
不同剂量矿质肥对丘北辣椒根际细菌群落结构的影响
2022-09-29 08:11:04王灿袁恩平张雪廷王绍祥赵水灵李云李罡
湖北农业科学 2022年16期
王灿,袁恩平,张雪廷,王绍祥,赵水灵,李云,李罡
(文山州农业科学院,云南 文山 663000)
辣椒(Capsicum annuumL.)作为中国普遍栽培的蔬菜,是西南边陲农民增收的经济作物之一。辣椒适应性广、产业链长,是日常生活中常见的蔬菜,不仅营养丰富而且具有较大商业开发潜力,栽培面积逐年上升。云南省作为辣椒种植大省,其中,文山州种植面积较大,占云南省辣椒种植面积的81.32%[1],文山州特殊的地理气候环境造就了丰富的辣椒资源,如丘北辣椒、小米辣、涮辣等。丘北辣椒有400多年的种植历史,1983年12月被对外经济贸易部授予丘北辣椒金质奖章[2],是文山州除三七、烤烟外大力推进的产业,在增加农民收入、促进农业产业结构调整上起到了积极作用[3]。研究者常把研究聚焦于N、P、K肥施用比例对丘北辣椒生长发育的影响上,而忽略了微量矿质元素如钙、镁、铁等的作用。韩明珠等[4]通过试验得到丘北辣椒最佳施氮量为180 kg/hm2。湛方栋等[5]研究表明肥料中N∶P2O5∶K2O为12∶10∶14的处理增加的丘北辣椒土壤微生物数量最多。
本研究以云南省文山州地方特色品种丘北辣椒为研究对象,采用16S rDNA扩增子测序,旨在从微生物群落结构多样性角度探究矿质肥在丘北辣椒苗期的应用规律,为云南省文山州丘北辣椒根际生长微环境及微生物群落定殖调控机理提供参考。
1 材料与方法
1.1 材料
1.1.1 供试品种辣椒为文干椒1号,由文山州农业科学院提供。
1.1.2 供试材料矿质肥料“矿物之星”由日本群马长石公司提供,是一种天然矿物质(石矿粉中提取)和腐殖酸复合的制剂,其主要矿质成分见表1。育苗基质为商品育苗基质,‘湘正农科’牌商品育苗基质由湖南农业大学湘晖农业技术研究所研制,主要成分为草炭土。

表1 矿质肥料主要矿质组成成分及pH
1.2 方法
1.2.1 盆栽试验及样品采集试验地点为文山州农业科学院,时间为2020年12月至2021年5月。将发芽整齐一致的辣椒种子播种于苗盘(21孔)中进行盆栽试验。试验共5个处理,分别是CK处理(0 g/L)、处理A(15 g/L)、处理B(30 g/L)、处理C(45 g/L)、处理D(60 g/L)。每个处理3盘为1次生物学重复,并进行常规育苗管理。苗龄为51 d时采用五点取样法选取植株,将植株整株拔出抖落附着土壤基质后,用单独无菌刷刷取粘附在根表面的土壤基质,并将各样品混合后放入50 mL无菌离心管中,迅速置于干冰中保存运输,后置于-80℃冰箱保存备用,随后将抖落土壤基质收集用于土壤养分检测。
1.2.2 DNA浓度检测使用Magen核酸提取试剂盒(MagPure Soil DNA KF Kit),Equalbit dsDNA HS Assay Kit检测DNA浓度。以30~50 ng DNA为模板,采用GENEWIZ公司设计的PCR引物扩增原核生物,16S rDNA上包括V3及V4的2个高度可变区,引物系列如下。F:5′-CCTACGGRRBGCASCAGKVRVG AAT-3′和R:5′-GGACTACNVGGGTWTCTAATCC-3′。总反应体系为50 μL,5 μL Buffer,4 μL DNA模板,4 μL Mg2+,4 μL dNTP,上、下引物各1 μL,1 μLTaqDNA聚合酶,30 μL去离子水。细菌PCR反应程序为95℃预变性7 min,95℃变性60 s,56℃退火60 s,共35个循环,72℃延伸15 min,4℃保存,通过Illumina MiSeq进行双端测序。
