
Cyanobacterial extracellular alkaline phosphatase: detection and ecological function*
2022-10-08 02:23:28LinglingWANChunleiSONGYiyongZHOUXiuyunCAO
Lingling WAN, Chunlei SONG, Yiyong ZHOU, Xiuyun CAO
Key Laboratory of Algal Biology, State Key Laboratory of Freshwater Ecology and Biotechnology, Institute of Hydrobiology,Chinese Academy of Sciences, Wuhan 430072, China
Abstract Dissolved inorganic phosphorus is an important form of directly bioavailable phosphorus for cyanobacteria in natural water. Dissolved organic phosphorus could be used by cyanobacteria via alkaline phosphatase, which is produced mainly by bacteria and also cyanobacteria itself. Herein, we review the current knowledge of extracellular phosphatase excreted by cyanobacteria, highlighting the development of detection method and its ecological roles in regulating phosphorus cycling in freshwater systems, which is based on reports for around 100 species of cyanobacteria. Recommendations are suggested concerning the extracellular phosphatase produced by bloom-forming cyanobacteria in terms of the ecological role,followed by a discussion of the future prospects for the study.
Keyword: alkaline phosphatase; phosphate; dissolved organic phosphorus; cyanobacteria; ecological role
1 INTRODUCTION
Harmful cyanobacterial blooms in lakes are serious aquatic environmental problems. Phosphorus (P) is one of the limiting elements of phytoplankton growth in natural waters (Schindler, 1977; White et al.,2003). Dissolved inorganic P (DIP) is the preferred form of directly bioavailable P for phytoplankton(Cotner and Wetzel, 1992; Björkman and Karl, 1994;Baken et al., 2014). Dissolved organic P (DOP)maintains the supply of DIP via alkaline phosphatase catalyzing (Berman, 1970; Cao et al., 2018; Lim et al., 2018; Feng et al., 2020). Alkaline phosphatase(APase, EC 3.1.3.1) is a type of organophosphate hydrolase, which could catalyze the hydrolysis of phosphate ester bonds containing C-O-P ester bond to release orthophosphate (Pi) (Hoppe, 2003). The substrates of APases include phosphate glycosides,phosphomonoesters, and phosphate diesters, etc.(Bentzen et al., 1992; Yamaguchi et al., 2005), which contributed over 75% of DOP in the water (Kolowith et al., 2001; Young and Ingall, 2010; Yates et al.,2019). In addition to phototrophic and heterotrophic prokaryotes, protozoa and zooplankton, phytoplankton was found to be able to produce extracellular APases(Hoppe, 2003). Despite there have been numerous studies on the eco-physiological responses to P and increasing researches on genomics in bloom-forming algae in the past decades, few attempts to synthesize information in terms of extracellular APases produced by cyanobacteria were reported. Generally,extracellular enzymes are principally located outside the cell membrane and no longer in contact with their producers, while ecto-enzymes were def ined as the enzymes located outside the cellular outer membrane,but still in contact with the cell (Chróst, 1991).Here, we consider that extracellular phosphatase includes extracellular (or free) and ecto-phosphatase(Fig.1). Previously, Jansson et al. (1988) and Hoppe(2003) have systematically reviewed the origin,characteristics, and function of phosphatases in lake and ocean. Physiological ecology and molecular mechanisms responding to P status of ecto-APases were subsequently summarized in marine typical bloom-forming eukaryotic algae (Dyhrman, 2005;Lin et al., 2016). In this paper, we give a brief review of the determination method development in cyanobacterial extracellular phosphatase and its ecological function both in fresh and marine water.
