
西南湿润区微灌猕猴桃果园不同尺度光利用效率与驱动因子
2022-09-26 06:25杨斌环崔宁博何清燕姜守政郑顺生黄耀威
排灌机械工程学报 2022年9期
杨斌环,崔宁博*,何清燕,姜守政,郑顺生,黄耀威
(1.四川大学水力学与山区河流开发保护国家重点实验室/水利水电学院,四川 成都 610065; 2. 四川省农业机械研究设计院,四川 成都 610066)
光利用效率(LUE)是衡量植被将辐射能转化为化学能的重要指标[1],也是研究生态系统碳循环和以LUE为参数评估总初级生产力的基础[2].以往关于LUE的研究工作多集中在生态学领域;近年来随着作物模型的发展,基于LUE评估农田下垫面光碳通量、农作物产量的模型也在不断发展[3],因此深入了解光碳通量控制机理,探索作物不同尺度LUE变化规律及其对环境因子的响应机制,对揭示全球气候变化下农田碳循环过程和作物产量预测具有重要意义.
国内外学者针对作物不同时空尺度LUE变化规律和影响因素开展了大量研究.在叶片尺度上,叶片瞬时光利用效率(LUEi)被定义为叶片净光合速率(Pn)与叶片瞬时光合有效辐射(PARi)的比值[4].LUEi被广泛应用于评价植物叶片对光能的利用能力.大多数植物LUEi在低光强时随光强增加而增加;当超过某个光强阈值点后,其值随光强增加呈非线性下降[5].在冠层尺度上,冠层光利用效率(cLUE)被定义为冠层截获的光合有效辐射(APAR)与总初级生产力的比值,被广泛应用于基于MODIS数据的模型模拟区域GPP[6];cLUE与气候条件、植被结构、土壤类型和植物年龄范围有关[7],其也取决于入射的太阳辐射,并且在晴朗的天空条件和高PAR条件下会达到饱和[8].在生态系统尺度上,生态系统光利用效率(eLUE)被定义为光合有效辐射与总初级生产力的比值,主要应用于基于涡度协方差的LUE(EC-LUE)模型将GPP扩展到生物群落和区域层面[9];eLUE受到环境因子(光合有效辐射、空气温度、大气干湿变化和土壤含水率)、生理因子(叶龄和气孔导度)、植被结构(叶面积指数和缝隙度)等因子的影响[10].STOCKER等[11]发现土壤含水率(SWC)、空气温度(Ta)和水汽压差(VPD)通过调控叶片气孔开度和相应的光合过程对eLUE影响显著.GITELSON等[12]研究表明植被指数(归一化植被指数(NDVI),叶面积指数(LAI))通过影响冠层光截获量对光合作用起到积极作用.LUE的主要驱动因子在不同时空尺度上可能因不同植被和生态系统而异[11].因此,有必要探索作物不同时空尺度LUE变化规律和影响因素.
四川成都平原是中国五大猕猴桃产区之一.近年来,随着西南地区经济的高速发展,猕猴桃种植面积仍持续增大,环境污染问题也备受瞩目.不科学的种植方式和农艺管理模式会改变猕猴桃的生长生理特性和最佳生长环境,这些生理环境因子的改变均会对猕猴桃水分利用效率和光能利用效率产生影响,进而对猕猴桃水分消耗、果实产量形成甚至品质产生影响[13].目前,国内外研究生理环境因子对猕猴桃水分利用效率的影响较多[14-16],而研究生理环境因子对猕猴桃光能利用效率的影响较少[17].因此,文中选取中国猕猴桃之都—蒲江县猕猴桃果园为研究区域,应用涡度相关系统、光合仪和气象站实测的光碳通量及相关环境数据,分析猕猴桃果园光利用效率在叶片、冠层和生态系统尺度上的变化规律,并应用通径分析方法量化生理、环境和植被因子对猕猴桃果园不同尺度光利用效率的影响,以期揭示西南地区农田生态系统碳循环过程,为猕猴桃果园生产力评估提供相关依据.
1 材料和方法
1.1 试验区域概况
研究区位于四川省成都市蒲江县猕猴桃产业示范基地(103°25′E,30°19′N).研究区属于亚热带季风湿润气候,多年平均温度为16.3 ℃,平均降水量为1 280 mm,年日照时数为1 122 h,相对湿度为84%;试验区地形属于浅丘区,土壤类型以黄壤土为主.研究对象为9 a生金艳猕猴桃果树,果树冠层高度为1.70~2.00 m,种植密度约为450株/hm2,猕猴桃生育期从3月中旬到11中旬.园区采用旋转倒挂微喷头灌溉,当果树根区土壤体积含水率低于25 %时灌水,喷头流量为80 L/h,每次灌溉时间设定为4 h,分为4个轮灌组.