1.2.3 数据质控与分析原始数据整理采用DPS 7.05软件。测序数据质量优化使用Cutadapt(v1.9.1)、Vsearch(1.9.6)、Qiime(1.9.1)软件分析。测序数据优化后质量如表2所示。

表2 测序数据质量
2 结果与分析
2.1 不同处理辣椒幼苗根际细菌群落结构分析
5个样本中共同含有的OTU为130个,除处理C含1个独有OTU外,其余处理均无独有OTU(图1a)。在细菌门水平下,共有10个物种种类,变形菌门(Proteobacteria)为优势菌,其次是拟杆菌门(Bacteroidetes)。值得注意的是,在5个样本处理中随矿质肥料剂量的增加,变形菌门数量占比呈下降趋势,最高的是CK,达65.70%,最低的是处理D,为54.04%;而厚壁菌门(Firmicutes)数量则呈上升趋势,最高的是处理D,达7.90%,最低的是处理A,为1.38%(图1b)。在属水平下(排名前十),CK中鞘氨醇单胞菌属(Sphingomonas)相对丰度最高,达16.01%。此外,处理C、处理D中f_Chitinophagaceae_Unclassified相对丰度分别是13.52%和10.07%,高于处理A(6.72%)、处理B(4.27%)及CK(6.13%)。芽孢杆菌属(Bacillus)中处理C、处理D相对丰度分别是3.56%、7.38%,高于处理A(0.86%)、处理B(0.82%)及CK(1.69%)。但 在Pseudolabrys中 处 理A(4.85%)、处理B(4.61%)及CK(5.39%)高于处理C(3.73%)和处理D(3.08%)(图1c)。图1d显示了基于Brary-Curtis距离矩阵聚类情况,其中处理A、处理B、CK聚为一类,处理C、处理D聚为一类。
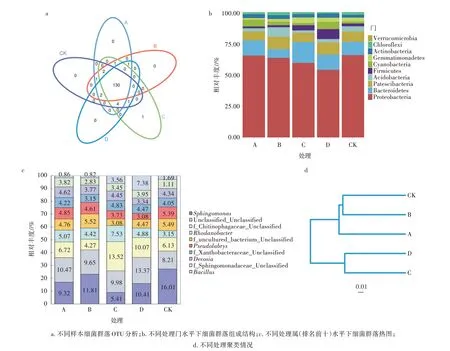
图1 不同处理细菌群落组成分析
2.2 不同处理辣椒幼苗根际细菌群落多样性分析
α多样性比较中,Ace和Chao1是反映菌群丰富度的指标,数值越小表明丰富度越低。由表3可知,处理B、处理C、处理D的Ace和Chao1指数低于处理A和CK,说明处理B、处理C、处理D的丰度低于处理A和CK。这表明矿质肥施入量的增加降低了根际细菌群落的物种丰度。Shannon、Simpson是反映菌群多样性的指标,其数值越小表明群落多样性越低。处理A、处理B、处理C的Shannon和Simpson指数均高于处理D和CK,说明处理D和CK降低了其根际细菌的多样性。

表3 不同处理细菌多样性指数