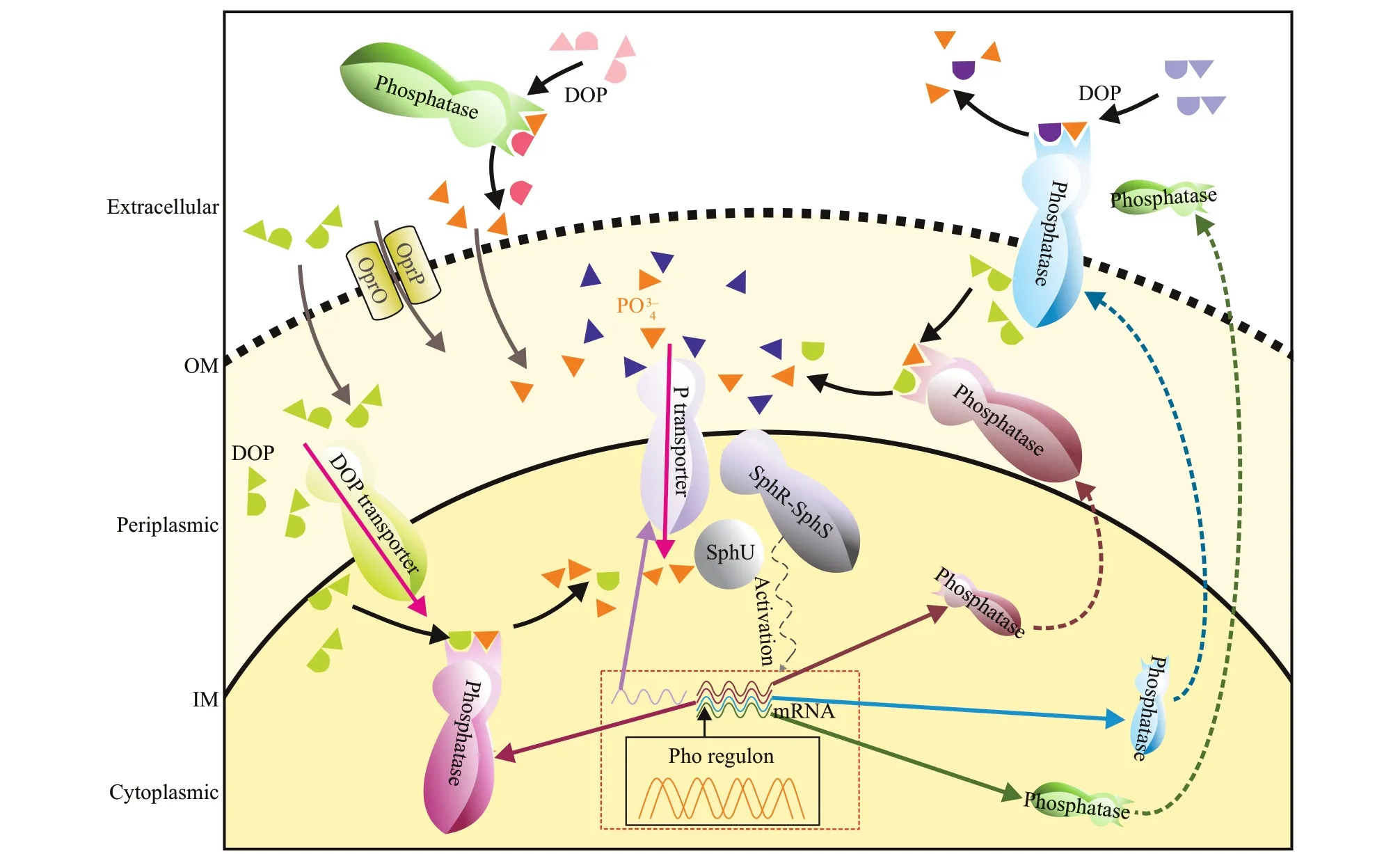
Fig.1 Subcellular localizations of alkaline phosphatase and its regulation by Pho regulons in cyanobacteria
2 DETECTION OF EXTRACELLULAR PHOSPHATASE FROM CYANOBACTERIA
Since Steiner (1938) f irst discovered that DOP could be enzymatically hydrolyzed in lakes, the research on extracellular phosphatases in water bodies have been a rapid development (Berman,1970; Zhou and Zhou, 1997; Luo et al., 2009;Zhang et al., 2016; Wang et al., 2021). Tracing the origin or producer of extracellular phosphatase is of great ecological signif icance. The accuracy mainly depends on the advances in the analytical methods,while extracellular phosphatase from cyanobacteria could be detected by axenic culture and enzyme purif ication. In-situ detection by enzyme labelled f luoresce (ELF) technology and molecular biological methods could give more solid evidence for the origin of extracellular phosphatase.