1.2 试验观测和数据处理
1.2.1 猕猴桃叶片生理指标测定
采用Li-6400便携式光合仪对叶片生理指标及叶片表面气象指标进行测定.每次测定,在猕猴桃冠层东西方向遮阴和向阳叶各选取3片叶子,合计6片/棵,主要测定其净光合速率[Pn,μmol/(m2·s)]、蒸腾速率[Tr,mmol/(m2·s)]、气孔导度[gs,mol/(m2·s)]等指标及对应时刻的环境因子.测定时间为2018年猕猴桃生育期内的3个晴天(5月13日、6月5日和7月25日),时段为9:00—19:00,约2 h观测1次.
1.2.2 气象及土壤水分数据采集
气象数据由自动气象站采集,土壤水分数据通过传感器TDR-3测试,采集时间为2018—2020年猕猴桃全生育期,采集的数据包括总辐射(Rg,W/m2)、光合有效辐射(PAR,W/m2)、净辐射(Rn,W/m2)、土壤热通量(G,W/m2)、空气相对湿度(RH,%)、空气温度(Ta,℃)、土壤温度(Ts,℃)、降雨(P,mm)、2 m处风速(U2,m/s)以及土壤体积含水率(SWC,%).
1.2.3 叶面积指数测定及其数据处理
冠层叶面积指数(LAIc)采用LAI-2000冠层分析仪(Plant Cnopy Analyzer, Li-Cor)每1—2周进行测定.测定时间为2018—2020年猕猴桃全生育期,测量方法为在冠层下方50 cm处沿枝条伸展方向每30 cm间隔测1组,每棵树测10组,选择8棵树进行测量,不同时间间隔的冠层叶面积指数通过样条插值法插值成日值.猕猴桃果园下垫面属于林-草复合的系统,下垫面总的叶面积指数(LAIt)通过(MODIS 15A2H,https://modis.ornl. gov/data.html)下载,下载的叶面积数据为500 m×500 m分辨率,8 d时间间隔.研究中,采用TIMESAT软件对原始下载数据进行平滑校正,去除由于云层和降雨天气造成的异常值,并用线性内插法插补,最后8 d的LAIt通过样条插值法插值成日值.
1.2.4 水光碳通量观测及其数据处理
采用闭路涡度相关系统(CPEC,Campbell Scientific Inc, USA)观测猕猴桃果园区水光碳通量变化.CPEC系统安装在园区的中南部(距离园区边缘3 km),安装高度为8 m,满足风浪区1∶100的要求.观测时间为2018—2020年猕猴桃全生育期,原始的10 Hz通量数据经过Datalogger Support Software 4.4处理,输出为30 min的通量数据.处理方法包括二次坐标旋转,时间延迟校正,高低频损失修正,水汽和CO2通量的WPL密度校正(空气密度脉动校正).
1.3 研究方法
1.3.1LUE的计算
采用的LUE包括LUEi,cLUE和eLUE,其计算公式为
(1)
(2)
(3)
APAR=PAR(1-e-k·LAIc),
(4)
式中:LUEi为叶片瞬时光利用效率,μmol/μmol;PARi为叶片瞬时光合有效辐射,μmol/(m2·s);eLUE为生态系统光利用效率,g/MJ或mg/MJ;GPP为总初级生产力,g/(m2·d)或mg/(m2·s);PAR为光合有效辐射,小时尺度为MJ/(m2·h),日尺度为MJ/(m2·d),月尺度为MJ/(m2·月);cLUE为冠层光利用效率,g/MJ或mg/MJ;APAR为冠层截获的光合有效辐射即吸收的PAR,小时尺度为MJ/(m2·h),日尺度为MJ/(m2·d),月尺度为MJ/(m2·月);LAIc为冠层叶面积指数,m2/m2.
1.3.2 通径分析方法
通径分析是一种典型的多元统计分析方法,用于研究自变量对因变量的影响强度,其优势在于不仅可以量化自变量对因变量的直接影响,而且可以量化某一自变量通过其他自变量对因变量的间接影响.