基于Brary-Curtis距离矩阵进行PCoA作图,分析得知第一主坐标对样本差异的贡献率为63.24%,第二主坐标对样本参与的贡献率为15.09%(图2)。图中样本点距离的远近代表了样本中微生物群落的相似性,距离越近,相似度越高。如图2所示,处理A与处理B较近,处理C与处理D较近,CK与其他处理较远。说明处理A、处理B的细菌群落结构相近,处理C、处理D的细菌群落结构相近。
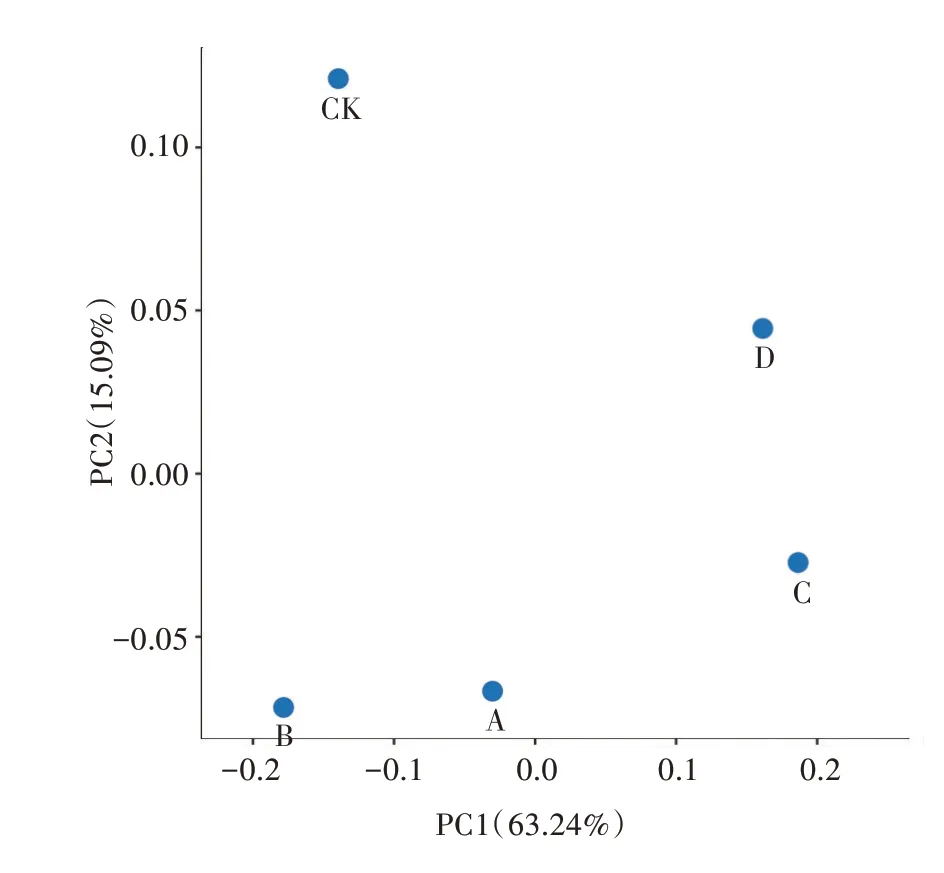
图2 不同处理细菌群落PCoA分析
2.3 不同处理细菌群落与土壤基质环境关联性分析
对各处理土壤样本进行检测后发现,N(氮)、P(磷)、K(钾)、Na(钠)、Mg(镁)、pH、EC(电导率)等化学指标随添加剂量的增加呈上升趋势(表4、表5)。如图3a所示,在门水平下,变形菌门与N、Mg呈显著负相关,与P、Na、B(硼)、Mn(锰)、EC呈极显著负相关;厚壁菌门则与Mn、EC呈显著正相关,与P、B呈极显著正相关。如图3b所示,在属分类水平下,鞘氨醇单胞菌属与Si(硅)呈显著负相关;Unclassified_Unclassified与P、B呈显著正相关;Pseudolabrys与P、K、C/N(碳氮比)、B、Ca(钙)、Fe(铁)、Mg、pH呈显著负相关,与N、Na、Mn、EC呈极显著负相关;Bacillus则与P、B呈显著正相关。

图3 不同处理细菌群落与土壤化学性质的相关性分析

表4 不同处理土壤基质化学性质
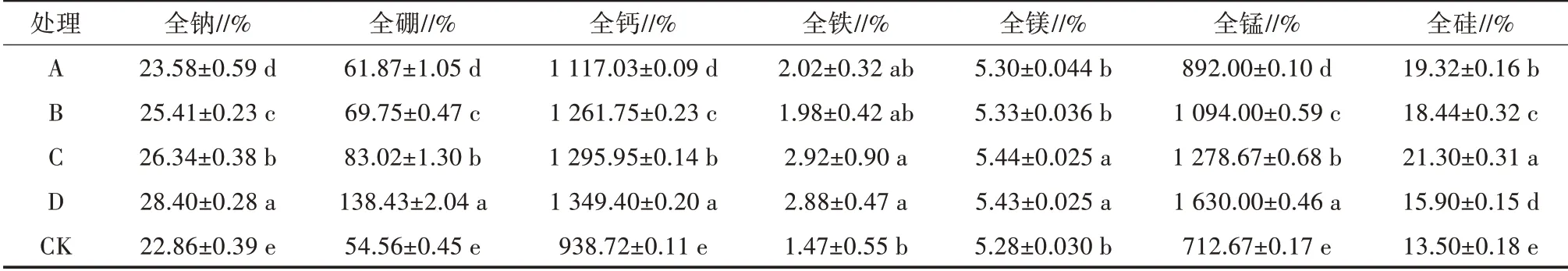
表5 不同处理土壤基质微量元素
3 小结与讨论
土壤微生物群落与土壤质量息息相关,是维持土壤质量的重要因素,微生物多样性在一定程度上可以反映土壤的肥力水平和施肥效果[6]。前期研究表明当土壤基质中矿质肥施用量为15~30 g/L时,育苗效果显著高于0 g/L和45~60 g/L,可溶性糖、根系活力、土壤蔗糖酶、脲酶、磷酸酶活性均高于其他处理[7-9]。进一步对各处理根际微生物群落高通量测序分析,结果表明,在5个样本处理中随矿质肥料剂量的增加,变形菌门占比呈下降趋势,占比最高的是0 g/L处理,达65.70%,占比最低的是60 g/L处理,为54.04%;厚壁菌门占比则呈上升趋势,最高的是60 g/L处理,达7.90%,占比最低的是15 g/L处理,为1.38%。说明高浓度处理降低了变形菌门的丰度而提高了厚壁菌门的丰度。这与孔帆[7]的研究结果一致,长期施用高浓度肥料提高了厚壁菌门的相对丰度。在属水平中通过对排名前十的细菌属聚类分析表明,浓度较低的CK、处理A、处理B(0~30 g/L)聚为一类,高浓度处理C、处理D(45~60 g/L)聚为一类。这表明添加剂量的大小对辣椒根际微生物群落结构有显著的影响。在α多样性分析中,处理B、处理C、处理D的Ace和Chao1指数低于处理A和CK,说明在高浓度处理下根际微生物群落丰度降低,在多样性比较中处理D的Shannon和Simpson指数低于处理A、处理B、处理C,表明对丘北辣椒施用高浓度矿质肥降低了微生物群落的多样性。李春越等[10]研究表明长期施用单一化肥使土壤酸碱度发生改变,对微生物的生命活动产生抑制作用。此外,郭振[11]研究发现,细菌α多样性和β多样性均表明不同施肥措施使细菌群落结构存在显著差异,长期施用有机肥处理的细菌群落多样性较为接近,而不施肥处理和单施化肥处理的细菌群落则单独成簇。