2.1 Extracellular phosphatase activity analysis from cyanobacteria
Cyanobacteria can produce APases attached to the cellular outer membrane or dissolved in the medium (Healey and Hendzel, 1979a, b; Smith and Kalff , 1981; Vaitomaa et al., 2002; Luo et al.,2009). Measurements of APase activity (APA) were eff ective means to quantitatively characterize APases in the f ield investigations and indoor experiments(Lehman et al., 2013; Yuan et al., 2016; Wan et al., 2019; Zhou et al., 2021). Determination of APA in the lab experiment can further prove the relationship between axenic cultured cyanobacteria and the origins of phosphatases to a certain degree,such as the contribution of cyanobacteria to soluble extracellular APases. For example, in theAnacystisnidulansandSynechococcusculture medium, APA was detected using para-nitro-pheneye phosphate(pNPP) as substrates (Ihlenfeldt and Gibson, 1975).APA detected using pNPP as substrates were related to cyanobacterial colonies (Rivularia) in the British rivers (Livingstone and Whitton, 1984). The same method was later used to detect APA fromNostoclinckia,Nostocmuscorum, andTrichodesmiumin the f ield survey and lab culture (Kumar et al., 1992; Stihl et al., 2001). Besides pNPP, other organic P compound could also be used as substrates. For example, bisp-nitrophenyl phosphate (bis-pNPP) was used to determine cell membrane attached and dissolved APA inNostoccommuneUTEX 584 culture (Whitton et al., 1990). Later on, 50 species of cyanobacteria were found to secrete extracellular APases using the same assay protocal (Whitton et al., 1991). Since the new century, 4-methylumbelliferyl phosphate(MUFP) has been used as substrate to determine APA in the f ield survey and lab cyanobacterial culture(Vaitomaa et al., 2002). pNPP and MUFP methods mainly focused on the liberation of Pi hydrolyzed by phosphomonoesterase, while less attentions were paid to phosphodiesterase (Sirová et al., 2013),which is a non-negligible enzyme involved in the organophosphorus hydrolysis pathway (Accoroni et al., 2017; Srivastava et al., 2021). Therefore, bis-pNPP or bis-(4-methylumbelliferyl) phosphate (bis-MUFP)was still suggested to access the phosphodiesterase activity and thus indicate the sources of organic P(Sirová et al., 2013). The accuracy of APA was also constrained by several factors, e.g. assay substrate,incubation time, pH, and sample sources, etc.(Hernández and Whitton, 1996). The peak month of APA ofRivulariacolonies was observed to be diff erent by using pNPP and MUFP as the sole substrate in intertidal pools at Tyne Sands, Scotland (Yelloly and Whitton, 1996). The comparison experimental results showed that the maximum release of the product (p-nitrophenol) appeared in over 20 min and at slightly acid pH, while the maximum 4-MUF appeared in more than 40 min and at neutral pH(Hernández and Whitton, 1996). Cyanobacterial blooms always occurred in alkalescent water bodies,and higher growth rates of cyanobacteria were also observed in the alkalescent culture mediums (Hong and Lee, 2008; de Souza Santos et al., 2011; Fang et al., 2018; Wei et al., 2022). Therefore, the MUFP method may be more suitable for the detection of APA from cyanobacteria.
