
LINC00710靶向miR-367/PTEN轴抑制肺癌A549细胞的增殖、迁移和侵袭
2022-10-21 04:28莫静欣
广西医科大学学报 2022年9期
莫静欣,冯 莉,杨 力
(十堰市人民医院呼吸内科重症监护室,十堰 442000)
肺癌已成为中国癌症死亡的主要原因。由于缺乏早期诊断标志物,超过50%肺癌患者确诊时已经处于晚期,使患者错过最佳治疗时机,导致5年生存率未超过15%[1-2]。因此,探讨肺癌的分子机制对寻找肺癌诊断和治疗靶点有重要的意义。长链非编码RNA(lncRNAs)是长度超过200 nt 的非编码RNA转录本,其通过直接或间接与miRNA、蛋白质相互作用等参与肿瘤的发展[3-4]。Zhao 等[5]采用lncRNA芯片技术和荧光定量PCR分析发现肺腺癌组织中LINC00710表达显著降低,但LINC00710在肺癌细胞中的具体生物学作用尚未可知。微小RNA(miR)-367是一种肿瘤相关因子,miR-367高表达与肺癌患者较差的预后、肿瘤大小、肿瘤分期、转移相关,抑制miR-367能促进肺癌细胞增殖和侵袭,并诱导细胞凋亡,进而促进肺癌发生和发展[6]。第10 号染色体缺失的磷酸酶和张力蛋白同源等位基因(PTEN)是一种抑癌基因,PTEN低表达对肺癌的发生、浸润及转移具有促进作用。生物信息学分析发现,LINC00710 与miR-367、miR-367 与PTEN 之间存在互补结合位点,但LINC00710 是否靶向miR-367/PTEN 轴参与肺癌进展尚未可知。本研究以miR-367、PTEN 为关键点,旨在探讨LINC00710 对肺癌细胞恶性行为的影响和可能的分子机制。
1 材料与方法
1.1 实验组织
收集2017 年12 月至2018 年12 月于本院确诊为肺癌的33 例患者,其中男15 例,女18 例;所有患者术前均未接受化疗、放疗或其它治疗;TNM 分期:Ⅰ~Ⅱ期17 例,Ⅲ期16 例;淋巴结转移20 例,未转移13 例。将切除的肺癌患者的癌组织及癌旁组织分割成小块,生理盐水漂洗去除血渍和污物,置于液氮中冷冻,置于-80 ℃冰箱中保存。本研究符合伦理委员会的规定,并在患者知情同意的情况下进行。
1.2 细胞和试剂
肺癌A549 细胞购自中国科学院上海生物科学研究所。青/链霉素双抗、DMEM培养基、胎牛血清购于美国Gibco 公司;TRIzol 试剂、Opti-MEM 培养基、Lipofectamine 2000 购于美国Invitrogen 公司;LINC00710 过表达质粒(pcDNA3.1-LINC00710)、空载体质粒(pcDNA3.1)、miR-367 模拟物(miR-367 mimic)、双荧光素酶报告基因载体购于上海吐露港生物技术公司;miRNA荧光定量PCR试剂盒和逆转录试剂盒购于北京百奥莱博生物公司;Prime-ScriptTMRT 试剂盒、2×SYBR Premix ExTaq 试剂盒购于大连TAKARA 生物;细胞增殖检测试剂盒(CCK-8)购于上海晶抗生物公司;基质胶、Transwell培养板购于美国BD 公司;兔源Ki67 抗体、羊抗兔IgG 二抗、兔源PTEN 抗体、兔源上皮细胞钙粘蛋白(E-cadherin)抗体和兔源神经钙黏素(N-cadherin)抗体、兔源磷酸甘油醛脱氢酶(GAPDH)抗体购于上海赛信通公司。
