
猪传染性胃肠炎病毒诱导PK-15细胞自噬及其机理
2022-10-24 02:46袁广富张增贤赵云环范京惠左玉柱
中国兽医学报 2022年7期
刘 莹,袁广富,张增贤,张 毅,王 晶,郭 禹,赵云环,范京惠 *,左玉柱*
(1.河北农业大学 动物医学院,河北 保定 071001;2.宁晋县农业农村局,河北 宁晋 055550 3.邢台市动物疫病预防控制中心,河北 邢台 054000)
猪传染性胃肠炎病毒(transmissible gastroenteritis virus,TGEV)为冠状病毒科冠状病毒属成员,是一种急性、高度接触性的肠道冠状病毒,在世界范围内给养猪业造成了重大的经济损失。由于TGEV对高温更敏感,因此在1-2月份为该病的发病高峰,其中2周龄内仔猪的感染率和病死率高,最高时可达100%[1-2]。1946年,TGE首次在美国被报道,随后许多国家相继检测出TGEV的存在并流行。1956年,我国广东首次发现该病并报道,随后在全国大部分地区均发现该病的存在,给养猪业造成了严重的经济损失[1,3]。
自噬(autophagy)(来自希腊语,即“自己吃自己”)是指任何细胞降解途径,包括将细胞质输送到溶酶体,已经确定了3种形式即伴侣介导的自噬、微自噬和巨自噬——它们是生理功能和向溶酶体输送的方式不尽相同[4]。最典型的自噬的核心机制包括以下步骤:自噬体(autophagosomes,AP)在细胞质中完成,AP和溶酶体在微管上运输,AP与溶酶体对接融合,形成自噬溶酶体(AL),自噬降解循环,最终形成自噬溶酶体重组(ALR)循环。典型的自噬通过Ⅲ类磷酸肌醇3-激酶(PI3K)复合物(PI3KC3)/雷帕霉素复合物1(mTORC1)信号轴的机制靶点发生[5-7],该靶点通过磷酸化和抑制丝氨酸/苏氨酸蛋白激酶、UNC51样激酶1(ULK1),直接调节自噬活性,参与自噬起始的第一个蛋白质复合物的一部分[8]。