Also of note is that, the sources of APases cannot be distinguished by pNPP and MUFP methods both in f ield investigations and indoor experiments,indicating that the contribution of APA from algae will be overestimated if the contribution of bacteria cannot be ruled out. Reducing the growth of bacteria by adding antibiotic or using UV light is an eff ective way to exclude its interference to algal APA in the cultivations (Gerloff et al., 1950; Harke et al., 2012;Li et al., 2015b). Purif ication of APases could relate to the APA and its producers partially (Martland and Robison, 1929; McComb et al., 1979). Jansson (1976)separated and purif ied the phosphatase in the cell and culture medium. Dissolved extracellular phosphatase in water might be derived from a zooplankton,Bosminaobtusirostris, by comparing the physical and chemical characteristics of the enzymes. Phosphatase isolated and purif ied from cyanobacteria (Anacystisnidulans) had diff erent characteristics from other phosphatases described previously (Ihlenfeldt and Gibson, 1975). Doonan and Jensen (1980)extracted attached and free APases from eighteen cyanobacteria and demonstrated the inducibility of eleven of them. APases, purif ied from cyanobacteria(Arthrospiraplatensi) using Triton X-114, was proven to be a calcium-dependent protein (Asencio et al., 2012). The purif ication method has not been widely used in the f ield surveys of phytoplankton ecology or environmental science, since it is complex and ineffi cient to distinguish between intracellular and extracellular phosphatases attached to the cell(Malherbe et al., 2019). Size fraction measurement of APA was a rough way to evaluate the contributor of particle (including cyanobacteria) APases (Dyhrman and Ruttenberg, 2006; Lim et al., 2018). If the large particle size contributed most to the total APA,extracellular APases were assumed being mainly contributed by the dominant or cultured cyanobacteria(Raoui et al., 2002; Wan et al., 2019).
2.2 Visualization of extracellular phosphatase
Enzyme labelled f luorescence (ELF) method provided an artif icial substrate to visually detect extracellular APases in cyanobacteria or algae.The basic principle of this method is as follows:ELFTM97 phosphate (ELFP) is a specif ic substrate of phosphatase, which could be hydrolyzed by APase into Pi and f luorescent organic residues (ELFA). The insolubility of organic residues makes the precipitate attach to the enzymatic reaction, so the f luorescence (or confocal) microscope or f low cytometer can not only determine the presence of extracellular phosphatase of cyanobacteria or algae, but also mark the occurrence site (Dyhrman and Palenik, 1999). Extracellular phosphatase was f irstly determined by ELF method in the culture of marine algae (González-Gil et al., 1998).ELF method was introduced and developed to detect algal and cyanobacterial extracellular phosphatase in Czech reservoirs by Štrojsová et al. (2003, 2005).Nedoma et al. (2007) further proved that the ELF assay could ensure a high saturation of extracellular phosphatases in the marine (>99%) and fresh (>90%)waters. Since then, this method has been widely used in the monitor of extracellular phosphatase of the marine and freshwater cyanobacteria, which has led the utilization of f luorescence microscopy into the visualization of APases in the cyanobacterial cells(Table 1). Signif icantly, not all tested cyanobacteria displayed f luorescent precipitates of ELFA (Table 1,Dignum et al., 2004b; Rychtecký et al., 2015; Wan et al., 2019). Possible reasons include: f irstly, the evoked threshold concentration of P is variable for initiating the secretion of APases by diff erent cyanobacteria.The same P concentration or environmental condition might not trigger the secretion of APases for some cyanobacteria (Nausch, 1998; Dyhrman and Palenik,1999; Sebastián et al., 2004; Cao et al., 2007; Girault et al., 2013; Zhang et al., 2021). Secondly, some cyanobacteria may secrete dissolved APases, which cannot trace back to the producers by ELF assay(Wang et al., 2021). Overall, there were still some limitations in analyzing cyanobacterial extracellular APA: f irstly, absolute axenic conditions were diffi cult to achieve, as a result, the contribution of bacteria to enzyme activity cannot be ruled out. Secondly, the physiological, biochemical, and external conditions of cyanobacteria grown were not consistent.Therefore, the chemical measurements of APA have many objective disadvantages, and multiple methods must be taken into consideration to comprehensively evaluate the cyanobacterial APases.