1.3 方法
1.3.1 细胞培养 A549 细胞培养于DMEM 中(含10%胎牛血清、100 U/mL 青霉素与100 μg/mL 链霉素)。当细胞80%汇合时进行0.25%胰蛋白酶消化,1∶3 比例传代,每3~4 d 传代一次。培养箱环境为37 ℃、5%CO2。
1.3.2 RT-qPCR检测LINC00710和miR-367表达 用TRIzol 试剂从肺癌组织和癌旁组织中提取总RNA。为检测LINC00710和miR-367表达,分别用PrimeScriptTM和miRNA 逆转录试剂盒合成cDNA,再分别用2×SYBR Premix ExTaq荧光定量PCR试剂盒进行RT-qPCR。LINC00710 上游引物:5’-AGCACCTGTTCCCCTCTACT-3’,下游引物:5’-CCAGGGTGAATGGAAACCCA-3’;miR-367 上游引物:5’-TCCACCACCCAGTTGCTGTA-3’,下游引物:5’-CGAGCAATTGCACTTTAGCAAT-3’;GAPDH 上游引物:5’-CCCACATGGCCTCCAAGGAGTA-3’,下游引物:5’-GTGTACATGGCAACTGTGA-GGAGG-3’;U6 上游引物:5’-GGTCGGGCAGGA-AAGAGGGC-3’,下游引物:5’-GCTAATCTTCTCTGTATCGTTCC-3’。采用2-ΔΔCT方法计算LINC00710(GAPDH 为内参)和miR-367(U6 为内参)的相对表达水平。
1.3.3 转染和实验分组 将A549 细胞以2×105个/孔接种到6 孔板中,当细胞50%汇合时进行细胞转染。利用脂质体转染试剂Lipofectamine 2000、Opti-MEM 培养基 分别将pcDNA3.1、pcDNA3.1-LINC00710、miR-367 mimic、miR-367 mimic+pcDNA3.1-LINC00710 转染A549 细胞,依次记为pcDNA3.1组、pcDNA3.1-LINC00710组、miR-367 mimic组、miR-367 mimic+pcDNA3.1-LINC00710 组。另设置对照组,不进行转染。6 h后更换为新鲜含血清DMEM 培养基,转染48 h 后收集细胞,用RT-qPCR法检测转染效果。
1.3.4 CCK-8 法检测细胞活力 转染细胞以5×103个/孔的密度接种于96 孔板中,培养24 h,每孔加入90 μL 的DMEM 和10 μL 的CCK-8。采用酶标仪检测孵育2 h后各孔细胞在450 nm波长处吸光度值。
1.3.5 集落形成实验检测克隆形成数 取0.25%胰蛋白酶、0.02%EDTA 溶液消化各组A549 细胞。按照每孔600 个细胞接种60 mm 培养板,每2 d 更换1 次培养基,当出现肉眼可见的克隆时停止培养。4%多聚甲醛固定细胞集落,0.1%结晶紫对细胞集落染色。随后显微镜对菌落进行计数。