自噬不仅仅是一种自我降解的细胞生理途径,同时也是一种保护宿主免受病毒感染的防御机制[9]。它可以通过将病毒运送到溶酶体室,抵御入侵病毒[10]。而另一方面,病毒也有许多机制来阻止自噬,甚至操纵自噬。自噬可以通过多种方式促进病毒的复制,包括协助病毒的对机体的侵袭、排出及传入病毒RNA的翻译。病毒也会利用自噬作为复制平台来增强其复制[11-12]。
猪TGEV是α冠状病毒属成员,在过去几年中,对于Togaviridae、Picornaviridae等成员的研究数量迅速增加[13-14]。对于SARS-CoV和IBV的进一步研究也证明了存在自噬体蛋白标记物LC3或自噬样过程。然而,迄今为止所开展的工作主要集中在Betacoronavirus以及Gammacoronavirus[15]。自噬途径在α冠状病毒(如TGEV)复制中的作用仍有待研究。本试验拟探讨TGEV的感染是如何影响自噬以及自噬是如何影响TGEV的感染,以及两者之间的关系及作用机制,为TGEV流行毒株感染致病的分子机制及生物制品的研发提供理论依据。
1 材料与方法
1.1 细胞、病毒株及菌株猪肾细胞(PK-15),由河北农业大学动物医学院预防兽医系保存;TGEV,由本研究在临床样品中分离获得;大肠杆菌DH5α感受态细胞,为本实验室制作并保存。
1.2 主要试剂与仪器DMEM培养基、胎牛血清购自Gibco公司;β-actin购自美国Abclonal公司;3-Methyladenine购自美国Cayman公司;雷帕霉素购自美国BioVision公司;FITC-羊抗兔IgG、高效RIPA裂解液购自北京索莱宝科技有限公司;ECL Reagent购自RedPartyTech公司;2×SYBR Green qPCR Master Mix购自苏州宇恒生物科技有限公司;凝胶成像仪购自美国ProteinSimple公司;荧光倒置显微镜购自OPTEC公司;半干转印槽转膜仪购自BIO-RAD公司;酶标仪购自Thermo Scientific 公司。
1.3 PK-15细胞的培养将猪肾细胞(PK-15细胞)接种于含有10%胎牛血清、1%青霉素-链霉素贮存液的DMEM营养液的T25细胞培养瓶中,置于37℃、5%CO2的培养箱中培养。当细胞培养瓶中的PK-15细胞生长形态良好且细胞生长至70%~80%进行传代培养。