2.3 Application of molecular biological methods
Development of molecular biological methods provided deep insights into the origin of extracellular phosphatase, if combined with the quantif ication and ELF technique. The dominant molecular model, used to describe transformations and assimilations of P, has been the Pho regulon of the gram-negative bacteriumEscherichiacoli(Vershinina and Znamenskaya,2002). So far, the most important APases in prokaryotes include PhoA, PhoD, PhoV, PhoX, and atypical phosphatases (Luo et al., 2009; Kageyama et al., 2011; Lin et al., 2016). The general elements of the Pho regulon in cyanobacteria and its regulatory mechanisms might be similar to heterotrophic bacteria (Fig.1). The full-length of APase genes were sequenced and expressions of these genes were quantif ied under diff erent environment in the past two decades. The corresponding complementary DNA and RNA sequencing was coupled to a comprehensive metabolomics survey afterwards. Gene sequence analysis has revealed thatphoDencodes APase inAnabaena(Singh et al., 2015). In addition, the genes encoding APases inAnabaena(later namedDolichospermum) includephoA,phoD, andphoS(Liu and Wu, 2012).phoAandphoXhave been also reported in pico-cyanobacteria or cyanobacteria(Vershinina and Znamenskaya, 2002; Su et al.,2003; Moore et al., 2005; Sebastian and Ammerman,2009; Tetu et al., 2009; Kathuria and Martiny, 2011;Harke et al., 2012). Atypical APase genes like other cyanobacterialphoAand a classicalphoAgene are predicted in the genome ofAnabaenasp. PCC 7120(Luo et al., 2010).
The development of protein sequencing technology has further revealed the structures and functions of cyanobacterial APases. Ray et al. (1991) found an atypical APase inSynechococcussp. PCC7942,whose size (145 kDa) is larger than the previously reported one (47-87 kDa). The PhoA-type APase has a Zn2+cofactor, while PhoX associated with unculturedProchlorococcusis an active phosphatase with a Ca2+cofactor (Kathuria and Martiny, 2011).PhoX is a monomeric enzyme activated by Ca2+and Fe3+(Majumdar et al., 2005; Monds et al., 2006;Yong et al., 2014) with a lower substrate specif icity for C-O-P bonds, such as nucleotides, phosphorylated carbohydrates, and amino acids (Zaheer et al., 2009).PhoX inMicrocystisaeruginosaFACHB7806 is strongly activated by Mg2+, followed by other divalent ions (like Co2+, Ca2+, Zn2+, and Mn2+), but it is inhibited by Ni2+(Hong et al., 2021). Proteome results suggest that PhoA (Mg2+and Zn2+) and PhoX(Ca2+) has been expressed inSynechococcussp. WH 8102 (Cox and Saito, 2013). PhoD fromAphanothecehalophyticais a hydrolase activated by Ca2+that can hydrolyze phosphomonoesters and phosphodiesters(Kageyama et al., 2011). PhoD from a unicellular N2-f ixing cyanobacteria (Halothecesp. PCC 7418) shows the connection between Ca2+and Fe3+, which harbors eight copies of APase encoding genes (Fernández-Juárez et al., 2019). PhoV is a hydrolase activated by Zn2+, which can hydrolyze phosphate monoesters and has a wide range of adaptation to pH, but it is inhibited by Mn2+(Wagner et al., 1995).