1.3.6 Transwell 实验检测细胞迁移和侵袭 向24孔板下室中加600 μL含10%血清的细胞培养基,将包被(侵袭检测)或未包被(迁移检测)基质胶的Transwell上室中加入200 μL。培养24 h后,去除未穿膜细胞,用4%多聚甲醛固定侵袭细胞,PBS 冲洗3次,0.1%结晶紫染色10 min,PBS冲洗3次,随机选择5个视野进行细胞计数、观察和拍照。
1.3.7 Western blotting 检测PTEN、Ki67、E-cadherin和N-cadherin 蛋白表达 预冷RIPA 缓冲液裂解细胞,离心收集细胞上清液,BCA 法进行蛋白定量。按照30 μg 每孔上样进行电泳,转移至PVDF 膜上,4 ℃下用5%脱脂牛奶封闭1 h,用1∶1 000 稀释的PTEN、Ki67、E-cadherin、N-cadherin、GAPDH 一抗溶液室温孵育2 h,加入1∶1 000 稀释的二抗溶液孵育,加入化学发光试剂暗室显色后,用Image J 软件定量分析目的条带灰度值。结果以目的蛋白和内参GAPDH灰度值的比值表示。
1.3.8 双荧光素酶报告基因实验 将含有miR-367预测结合位点的LINC00710(或PTEN-3’UTR)野生型序列或不含预测结合位点的LINC00710(或PTEN-3’UTR)突变型序列分别克隆到pmirGLO载体,构建重组载体WT-LINC00710、WT-PTEN、MUTLINC00710、MUT-PTEN。将A549 细胞以1×104个/孔的密度接种于24孔板中,当细胞汇合度达到50%时,使用Lipofectamine 2000试剂分别将重组载体与miR-367 mimic 或control 共转染到A549 细胞中。检测培养48 h后细胞相对荧光素酶活性。
1.4 统计学方法
实验独立重复3次,采用SPSS 20.0软件进行数据分析,符合正态分布的计量资料用均数±标准差()表示,两组间比较用t检验,多组间比较用单因素方差分析,进一步两两比较采用SNK-q检验。以P<0.05为差异有统计学意义。
2 结果
2.1 LINC00710、miR-367在肺癌中的表达
与癌旁组织比较,肺癌组织中LINC00710的表达水平降低,miR-367 的表达水平升高(均P<0.05),见表1。
表1 LINC00710、miR-367在肺癌组织中的表达
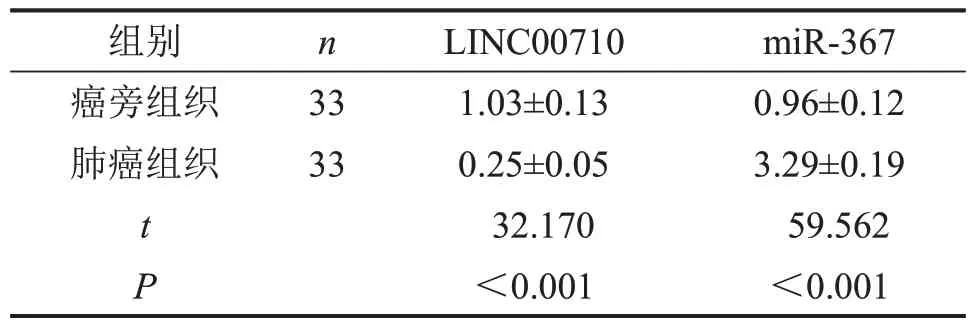
表1 LINC00710、miR-367在肺癌组织中的表达
2.2 各个处理组对A549增殖迁移、侵袭的影响
与对照组、pcDNA3.1 组比较,pcDNA3.1-LINC 00710 组A549 细胞增殖活性降低,克隆形成数、迁移及侵袭细胞数减少(均P<0.05);与对照组比较,miR-367 mimic 组A549 细胞增殖活性升高,克隆形成数、迁移和侵袭细胞数增多(均P<0.05);与pcDNA3.1-LINC00710 组比较,miR-367 mimic+pcDNA3.1-LINC00710 组A549 细胞增殖活性升高,克隆形成数、迁移和侵袭细胞数增多(P<0.05),见表2、图1。

图1 LINC00710、miR-367对A549细胞增殖、迁移和侵袭的影响