通径分析的计算方法:对于具有一个因变量y和多个自变量xi(i=1,2,…,n)的线性相关系统,即
y=b0+b1x1+b2x2+…+bnxn.
(5)
基于式(5)可建立矩阵方程
(6)
(7)
式中:rxixj为自变量xi与xj的简单相关系数(j=1,2,…,n);Pxiy为xi对y的直接通径系数(量化的直接影响);bi为y对xi的偏相关系数;σxi和σy为xi和y的标准偏差.
rxixj·Pxiy表示自变量xi通过xj对y的间接通径系数(量化的间接影响).
2 结果与分析
2.1 叶片尺度瞬时光利用效率与驱动因素
猕猴桃叶片净光合速率Pn和瞬时光利用效率LUEi与光合有效辐射PAR的关系如图1所示,Pn与PAR呈显著的二次抛物线关系(R2=0.66,P<0.01),Pn随PAR呈先增后降的趋势;LUEi与PAR呈极显著线性关系(R2=0.82,P<0.01),LUEi随PAR增大而降低,变化幅度为0.006~0.026 μmol/μmol.

图1 猕猴桃叶片净光合速率Pn和瞬时光利用效率LUEi与光合有效辐射PAR的关系Fig.1 Relationship between net photosynthetic rate and instantaneous light use efficiency and photosynthetically active radiation of kiwifruit leaves
环境因子(Ta,PAR,VPD)和生理参数(gs,Pn)对猕猴桃叶片尺度瞬时光利用效率LUEi的影响见表1,表中A为影响量化值.通径分析结果表明PAR和Pn对LUEi均具有显著影响,量化的总影响分别为-0.80和-0.56(P<0.01);PAR对LUEi的作用主要来自于其对LUEi的直接影响(-0.99),Pn对LUEi的影响主要来源于其对LUEi的间接影响(-0.81);此外,gs和Ta对LUEi也有较高的间接影响(-0.41和-0.30),而VPD对LUEi影响不具有统计学意义(P>0.05).

表1 环境因子和生理参数对猕猴桃叶片尺度瞬时光利用效率(LUEi)的影响量化值Tab.1 Quantification values of effects of environmental factors and physiological parameters on leaf-scale instantaneous light use efficiency (LUEi) of kiwifruit
可见植物的光合作用是一个对生理环境因子敏感复杂的过程,生理环境因子不仅直接影响光合作用,而且还通过相互间的影响进而影响光合作用,各种因子之间有着错综复杂的关系.
2.2 冠层和生态系统尺度光利用效率变化规律
2018—2020年猕猴桃生育期eLUE与PAR月平均日内变化如图2所示,PAR在日内呈现出规则的单峰变化,在日出和日落时值较低,在正午时达到峰值;而eLUE日内变化呈U形,在日出和日落时值较高,在正午达到最小值.文中第2.1节基于叶尺度LUEi的研究也表明LUE随着PAR增加而降低,其原因可能为叶片光合和蒸腾随着PAR增加而增大,但当PAR达到一定程度后,叶片为了减少水分散失降低气孔开度,限制了大气与叶肉细胞间CO2气体的交换,叶片出现了光饱和现象[5],从而导致在较高PAR下的LUE反而小于较弱PAR下的LUE.
2018—2020年猕猴桃生育期累积cLUE,eLUE,PAR以及APAR月变化如图3所示,月累积cLUE与eLUE变化趋势基本一致,cLUE总大于eLUE,但两者均未呈现明显的季节性变化规律.2018—2020年生育期月累积cLUE分别为0.97~1.83,0.77~2.67和1.01~1.74 g/MJ,月累积eLUE分别为0.69~1.63,0.58~1.54和0.81~1.40 g/MJ.月累积PAR和APAR呈现明显的季节性变化,2018—2019年8月PAR和APAR值最大,分别为211.70和196.99 MJ/(m2·月),225.80和209.53 MJ/(m2·月);2020年5月,PAR和APAR值最大,分别为223.01和195.06 MJ/(m2·月).