Table 1 Extracellular phosphatases of cyanobacteria evidenced by ELF method
The detection of APase encoding genes and their expression levels enable the interpretation of APA and visualization of APases more reasonable.For example, the increased expression of putative APase gene (phoX) inM.aeruginosaunder low P supply evidenced that the extracellular phosphatases originated fromM.aeruginosa, together with a signif icantly positive correlation between P concentrations and APA in a monocultural experiment(Harke et al., 2012). Similarly, up-regulatedphoXgene of the predominantMicrocystiswas observed in low-P regions of Lake Erie (Harke et al., 2016). In respond to the decreasing P concentrations, bloomformingHydrocoleumsp. elevated the expression level ofphoAgene (Moisander et al., 2022). The changes in the expression levels ofphoA-like andphoDgenes well explained the increase of APA ofRaphidiopsismediterraneaandPlanktothrixagardhiiin the P-starvation conditions (Aguilera et al., 2019). Increasing the expression level of cyanobacterial APase genes in response to low P supply is species specif ic in terms of the diff erences in genes copy numbers and gene types (Harke et al.,2012; Liu and Wu, 2012; Lin et al., 2018; Aguilera et al., 2019; Willis et al., 2019). For example,phoXgene was not identif ied inMicrocystisweisenbergiibut inM.aeruginosa(Harke et al., 2012).PhoA,phoD, andphoXgenes were identif ied inAnabaenacylindricaFACHB-170 (Lin et al., 2018). Even thoughRaphidiopsisraciborskiiCS-505 and CS-506 harbored the same gene type, the copy numbers of theirphoAgene were also diff erent (Willis et al.,2019). Furthermore, the combined observations in transcriptional changes and metabolic homeostasis inM.aeruginosaprovide novel and extensive insights into the complex cellular interactions that take place in this important bloom-forming organism(Steff en et al., 2014). Therefore, comprehensively assess molecular aspects of phosphatase encoding gene and its expression, transcriptional patterns in the important bloom-forming cyanobacteria might expand our knowledge in understanding its bloom mechanism.
3 ECOLOGICAL FUNCTIONS OF CYANOBACTERIAL EXTRACELLULAR PHOSPHATASES
3.1 Indicating P def iciency
Phytoplankton will produce extracellular phosphatase when suff ering from P def iciency in natural population and lab experiment (Schindler,1977; Healey, 1978; White et al., 2003; Cao et al.,2018; Lim et al., 2018; Feng et al., 2020). Therefore,algal APA or total APA was inversely proportional to P concentration (Smith and Kalff , 1981; Chróst et al.,1984; Francko, 1984; Pettersson, 1985; Vrba et al.,1993). APA of cyanobacterial mats in the Caribbean showed signif icantly negative correlation to the P concentrations, as well as the P content of the mat(Rejmánková and Komárková, 2005). Furthermore,APA would be inhibited in the presence of suffi cient inorganic P supply (Kuenzler and Perras, 1965).This relationship was summarized as “inductionrepression” mechanism (Jansson et al., 1988). As a result, APA has often been recommended as an indicator of P starvation in planktonic systems(Healey and Hendzel, 1980; Istánovics et al., 1992;Rose and Axler, 1997; Thingstad et al., 1998; Jamet et al., 2001) or cyanobacterial mats (Rejmánková and Komárková, 2000).
Normalized APA, such as chlorophyll-specif ic total APA (Istánovics et al., 1992), surface-area-specif ic APA (Newman et al., 2003) was more sensitive enough to show changes in the P status of algae than total APA (Zhang et al., 2021). The specif ic activity divided by algal biomass can substantially ref lect the intrinsic catalytic effi ciency of the enzyme, and thus serve as a suitable indicator for the P nutrient status of water and algae (Kuenzler and Perras, 1965; Perry,1972; Fitzgerald and Nelson, 1975; Moegenburg and Vanni, 1991; Istánovics et al., 1992). On the other hand, the reliability of using APase as an indicator of P def iciency in phytoplankton was questioned(Cembella et al., 1982). For example, in Florida Bay, APA was highest during cyanobacterial blooms and particulate APA was related to the bloom, but APA/chlorophyllaand dissolved APA showed no correlation to the bloom (Koch et al., 2009). Factors interfering APA as the indicator of P def iciency include variety of extracellular phosphatase producers such as bacteria and zooplankton (Koch et al., 2009),enzyme substrate diversity (Sharma et al., 2014;Zhang et al., 2020b) and its eff ect on APA (Fonseca-De-Souza et al., 2008; Harke et al., 2012; Li et al.,2015a), extracellular secretions such as extracellular toxins (Bar-Yosef et al., 2010; Dobronoki et al., 2019;Lu et al., 2021), species specif ic responses (Olsen et al., 1989; Wan et al., 2019), external P threshold concentrations for activated response (Pick, 1987;Ruiz et al., 1997), environmental factors, such as light (Giraudet et al., 1997; Rychtecký et al., 2015;Yadav et al., 2016) and temperature (Bai et al., 2021;Ivančić et al., 2021), and the level of intracellular P content (Olsson, 1983; Whitton et al., 1990; Zhang et al., 2020a). Furthermore, the activity of endogenous enzymes might be overestimated because of the exogenous input (Stevens and Parr, 1977).
