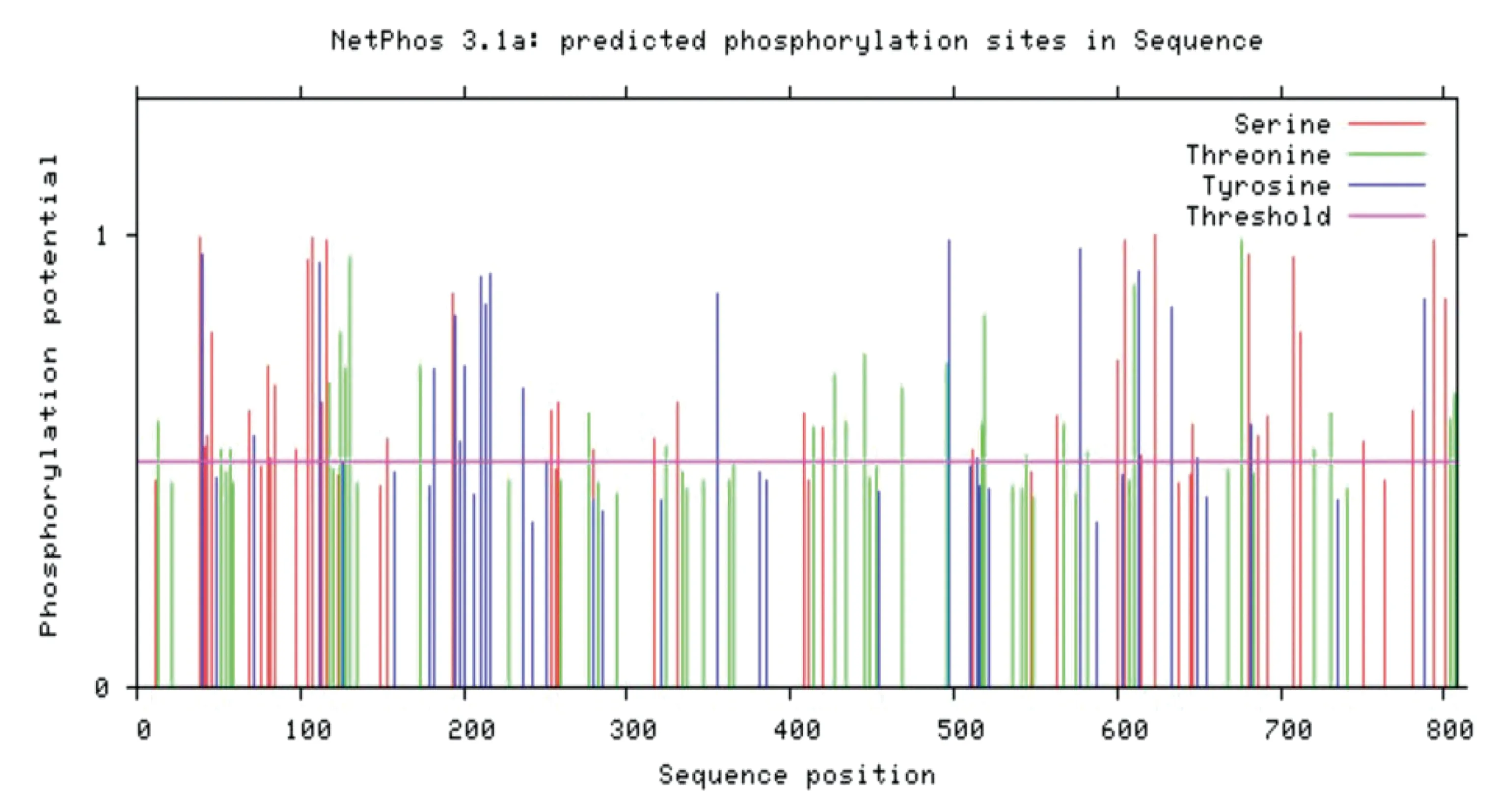
图8 ACE2蛋白质磷酸化位点分析结果
2.5 中国家鹅与人及其他动物的ACE2 S蛋白结合关键氨基酸比较已有报道,ACE2主要与SARS-CoV-2病毒S蛋白结合,与SARS-CoV-2 S蛋白中的受体结合域(RBD)接触的关键氨基酸残基有20个,这些氨基酸残基在不同动物中存在差异,这种差异可能与动物的SARS-CoV-2易感性有关[11]。因此,本试验将所得鹅ACE2中与S蛋白接触的氨基酸序列与SARS-CoV-2易感性较高的人(NM-021804.2)、猫(NM-001039456.1)和易感性较低的犬(NM-001165260.1)以及不易感的鸡(MK560199)、鸭(MK836421)的序列进行比较[13]。结果见表2,鹅与人该段序列有5个氨基酸残基(第330,353,355,357,393位)相同,与猫有6个氨基酸残基(第30,330,353,355,357,393位)相同,与犬有7个氨基酸(第30,41,330,353,355,397,393位)相同,与鸡有3个氨基酸(第42,83,354位)不同,与有鸭5个氨基酸残基(第34,35,42,83,354位)不同。结果显示,鹅与对SARS-CoV-2有易感性的人、猫、犬等动物的ACE2该段序列差异较大,而与对SARS-CoV-2不易感的鸡、鸭等动物的差异较小,提示鹅可能对SARS-CoV-2不易感。

表2 鹅与人及其他动物ACE2中和SARS-CoV-2 RBD接触氨基酸残基比较
3 讨论
2000年,人ACE2完整cDNA首次被克隆,并发现该基因定位于X染色体的p22位置,含有18个外显子,编码805个氨基酸,其编码蛋白为Ⅰ型跨膜蛋白,含有催化胞外区、单个跨膜区和胞内羧基结构域[1,8]。此后,多种动物的ACE2基因序列相继被克隆。但直到2009年,研究人员首次克隆出了野猪ACE2的mRNA[14]。2010年,TSENG等[15]克隆了长白猪的ACE2基因。本实验室杨维维等[16]于2015年克隆出了山羊ACE2完整基因;肖航等[17]于2018年克隆出了断奶仔猪的ACE2全长基因。免疫组化试验等证实ACE2在这些动物体内分布广泛,在多种组织如在心脏、肾脏、睾丸、肺脏、肝脏、肠、脑中均有表达[18]。初步的研究结果证实,广泛存在于动物体内的ACE2与人体中ACE2相同,也能够作为RAS负调节轴的关键酶降解AngⅡ而发挥抗炎、抗损伤等作用[4],对仔猪肠道炎症、氨基酸转运及奶牛乳腺炎症等有一定保护作用[19-21]。
目前,对ACE2的研究主要集中在哺乳动物,而对禽类的研究较少。本实验室纪晓霞等[22]于2020年克隆了白羽肉鸡的ACE2基因序列,该基因全长2 444 bp,编码808个氨基酸;同年李帅等[12]克隆了中国麻鸭的ACE2基因,编码序列长度为2 435 bp,编码805个氨基酸。本试验对鹅的ACE2基因进行了克隆,通过分析结果显示该鹅ACE2氨基酸序列与中国麻鸭ACE2氨基酸序列同源性为93.1%,并处在同一进化分支上。对该鹅ACE2编码蛋白质进行生物信息学解析,发现其ACE2蛋白含有较多的亲水区域,总体表现为亲水性。含有1个跨膜区,属于Ⅰ型跨膜蛋白。与上面在白羽肉鸡和中国麻鸭上的研究结果相似。