注:图中PAR和eLUE 为每0.5 h变化(0.5 h尺度)
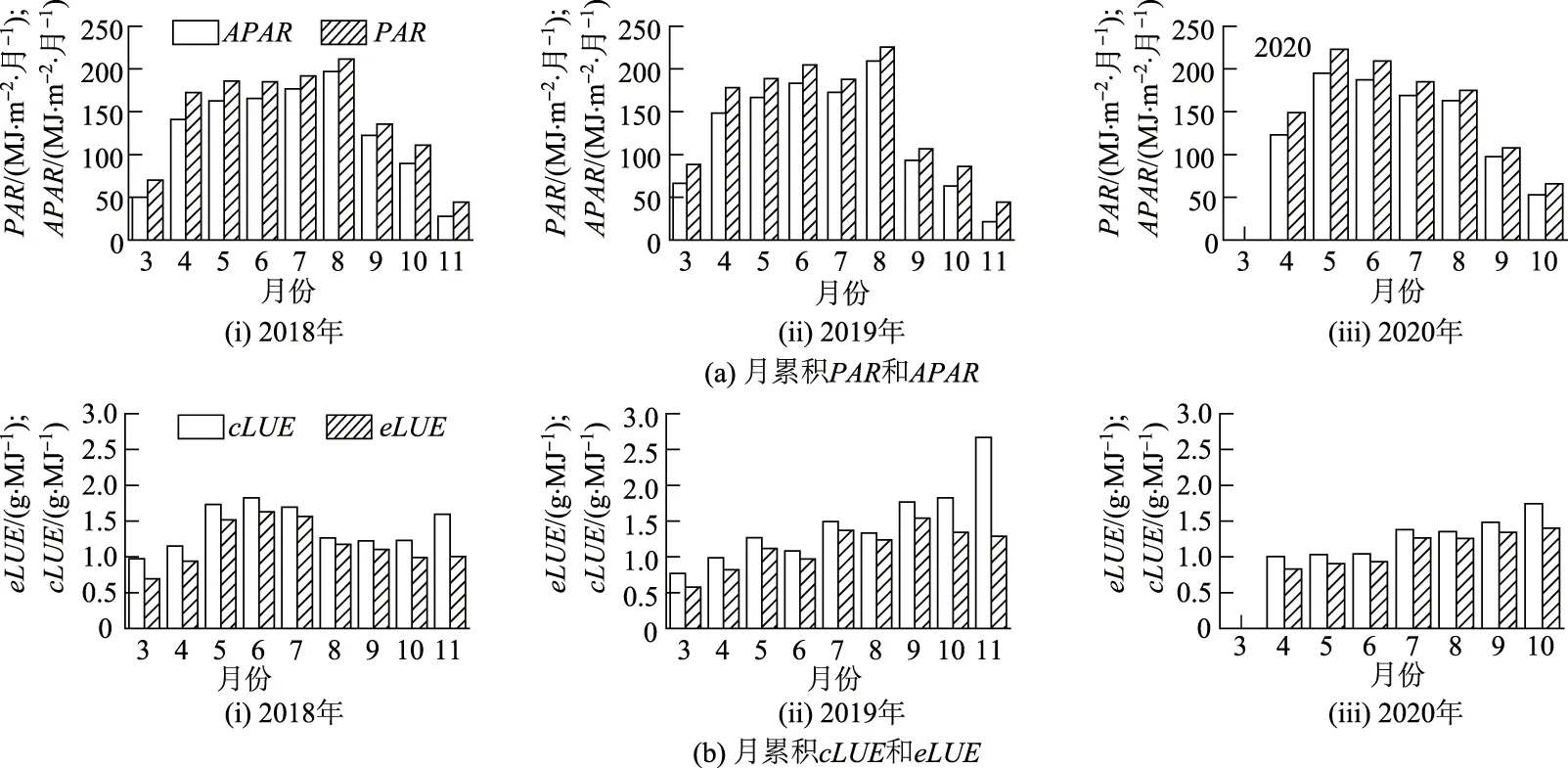
注:图中PAR,APAR,cLUE和eLUE 为每月变化(月尺度)
2.3 冠层和生态系统尺度光利用效率驱动因素
表2为2018—2020年猕猴桃生育期内气象、土壤水分和植被因子对cLUE和eLUE的总影响,表中At为总影响量化值.

表2 猕猴桃果园气象、土壤水分和植被因子与冠层光利用效率cLUE和生态系统光利用效率eLUE的相关性Tab.2 Correlation between weather, soil moisture and vegetation factors in kiwi orchard and canopy light use efficiency and ecosystem light use efficiency
图4为气象、土壤水分和植被因子对cLUE和eLUE的直接和间接影响,图中Ad为直接影响量化值,Ai为间接影响量化值.