Biochemical components unrelated to P can also cause increase or decrease in APA (Wilkins, 1972;Francko and Wetzel, 1982). In addition to diff erences in gene homology, the metal ions, located in the active sites of phosphatases, are also diff erent (Luo et al., 2009). Therefore, the lack of external related metal elements will also aff ect the synthesis and function of cyanobacterial extracellular APA (Singh et al., 2006; Cox and Saito, 2013). APA contributed by dissolved APases could be spontaneously released during cyanobacterial blooms, other than driven by the P limitation. This explained coexistences of high Pi concentrations and high potential APA in water observed previously (Wang et al., 2021). The activity of dissolved phosphatases can be maintained for several weeks (Jansson, 1981; Olsson, 1983),which means that enzymes produced in one place may function in a wider range of waters. Shortly, the relationships between APA and ambient bioavailable P concentration are complicated. There are several limitations to use extracellular APA as a general indicator for describing P def iciency of phytoplankton(Cao et al., 2010). The multiple complex regulatory factors make it cautious when using extracellular APA as indicators of P def iciency.
3.2 Alleviating P stress
Algae could use extracellular phosphatases to obtain nutrients and grow normally with a diff erent organophosphates supply (Ruiz et al., 1997). It was found that 17%-82% of the P absorbed into phytoplankton was from organophosphorus in the Sargasso Sea (McLaughlin et al., 2013). In the eutrophic Lake Nantua, the Pi released from DOP via APases could temporarily maintain the P supply for the algae dominated byOscillatoriarubescenswhile DIP was def iciency in the summer (Feuillade et al., 1990). Moreover, the growth of cyanobacteria could be mainly supported by hydrolyzing dissolved organophosphate via APase in Lake Taihu and other warm-monomictic lake (Gao et al., 2006; Prentice et al., 2019). While in the absence of inorganic P,organophosphorus, e.g. pesticides, can be used by f ilamentous-heterocystous cyanobacteria as the only source of P (Subramanian et al., 1994). In the Grangent reservoir, cyanobacteria often relied on the APase to mineralize organic P (Giraudet et al., 1999).The aggregates dominated byNodulariaspumigenain the Baltic Sea were the key contributor to P regeneration (Stoecker et al., 2005). The increasing biomass ofAphanizomenonin a P-limited summer and autumn was accompanied by high enzymatic activity of APase (Hadas et al., 1999). Filamentous cyanobacteria,NodulariaandAphanizomenon, from the Baltic Sea, showed higher APA with the decline of ambient Pi concentrations (Degerholm et al., 2006).Both ELF methods and molecular biology approaches supported the view thatCylindrospermopsisraciborskiicould use diff erent organophosphates to sustain growth when P was limited (Bai et al., 2014).Besides increasing potential APA, cyanobacteria may compensate for P stress by lowering Michaelis constant (Km) values of APases in the meanwhile(Zhang et al., 2021). During aMicrocystisbloom outbreak, a large number of P sources was hydrolyzed by extracellular APases and released to sustain the growth ofMicrocystis(Chuai et al., 2011), but the extracellular APases might be excreted mainly by bacteria rather thanMicrocystis(Dai et al., 2018).Therefore, APase plays a crucial role in the process of cyanobacterial growth and blooms, although it might be secreted by microorganisms rather than cyanobacteria themselves (Zhao et al., 2012).