2.3 LINC00710 靶 向miR-367 及miR-367 靶 向PTEN
DIANA Tools 数据库预测发现LINC00710 与miR-367、miR-367 与PTEN 之间存在互补结合位点,见图2。双荧光素酶报告实验显示,与对照组和WT-LINC00710(或WT-PTEN)共转染组比较,miR-367 mimic 和WT-LINC00710(或WT-PTEN)共转染组A549 细胞的相对荧光素酶活性降低(P<0.05);与对照组和MUT-LINC00710(或MUT-PTEN)共转染组比较,miR-367 mimic 和MUT-LINC00710(或MUT-PTEN)共转染组A549细胞的相对荧光素酶活性无显著变化(P>0.05),见图3。

图2 miR-367与LINC00710、PTEN的靶向关系
表2 LINC00710、miR-367对A549细胞增殖、迁移和侵袭的影响,n=3

表2 LINC00710、miR-367对A549细胞增殖、迁移和侵袭的影响,n=3
与对照组比较,*P<0.05;与pcDNA3.1组比较,#P<0.05;与pcDNA3.1-LINC00710组比较,&P<0.05。
2.4 miR-367 能减轻LINC00710 对A549 中Ki67、E-cadherin、N-cadherin蛋白表达的影响
与对照组、pcDNA3.1 组比较,pcDNA3.1-LINC00710 组A549 细胞miR-367 表达、Ki67 和Ncadherin 蛋白表达降低,PTEN 和E-cadherin 蛋白表达升高(P<0.05);与对照组比较,miR-367 mimic组A549 细胞miR-367 表达、Ki67 和N-cadherin 蛋白表达升高,PTEN和E-cadherin蛋白表达水平降低(P<0.05);与pcDNA3.1-LINC00710 组比较,miR-367 mimic+pcDNA3.1-LINC00710 组A549 细胞miR-367 表达、Ki67 和N-cadherin 蛋白表达升高,PTEN和E-cadherin蛋白表达降低(P<0.05),见表3和图4。
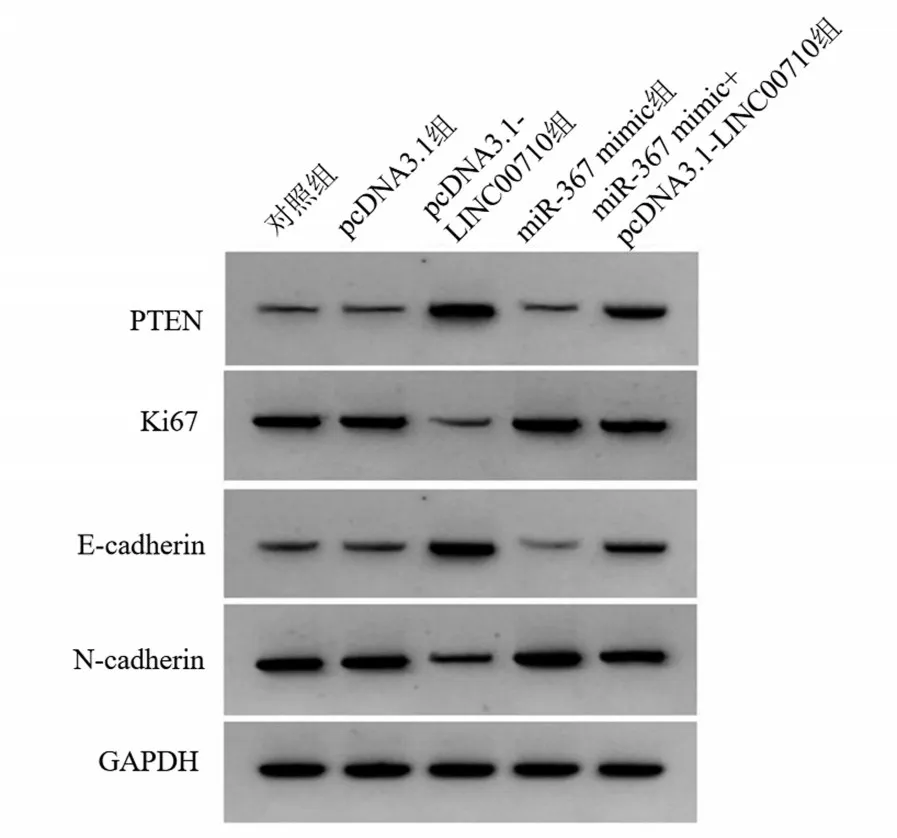
图4 PTEN、Ki67、E-cadherin、N-cadherin蛋白的表达
表3 miR-367能减轻LINC00710对A549中PTEN、Ki67、E-cadherin、N-cadherin蛋白表达的影响,n=3

表3 miR-367能减轻LINC00710对A549中PTEN、Ki67、E-cadherin、N-cadherin蛋白表达的影响,n=3
与对照组比较,*P<0.05;与pcDNA3.1组比较,#P<0.05;与pcDNA3.1-LINC00710组比较,&P<0.05。
3 讨论