土壤中营养成分、物理性状不同可能会导致微生物群落结构差异[12],Edwards等[13]研究指出,在控制条件下水稻根际和根内微生物群落结构受土壤类型和宿主植物基因型的共同调控。刘东海等[14]在小麦根际微生物真菌群落中研究发现施有机肥可提高微生物多样性,其原因是有机肥改变了土壤理化性质。Dombrowski等[15]研究多年生黄花亭荠在自然和控制条件下根际细菌群落组成的因素试验中,通过主坐标典范分析和单因素方差分析指出,土壤性质能够解释根内细菌15%的群落变异性,而环境条件和植物基因型最多解释了11%的群落变异性,且土壤类型对根内细菌门分类水平上的影响更加显著。陈梅春等[16]研究发现茉莉花植株内生细菌群落受土壤全氮、全钾、全磷和有机质含量影响,变形菌门数量与全钾含量呈负相关,与全磷、全氮和有机质含量呈正相关;放线菌门、拟杆菌门数量与全钾含量呈正相关,与全磷、全氮和有机质含量呈负相关。本试验在前人研究基础上分析了微量元素对辣椒根际微生物群落的影响,在处理样本中增加矿质肥添加量,各处理土壤基质中矿离子含量随之增加,微生物群落与环境因子相关性分析中,门水平下,变形菌门与N、Mg呈显著负相关,与P、Na、B、Mn、EC呈极显著负相关。厚壁菌门与Mn、EC呈显著正相关,与P、B呈极显著正相关。属分类水平下,鞘氨醇单胞菌属与Si呈显著负相关;Pseudolabrys与P、K、C/N、B、Ca、Fe、Mg、pH呈显著负相关,与N、Na、Mn、EC呈极显著负相关;芽孢杆菌则与P、B呈显著正相关。其中芽孢杆菌是一种革兰阳性杆菌。研究表明,芽孢杆菌具有较强的纤维素酶、蛋白酶、淀粉酶等多种活性,这些酶能够抑制或破坏周围其他菌群的生长[17]。在真菌性病害防治中,具有抗逆性强、抗菌广谱性、杀菌高效、代谢产物丰富等特点[18]。此外,芽孢杆菌在反刍动物胃肠道内也具有调节动物肠道菌群的功能[19]。这将为发掘利用对辣椒根际有益的微生物资源,控制潜在有害微生物,保持生态系统平衡,促进辣椒优质栽培生长奠定基础。
综上所述,向丘北辣椒苗期施用不同剂量矿质肥可显著改变根际细菌群落结构,表现为高浓度处理降低细菌丰度与多样性。除N、P、K、pH、EC外,微量元素对丘北辣椒根际细菌同样存在显著的影响,这对丘北辣椒根际细菌群落调控和肥料施用具有一定的指导意义。
猜你喜欢
中国比较医学杂志(2020年4期)2020-05-26 05:47:22
农村百事通(2019年21期)2019-12-06 06:33:17
水生生物学报(2019年4期)2019-07-20 08:08:10
生物安全学报(2019年3期)2019-02-15 16:54:12
川北医学院学报(2019年6期)2019-02-10 10:48:32
中成药(2018年11期)2018-11-24 02:57:08
食品工业科技(2014年9期)2014-03-11 18:15:44
文山学院学报(2012年4期)2012-03-25 13:07:28
植物营养与肥料学报(2011年3期)2011-10-24 06:14:44
中学生物学(2005年4期)2005-04-29 00:44:03