1.4 TGEV感染对自噬相关蛋白表达影响为从分子水平上研究TGEV感染PK-15细胞对细胞自噬的影响,取状态良好的PK-15细胞接种于6孔板中,当细胞生长至80%左右,病毒感染量分别为0.000,0.005,0.050,0.500 MOI接种于PK-15细胞中。将6孔培养板放置37℃、5%CO2细胞培养箱中培养1 h 后,每孔加入350 μL维持液继续培养。在感染不同时间(4,8,12 h)进行细胞样品的收取,通过Western blot免疫印迹分析,利用高效裂解液(RIPA∶PMSF=1 000∶1)对各组细胞沉淀样品充分裂解,对蛋白样品进行SDS-PAGE电泳,用半干转印法将蛋白转移至NC膜上,ECL高效发光液避光孵育并在成像仪中进行曝光拍照保存。
1.5 紫外线灭活TGEV感染对LC3蛋白表达的影响为探明TGEV感染后LC3-Ⅱ蛋白的表达是否依赖病毒的增殖,本试验将紫外灭活6 h的TGEV接种于PK-15细胞,同时设置阳性对照组和阴性对照组。通过间接免疫荧光,在接毒 24 h 之后PBS洗涤 3 次,用4%多聚甲醛固定后用PBS清洗细胞,向细胞中加入0.2%Triton X-100通透10 min后,再次用PBS清洗,5%BSA在37℃孵育1 h,PBS清洗4次后,将1∶100倍稀释的兔抗TGEV全病毒多抗加入细胞中,室温孵育2 h,PBS清洗 3 次,加入FITC-羊抗兔IgG,避光反应 1 h后,放置于荧光倒置显微镜下观察并拍照保存。利用SDS-PAGE电泳分析各组的自噬相关蛋白表达的变化情况。
1.6 药物处理对TGEV增殖的影响为研究细胞自噬对TGEV的增殖的影响,选用自噬激活剂雷帕霉素(25 nmol/L)和自噬抑制剂3-MA对PK-15细胞进行预处理12 h后感染病毒量为 1 MOI的TGEV HB-FP株,其培养细胞所用的维持液也含有相同浓度的雷帕霉素。分别收集雷帕霉素未处理组、雷帕霉素预处理组和空白对照组感染病毒8,12,24,36,48,60 h时的细胞病毒培养物,对其TGEV核酸量、病毒滴度及细胞自噬程度进行测定。