目前肺癌的诊断和治疗方法还很有限,其治疗效果欠佳[8]。已发现许多lncRNA 与肺癌进展相关。如lncRNA成神经细胞瘤相关转录物1(NBAT-1)在肺癌组织标本中表达较癌旁组织显著降低,NBAT-1 低表达与患者肿瘤大小、淋巴结转移情况显著相关,过表达NBAT-1可抑制A549细胞增殖和细胞周期,诱导细胞凋亡[9]。肺癌细胞株中lncRNA X 染色体失活基因(XIST)的表达显著高于正常人支气管上皮细胞,抑制XIST表达可降低细胞增殖、迁移和侵袭能力,抑制A549 异种移植小鼠模型的肿瘤生长[10]。但lncRNA 在肺癌中的潜在分子机制仍需进一步研究。本研究结果发现,肺癌组织中LINC00710 表达较癌旁组织显著降低,与赵兵等[11]研究结论相一致。Ki67 是一种标记细胞增殖状态的核抗原,其表达水平可准确反映细胞增殖活性[12]。E-cadherin 表达增加可维持细胞形态和黏附功能,而N-cadherin 表达增加则促进细胞骨架发生改变,降低细胞黏附能力,使其易于脱离原发灶,向周围组织侵袭转移[13]。本研究结果显示,过表达LINC00710 可显著降低A549 细胞存活率、克隆形成、迁移和侵袭能力,降低Ki67和E-cadherin蛋白表达水平,提高N-cadherin表达水平,表明LINC00710对A549细胞增殖、迁移和侵袭具有显著抑制作用。
miRNA 表达与肿瘤发生、耐药、肿瘤转移密切相关,参与肿瘤发生和发展[14]。miR-367 位于染色体4q25位点,骨肉瘤、胰腺癌中miR-367表达增加,miR-367高表达对癌细胞迁移、侵袭具有诱导作用,miR-367 抑制剂可以消除这种致癌作用[15-16]。lncRNA癌易感性候选基因2(CASC2)通过下调miR-367 还可抑制肝癌细胞的迁移、侵袭和上皮间质转化进程[17]。本研究发现,肺癌组织中miR-367 表达增加,LINC00710 对miR-367 具有靶向负调控作用。过表达miR-367可提高A549细胞增殖、迁移和侵袭能力,表明miR-367在肺癌中发挥致癌作用,与Xiao等[18]报道一致。PTEN已被证实在肺癌中表达降低或缺失,恢复PTEN 表达能够抑制肺癌细胞增殖、迁移和血管生成[19-20]。本研究证实,miR-367 靶向负调控PTEN 表达,与Ling 等[21]报道在黑色素瘤葡萄膜细胞中两者靶向关系一致。本实验显示,过表达miR-367 显著减弱LINC00710 过表达对A549细胞增殖、迁移和侵袭的抑制作用以及对PTEN 表达的促进作用,进一步说明LINC00710 靶向miR-367可抑制肺癌A549细胞增殖、迁移和侵袭。
综上所述,LINC00710通过靶向miR-367/PTEN轴可抑制肺癌A549细胞增殖、迁移和侵袭,这对今后肺癌基因治疗具有一定的参考价值。
猜你喜欢
环球时报(2022-09-20)2022-09-20
中国糖料(2022年3期)2022-07-04
中国农学通报(2022年12期)2022-06-01
中国糖料(2022年2期)2022-04-06
天津医科大学学报(2021年4期)2021-08-21
广东蚕业(2021年1期)2021-03-18
中学生物学(2019年7期)2019-10-17
江苏农业科学(2019年23期)2019-03-03
医学信息(2016年29期)2016-11-28
小资CHIC!ELEGANCE(2015年14期)2015-09-23