2020年初,新型冠状肺炎在全球暴发,ACE2被证实是新型冠状肺炎病原SARS-CoV-2的功能性受体,人、蝙蝠、穿山甲、雪貂、猫等哺乳动物均通过ACE2感染病毒[13]。目前,动物冠状病毒与动物体内ACE2及与人SARS-CoV-2之间的关系的研究缺少确切结果。据报道,鸡、鸭对SARS-CoV-2不易感,序列分析发现鸡、鸭ACE2中与S蛋白接触的关键氨基酸残基与人差异较大[11,23]。本试验也通过比较鹅与人、猫、犬、鸡、鸭的ACE2 S蛋白结合关键氨基酸序列,发现鹅该序列与鸡、鸭等禽类相似,但与人、猫、犬存在较大差异。
YAN等[10]揭示了人ACE2与SARS-CoV-2结合的结构基础,其研究显示SARS-CoV-2 RBD的Q498、T500和N501与ACE2的Y41、Q42、K353和R357形成氢键网络,RBD的K417和Y453分别与ACE2的D30和H34相互作用,RBD的Q474与ACE2的Q24形成氢键,RBD的F486与ACE2的M82通过范德华力相互作用。在本研究中同样发现鹅ACE2第24,27,28,30,31,34,37,41,42,79,82,83位氨基酸在对SARS-CoV-2较敏感的动物中较保守,而禽类等非易感动物的ACE2中这些位置的氨基酸与易感动物均不同,这也与 ALEXANDER等[24]研究结果相符合,其提示这些位置的氨基酸残基可能有利于ACE2与SARS-CoV-2紧密结合,而鹅等家禽ACE2不利于与SARS-CoV-2结合而不易感。但需要注意的是,虽然鹅可能对SARS-CoV-2不易感,但鹅可以感染Ⅲ型冠状病毒[25]。目前,尚未见有报道说明鹅感染冠状病毒与ACE2相关,但鹅冠状病毒可以感染肾脏。本试验结果表明,肾脏ACE2表达量较高,提示鹅感染冠状病毒可能与ACE2存在关联,这方面的研究有待开展。
总之,本试验成功克隆了中国家鹅ACE2的编码区全基因序列,获得序列已上传GenBank。也对其编码蛋白进行了生物信息学预测与分析,并在氨基酸序列层面对鹅是否易感SARS-CoV-2给予了初步分析,这些结果将为鹅ACE2相关方面的研究提供基础和资料。
猜你喜欢
环球时报(2022-09-20)2022-09-20
中国畜牧杂志(2022年6期)2022-06-13
中国典型病例大全(2022年7期)2022-04-22
中国食品(2020年13期)2020-07-29
恋爱婚姻家庭·养生版(2018年3期)2018-03-24
计算机应用(2016年10期)2017-05-12
小资CHIC!ELEGANCE(2015年14期)2015-09-23
小资CHIC!ELEGANCE(2015年15期)2015-09-01
时代英语·高二(2015年2期)2015-05-18
中国计算机报(2009年27期)2009-04-27