1.7 TGEV感染PK-15细胞对自噬关键基因的影响自噬关键基因ATG-5、Beclin-1在自噬发生与调控过程中起到了非常重要的作用,本研究以1 MOI 的TGEV病毒量接种于生长良好的细胞中,分别收集8,12,24,36,48,60 h病毒液,通过荧光定量检测方法对不同时间的自噬关键基因进行检测。根据GenBank中登录的ATG-5及Beclin-1的序列分别设计2对荧光定量PCR引物,ATG-5-F:5′-CATCGCGCAACCAACAGATTGA-3′,ATG-5-R:5′-TTCCTGTTAGGCTTGCAGCAGTG-3′;Beclin-1-F:5′-GCATGAAGATGACAGCGAACAGC-3′,Beclin-1-R:5′-AGCTTCC TCCTGATCCAACCTCTC-3′。引物均由生工生物工程(上海)有限公司合成。
2 结果
2.1 TGEV HB-FP株感染引起自噬体标志蛋白LC3-Ⅱ表达的变化通过研究发现,分别在病毒的感染量相同和感染时间相同的情况下,与对照细胞组相比,LC3-Ⅱ蛋白的表达量均逐渐增加。通过Imag J软件进行条带灰度值分析表明LC3-Ⅱ/β-actin的比值明显增加。结果表明,TGEV HB-FP株的感染对PK-15细胞的自噬起到了诱导作用(图1)。
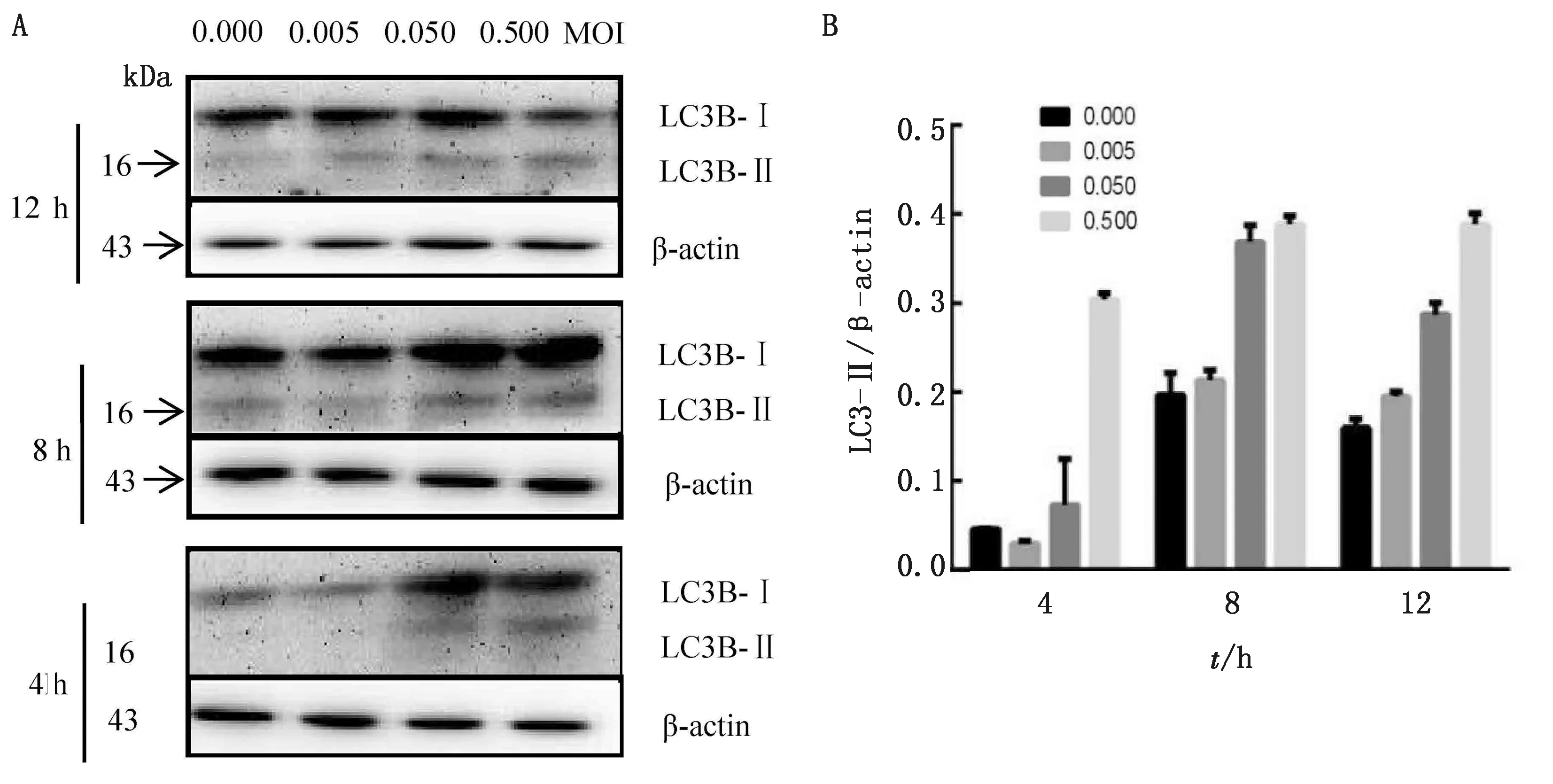
A.蛋白免疫印迹检测TGEV对自噬相关蛋白表达的影响;B.LC3-Ⅱ/β-actin灰度值比值
2.2 紫外线灭活TGEV感染对LC3蛋白表达的影响免疫荧光试验得出紫外线灭活组未检测到TGEV抗原的产生,证明TGEV被彻底灭活。通过Western blot对LC3蛋白表达量的变化进行检测,结果显示正常病毒组的LC3-Ⅱ/β-actin的表达量显著高于紫外灭活组(图2)。
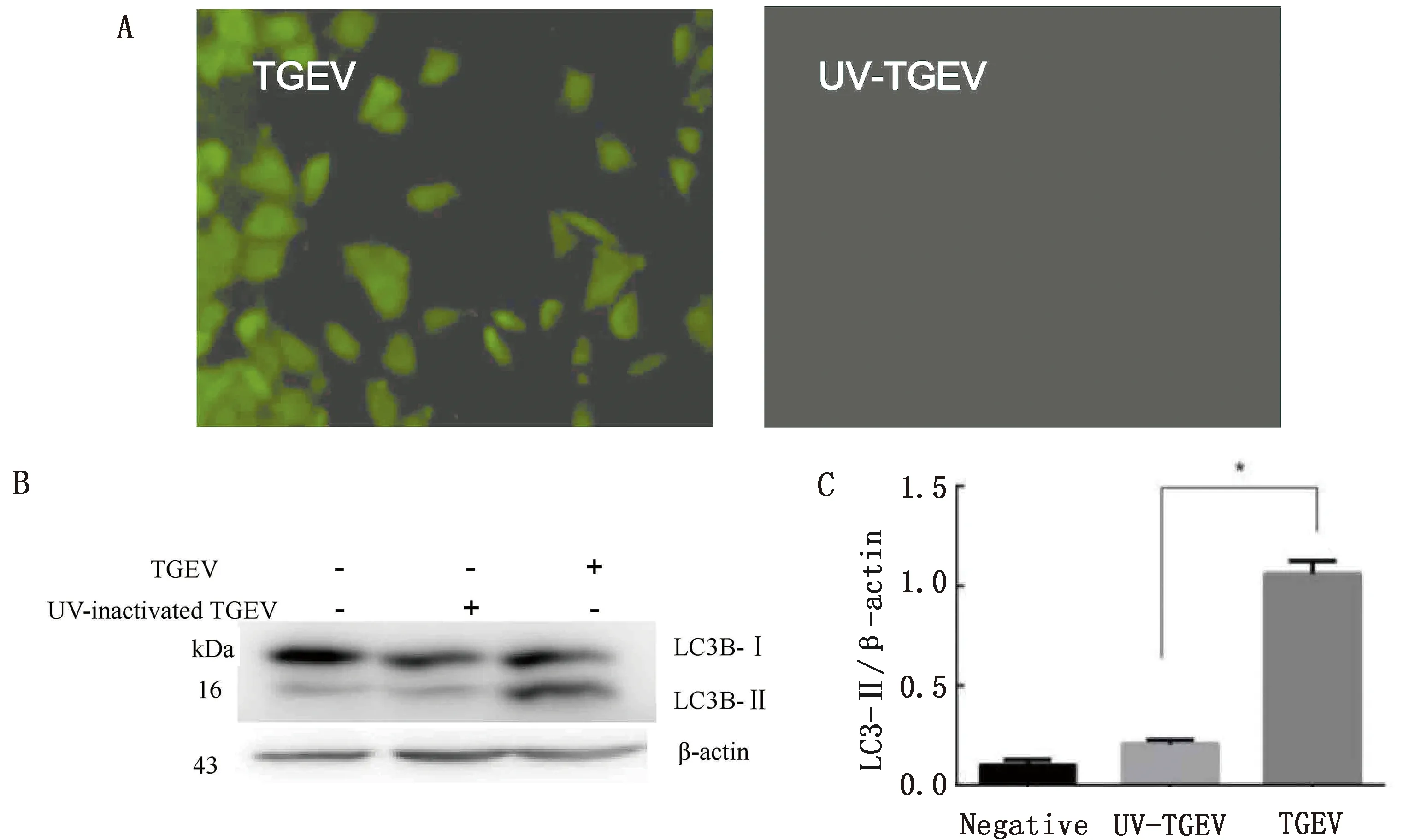