图4 猕猴桃果园环境因子对冠层光利用效率cLUE和生态系统光利用效率eLUE的直接和间接影响Fig.4 Direct and indirect effects of environmental factors of kiwi orchard on canopy light use efficiency and ecosystem light use efficiency
通径分析结果表明:Rg对cLUE具有显著的总影响(P<0.01),量化值为-0.60~-0.42,并主要来自于其直接影响;Ta,VPD及冠层叶面积指数(LAIc)对cLUE有显著直接和间接的影响,但直接和间接的影响相互抵消,最终没有表现出对cLUE有显著的总影响.Rg和下垫面总叶面积指数(LAIt)对eLUE具有显著的总影响,其中Rg对eLUE的总影响(量化值为-0.58~-0.42)主要来自于其直接的影响,而LAIt对eLUE的总影响(量化值为0.34~0.45)主要来自于其通过其他因子对eLUE的间接影响;Ta和CH2O对eLUE有显著直接和间接的影响,但直接的影响与其通过其他因子对eLUE的间接影响抵消,最终没有表现出对eLUE具有显著的总影响.
图5为2018—2020年猕猴桃生育期内气象、土壤水分和植被因子通过APAR(PAR)和GPP对cLUE和eLUE的间接影响.

图5 猕猴桃果园环境因子通过光合有效辐射PAR、吸收的PAR(APAR)和总初级生产力GPP对冠层光利用效率cLUE和生态系统光利用效率eLUE的间接影响Fig.5 Indirect effect of environmental factors of kiwi orchard on canopy light use efficiency and ecosystem light use efficiency through photosynthetically active radiation, absorbed PAR and total primary productivity
为了提高气象、土壤水分和植被因子对cLUE和eLUE影响的解释率,在通径分析中加入了PAR(APAR)和GPP.通径分析结果:气象因子中Rg,Ta和VPD通过APAR(PAR)和GPP对cLUE(eLUE)的影响较大(影响量化值超过0.50);Rg,VPD和Ta通过GPP对cLUE的影响量化值分别为0.81,0.77和0.82,而通过APAR对cLUE的影响量化值分别为-1.11,-0.87和-0.78;Rg,VPD和Ta通过PAR对eLUE的影响量化值分别为-1.25,-0.94和-0.76,而通过GPP对eLUE的影响量化值分别为0.82,0.78和0.84.说明Rg和VPD通过APAR(PAR)对cLUE(eLUE)的负影响大于通过GPP对cLUE(eLUE)的正影响,Ta则是正影响大于负影响,表明Rg和VPD主要是通过影响光吸收过程影响cLUE和eLUE,而Ta主要是通过影响碳同化过程(GPP)影响cLUE和eLUE.另外,冠层叶面积指数(LAIc)通过APAR和GPP对cLUE的影响量化值分别为-0.59和0.69,下垫面总叶面积指数(LAIt)通过PAR和GPP对eLUE的影响量化值分别为-0.52和0.70,表明植被因子主要通过影响碳同化过程(GPP)影响cLUE和eLUE.
综上结果表明,PAR和GPP主要是通过直接途径影响LUE,而生理因子、气象因子和植被因子主要通过影响PAR和GPP以间接途径控制LUE.结果可以帮助我们更好地理解生理、环境和植被因子对LUE的交互影响.
3 讨 论
LUE动态变化对于生态系统生产力估计和碳循环研究具有重要意义.文中研究表明,猕猴桃生育期PAR在日内呈现出规则的单峰变化,而eLUE日内变化呈U形,表明猕猴桃果园生态系统日出和日落时的光合作用效率更高,中午时较小,原因可能是中午的温度和光强太高,导致猕猴桃果园生态系统光合出现了光抑制现象,而在日出和日落时温度和光强较低,光合作用效率反而更高,这与前人的研究结果类似[18].文中研究还表明,猕猴桃生育期内cLUE与eLUE未呈现明显的季节性变化,且两者的变化趋势基本一致.朱亚平[19]通过对比分析长白山温带针阔混交林(CBM)和鼎湖山亚热带常绿阔叶林(DHM) 2种地带性森林生态系统LUE的季节变异特征,研究表明CBM的LUE随温度的季节变化表现出明显的单峰变化形式,而DHM由于全年水热条件均适宜植被生长,其LUE的季节动态特征并不明显,这与文中研究结果相近.仇宽彪等[20]发现沙地杨树人工林生态系统LUE存在明显的季节变化趋势,这与文中研究发现不一致.这可能是由于在猕猴桃整个生育期下垫面总叶面积指数LAIt较高,相对于干旱区LAI季节性变化不明显,导致LUE没有呈现出明显的季节性变化;此外,猕猴桃果园冠层光截获率较大,PAR和APAR比较接近,导致GPP/PAR与GPP/APAR的比值较为接近,所以cLUE与eLUE的变化趋势基本一致.