4 FUTURE RESEARCH
4.1 To what extent can extracellular phosphatases alleviate P stress
Researchers, mainly focusing on bloom-forming cyanobacteria, hold a hypothesis that secreting extracellular phosphatase to use DOP is their key competitive advantage, when they realized that APA was regulated by P supply (Kelly et al., 2019; Rabouille et al., 2022; Zhang et al., 2022). Unexpectedly, cells that secrete extracellular phosphatase are often not cyanobacteria even dominated in phytoplankton assemblages (Rengefors et al., 2001; Štrojsová et al.,2003), which makes the above hypothesis diffi cult to understand. If we can answer the question to what extent extracellular APase can alleviate P stress, the puzzle could be solved. Unfortunately, the current detection method cannot achieve the goal because of obvious def iciencies. Firstly, it is diffi cult to determine to what extent it can be able to represent a natural substrate for APase when using artif icial synthetic organophosphates as the substrate. Therefore, the cyanobacterial extracellular APA, strictly speaking,can only ref lect the qualitative rather than quantitative relationship between APase and environmental variables (Siuda, 1984). To overcome this uncertainty,ecologists have used the radiometric method to determine the availability of P in the organic P pool of water bodies (Bentzen et al., 1992). Hernández et al.(1996) used32P-G6P as a substrate to explore a more sensitive method for determining APA while there is still a problem since the radiometric element method might be selectively absorbed by phytoplankton. As a result, the function of phosphatase to alleviate P limitation may be overestimated for cyanobacteria.So, a combination study of cyanobacteria from molecular biology, biochemistry, f ield investigation,or indoor culture experiments with a more accurate test method is urgently needed.
4.2 Determine ecological function of extracellular phosphatases on species level
Excretion of extracellular APases is species specif ic among cyanobacteria (Rengefors et al., 2003; Dignum et al., 2004b). Cyanobacteria respond to low P supply by combining with other physiological process.Several perspectives deserve noting, including interspecif ic response between toxin and non-toxin producing cyanobacteria; the connection between toxin and extracellular APases producing; combining study among the diff erent response mechanisms (e.g.,P uptake, storage, regulating P demand, etc.). Since APases can be active, which are sometimes irrelevant to ambient P concentrations, other functions of APases are indeed worthy of attention at this moment, e.g. to constrain pigment biosynthesis, photosynthesis, fatty acid biosynthesis and cell division, which are of great signif icance in phytoplankton ecology. Although single-cell phosphatase of phytoplankton could be visualized and quantif ied by the ELF technology(Diaz-de-Quijano et al., 2014, 2020). Relation between transcriptional levels of APase encoding genes and the functional microbial compositions might give deep insight to the enzyme producers or origin (Bai et al., 2014; Dai et al., 2018). Integration of molecular biology and ELF assay is advised to give a comprehensive and reasonable result for the contributors of extracellular phosphatases at the species level. In summary, future research on specif ic species ecophysiology will benef it from a growing suite of tools available for assessing the activity and subcellular location of APase in f ield populations or cultures and ultimately the work done with specif ic species will be useful for studies of other harmful ones.
5 DATA AVAILABILITY STATEMENT
All data generated and/or analyzed during this study are included in this published article.
 Journal of Oceanology and Limnology2022年5期
Journal of Oceanology and Limnology2022年5期
- Journal of Oceanology and Limnology的其它文章
- Comparison of three f locculants for heavy cyanobacterial bloom mitigation and subsequent environmental impact*
- Proteomic analysis provides insights into the function of Polian vesicles in the sea cucumber Apostichopus japonicus post-evisceration*
- Key physiological traits and chemical properties of extracellular polymeric substances determining colony formation in a cyanobacterium*
- Involvement of the ammonium assimilation mediated by glutamate dehydrogenase in response to heat stress in the scleractinian coral Pocillopora damicornis*
- UV-B irradiation and allelopathy by Sargassum thunbergii aff ects the activities of antioxidant enzymes and their isoenzymes in Corallina pilulifera*
- Full-length transcripts facilitates Portunus trituberculatus genome structure annotation*