A.IFA检测灭活的TGEV是否有抗原产生;B.蛋白免疫印迹检测灭活TGEV对自噬相关蛋白表达的影响;C.LC3-Ⅱ/β-actin灰度值比值
2.3 自噬激活剂雷帕霉素增强了TGEV的增殖结果见图3。空白对照组未检测出TGEV核酸;在不同时间段,雷帕霉素预处理组TGEV核酸量及病毒滴度均高于雷帕霉素未处理组,且随着时间的增加而增加。感染TGEV后雷帕霉素处理的PK-15细胞,从病毒感染后的 24 h开始,其TGEV核酸量与未处理组呈现极显著差异(P<0.01),其病毒滴度与未处理组相比也均呈现极显著差异(P<0.01)。
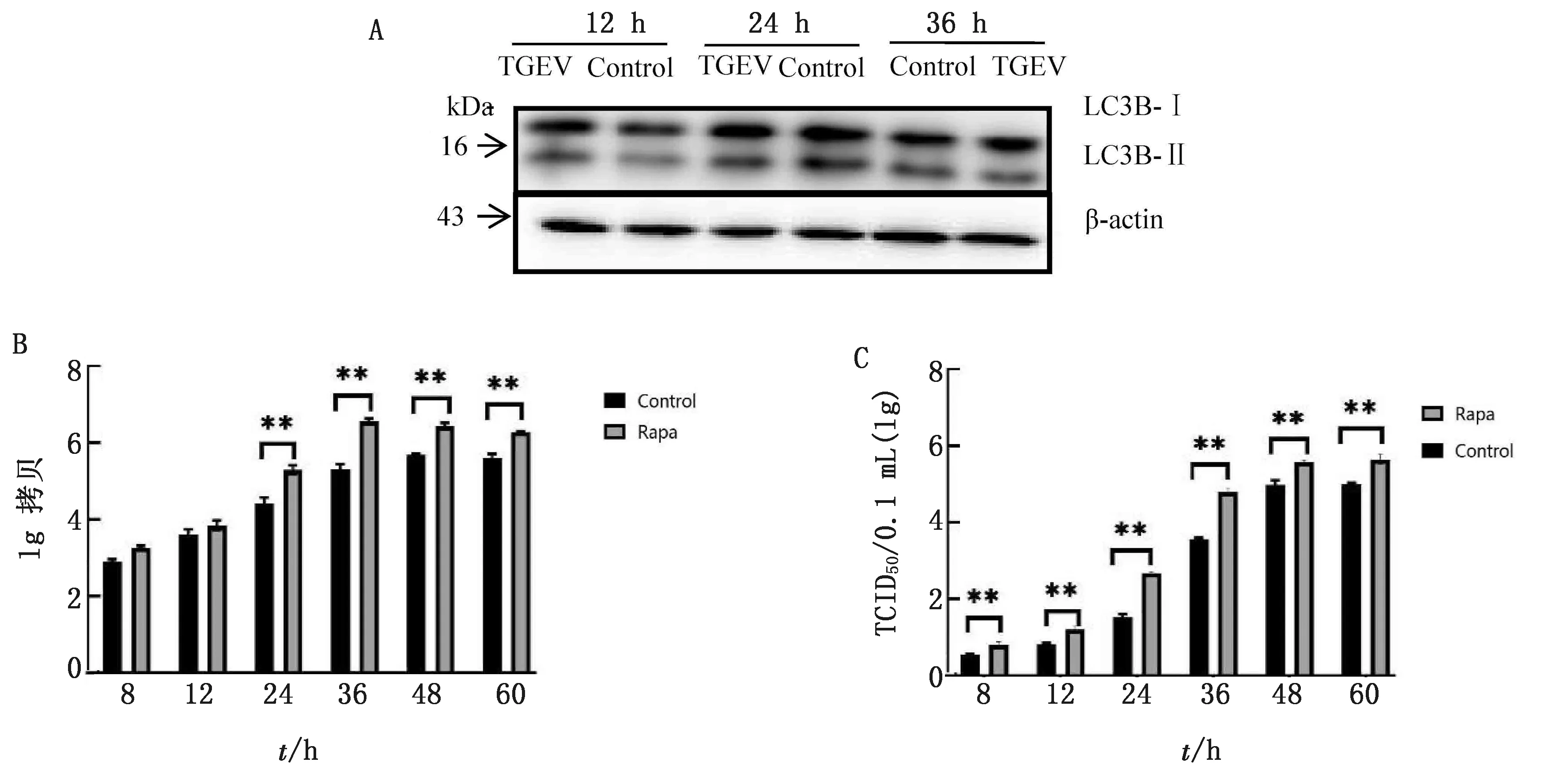
A.Western blot检测自噬相关蛋白表达;B.qPCR检测病毒核酸拷贝数;C.TGEV病毒滴度变化。*示P<0.05,**示P<0.01。下同
2.4 自噬抑制剂3-MA抑制了TGEV的增殖通过Western blot检测分析发现,3-MA预处理后LC3-Ⅱ蛋白的表达水平明显降低。RT-qPCR、ImagJ软件分析结果显示,在不同时间段,3-MA预处理组细胞的病毒核酸含量及病毒滴度均低于未处理组,其病毒核酸含量在TGEV感染12~48 h时,与未处理组相比呈现极显著性差异(P<0.01),而在感染60 h时差异降低,呈显著性差异(P<0.05);其病毒滴度在TGEV感染后12 h时与未处理组相比呈现显著性差异(P<0.05),24 h后呈现极显著差异(P<0.01)(图4)。综上所述,自噬抑制剂3-MA对TGEV的增殖起到抑制作用。
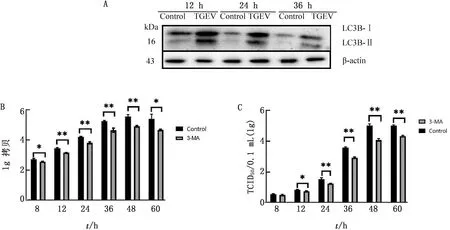
A.Western blot检测自噬相关蛋白表达;B.qPCR检测病毒核酸拷贝数;C.TGEV病毒滴度变化