LUE受不同的环境因子影响,例如辐射、土壤水分、空气温度Ta、饱和水汽压差VPD以及叶面积指数LAI等,都可以影响作物的生理生化过程[18, 21].文中研究表明,Ta和LAI主要是通过碳同化过程影响eLUE和cLUE,Rg和VPD主要是通过光吸收过程影响eLUE和cLUE.BROWN等[22]研究发现,在适宜温度时,紫花苜蓿的LUE随温度升高而增大;在较高温度时,蓖麻的LUE随温度升高而降低;温度过低时,豌豆的LUE会降低.这主要是温度较高时呼吸作用增强使光合产物损失增加,温度过低时叶片中光合色素含量较低,这都会显著影响叶片碳同化过程,引起LUE降低.JIANG等[17]研究不同晴空条件对猕猴桃eLUE的影响后发现,直射光利用效率(GPP/PARdir)随直射光合有效辐射PARdir增加呈指数下降,而散射光利用效率(GPP/PARdif)随散射光合有效辐射PARdif没有显著变化,这是由于PARdir只能照射猕猴桃冠层上部的部分叶子,这些叶片在晴朗天气很容易达到光饱和,但冠层内部大多数叶片由于受上部叶片的阻挡,直射光无法穿透而处于光照不足状态,最终表现为eLUE随光照强度增强而下降.因此,可以考虑在作物营养生长旺盛期时进行适当修剪,通过减小冠层叶面积的方式提高作物光利用效率,进而促使作物由营养生长更多地转变为生殖生长,以达到提高作物产量的目的.此外,土壤水分和养分影响着作物的生长,与作物光合作用密切相关.文中研究表明,土壤水分对LUE的影响不具有统计学意义,这主要是由于研究区属于湿润区,土壤水分基本不对光合作用产生限制.唐凡等[23]研究发现在果实膨大期适度的水分亏缺和钾肥亏缺处理有益猕猴桃对光合的响应,能够提高猕猴桃光饱和点和最大净光合速率,进而提高猕猴桃光利用效率.因此,可以考虑优化水肥管理模式,通过增大作物最大净光合速率,以提高作物光利用效率.
4 结 论
基于中国西南猕猴桃果园连续3 a的水热碳通量数据和气象数据,研究了猕猴桃叶片尺度、冠层尺度和生态系统尺度光利用效率随时间的变化特征及其控制因素.主要结论如下:
1) 猕猴桃LUEi随PAR增大而降低,cLUE和eLUE月均日内变化均呈U形,且在日出和日落时值较高,在正午达到最小值;月累积cLUE与eLUE变化幅度分别为0.77~2.67和0.58~1.63 g/MJ, 两者变化趋势基本一致且未呈现明显的季节性变化规律.
2)PAR和GPP主要是通过直接途径影响LUE,而生理因子、气象因子和植被因子主要通过影响PAR和GPP以间接途径控制LUE.其中PAR和Pn对LUEi分别具有显著的直接影响(量化值为-0.99)和间接影响(量化值为-0.81);气象因子(Rg,Ta,VPD)和植被因子(LAI)对cLUE和eLUE有显著影响,Rg和VPD主要通过影响光吸收过程影响eLUE和cLUE,而Ta和LAI则是通过碳同化过程影响eLUE和cLUE.
研究结果可以更好地理解生理、环境和植被因子对LUE的交互影响,例如可以考虑在作物营养生长旺盛期时进行适当的修剪,通过减小冠层叶面积的方式提高作物光利用效率;或者优化作物特定时期水肥管理模式,通过增大作物最大净光合速率来提高作物光利用效率,进而促使作物由营养生长更多的转变为生殖生长,以达到提高作物产量的目的.
猜你喜欢
农业工程学报(2022年8期)2022-08-08
作物学报(2022年10期)2022-07-21
作物杂志(2022年3期)2022-07-06
中国农业信息(2022年1期)2022-05-25
河南水利与南水北调(2022年1期)2022-03-16
农业科技通讯(2022年2期)2022-02-28
农业机械学报(2021年11期)2021-12-07
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
新疆农业科学(2020年1期)2020-02-14
干旱地区农业研究(2019年6期)2019-02-25