2.5 TGEV感染PK-15细胞对自噬关键基因的影响结果见图5。ATG-5与Beclin-1基因在TGEV感染细胞后,其在不同感染时间的表达量均高于空白对照组。ATG-5基因在8~24 h时表达量略高,但差异不明显;在 36~60 h 时 2 组的表达量均持续上升,呈现极显著性差异(P<0.01)。Beclin-1基因在 8~36 h 时,其感染组表达量呈线性增长,在 36~60 h 时其表达量趋于平稳,但其表达量与空白对照组均呈现极显著性差异(P<0.01)。结果表明,TGEV感染细胞后可对自噬关键基因的表达起促进作用。
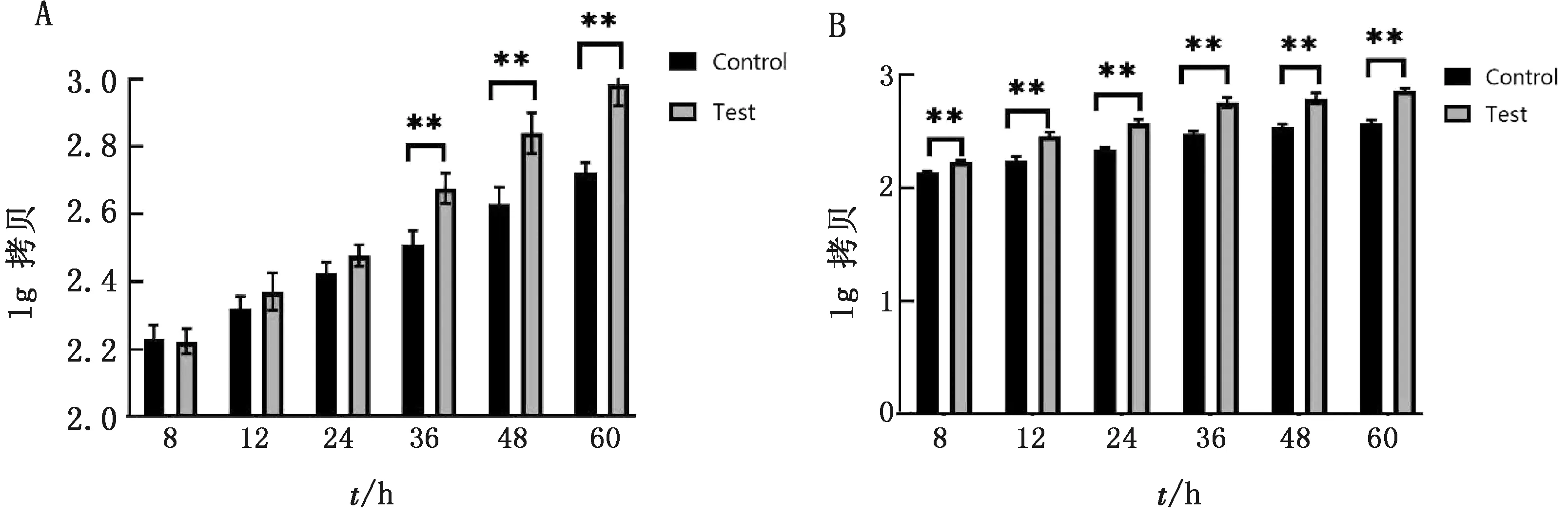
A.ATG-5;B.Beclin-1
3 讨论
自噬是进化上保守的细胞降解过程,其中细胞成分经溶酶体途径被消化[16]。在病毒感染的期间,自噬可以作为宿主监视机制,其中病毒抗原被递送到富含免疫传感器的内体和溶酶体中,这些免疫传感器可以在刺激时激活自噬[17]。反之,病毒也采用了几种方法来逃避这种抗病毒机制。虽然许多病毒阻断了自噬途径中的不同点,但有些病毒会为了自身的利益而主动颠覆自噬[18]。异种自噬是选择性自噬的一种形式,特异性识别细胞内微生物,并将这些病原体物理定位于自噬途径进行降解[19]。因此,一些病原体进化出多种策略来抑制自噬,而另一些则利用自噬体膜和自噬机制来增强其复制。例如:细胞自噬能够抑制猪伪狂犬病病毒、猪塞内卡病毒等的增殖,而能够促进猪瘟病毒、猪流行性腹泻病毒、流感病毒等的增殖。而目前国内关于细胞自噬与TGEV相互作用的文献报道相对较少,因此,本试验对TGEV HB-FP分离株与宿主细胞PK-15细胞自噬的相互作用与影响进行研究与分析。
最初,我们通过利用TGEV HB-FP株的感染造成自噬体标志蛋白LC3-Ⅱ的表达变化,从分子水平上探究TGEV感染PK-15细胞对细胞自噬的影响,分别在感染后4,8,12 h时收集细胞培养物,检测LC3蛋白的表达及转化水平,并利用紫外线灭活使TGEV失去感染力来探明TGEV感染细胞后引起的细胞自噬的增加是否依赖于病毒的复制,结果显示正常病毒感染组随病毒感染量的升高,LC3-Ⅱ/β-actin表达量比值显著提高,而紫外灭活组的LC3-Ⅱ/β-actin表达量虽也略有升高,但是其诱导细胞自噬的能力与正常病毒感染组相比明显下降。本试验结果表明,失活的病毒未完全失去诱导细胞自噬的能力,但病毒的复制可促进细胞自噬的程度。
既然,TGEV的复制可以促进细胞自噬的程度,那么自噬的发生是否有利于病毒的增殖呢?我们通过选用合适浓度细胞自噬的促进剂雷帕霉素及抑制剂3-MA处理细胞,经自噬促进剂雷帕霉素处理的细胞,在感染TGEV HB-FP株后与对照组相比,TGEV的增殖为正向调控。但在动物体内,自噬与TGEV病毒滴度之间的相互作用还有待进一步研究。
为进一步探究在自噬发生和调控过程中自噬关键基因与TGEV之间的作用关系,本试验通过监测Beclin-1、ATG-5基因mRNA转录量在细胞感染TGEV后的表达变化来揭示病毒增殖对自噬关键基因的影响。结果显示,对照组的2种自噬关键基因mRNA转录量变化并不显著,在36 h后mRNA转录量开始有所升高,出现此现象的原因可能是由于细胞培养时间过长,其营养液中的营养物质减少而导致细胞诱导自噬引起;而试验组细胞感染TGEV后自噬关键基因的mRNA转录量显著上升,结合病毒的增殖曲线也印证了TGEV感染增殖与细胞自噬的正向调控,在24~48 h时病毒属于一个快速增殖阶段,其自噬关键基因的mRNA转录量也处于较高水平。
对于TGEV来说,自噬与病毒之间是一种相互促进的关系,但是自噬作为一种机体的保护防御机制,为何会被病毒利用而有利于病毒的增殖,病毒是如何进行免疫逃逸的机制还需要进一步研究。
猜你喜欢
中国现代医生(2022年21期)2022-08-22
大众科学(2022年7期)2022-08-19
中国慈善家(2022年3期)2022-06-14
现代苏州(2022年9期)2022-05-26
广东教育·职教版(2021年3期)2021-04-20
医学前沿(2021年18期)2021-04-14
学校教育研究(2018年27期)2018-05-14
婚育与健康(2018年1期)2018-04-08
分析化学(2017年12期)2017-12-25
科学中国人(2016年1期)2016-01-13
