
沙地不同种类沙生植物根系呼吸对土壤呼吸的贡献
2022-09-21 14:00杨衍龙陈银萍姚彩萍
兰州交通大学学报 2022年3期
杨衍龙,陈银萍,姚彩萍,郑 彦
(兰州交通大学 环境与市政工程学院,兰州 730070)
根据政府间气候变化专门委员会的第六次报告,较1850~1900年,2010~2019年全球平均表面气温的升高幅度为0.9~1.2℃,而气温升高与大气二氧化碳(CO2)浓度升高之间存在密切的正相关性[1].全球土壤碳库量为1 300~20 009 pgC,占全球碳总储存量的67%,土壤碳库的微小波动都会显著改变大气中CO2浓度,从而引起气候变化[2].土壤呼吸是全球陆地生态系统碳循环的关键过程之一,也是全球气候变化的关键生态过程.土壤呼吸是大气CO2重要来源之一,其通量主要受非生物因素和生物因素的影响,非生物因素主要有土壤温度、土壤含水量、有机质等,生物因素主要有地表植被、土壤微生物、土壤动物活动等[3].
目前,土壤呼吸的相关研究主要集中在森林、湿地、草地、农田、城市绿地等[4]生态系统,而针对沙地生态系统的研究相对较少[5-6],尤其对于沙生植物根系呼吸对土壤总呼吸贡献的研究更是匮乏.干旱半干旱地区约占世界陆地面积的三分之一,土壤呼吸是该区域土壤碳损失的主要过程之一[7].科尔沁沙地是中国典型的北方农牧交错区和生态脆弱区,脆弱的生态基质在气候变化和人类活动双重影响下,沙漠化发展严重,致使原生植被遭到破坏,生境不断恶化[8].研究表明,沙漠化的生态恢复具有较高的固碳潜力,因此有关该区域生态系统演变与固碳过程的研究近年来引起了广泛关注[9-10].
刘鹏等[11]对毛乌素沙地油蒿灌丛土壤呼吸的研究发现,根系呼吸对土壤总呼吸的贡献率为40%~70%.Zhou等[12]在相关研究中发现,沙漠地区土壤呼吸强度随根系生物量的增加而提高.土壤呼吸除了受上述因素的影响外,还受到时间尺度变化的影响,表现出明显的季节变化和日夜变化特征[13].本研究选取科尔沁地区固定沙丘和流动沙丘风沙土,分别人工种植狗尾草、差巴嘎蒿、沙米和扁蓿豆四种沙生植物,进行同等条件管理,测定其48 h土壤呼吸速率动态,分析土壤呼吸组分与植物种类、根系生物量、土壤属性及环境温湿度等影响因素的关系,探究不同植物根系呼吸对土壤呼吸的贡献,以期为该区域碳平衡的深入研究提供基础数据.
1 材料与方法
1.1 研究地区概况
研究区位于科尔沁沙地中南部奈曼旗境内,本实验依托中国科学院奈曼沙漠化研究站(42°55′N,120°42′E,海拔345 m)开展.研究区属于温带半干旱大陆性气候,年平均气温5.5~6.5℃,≥10℃的积温为3 000~3 300℃,无霜期140~160天,年均降水量343~451 mm,年蒸发量1 600~2 700 mm,70%~80%的降水集中在夏季,年平均风速3.5~4.5 m/s.研究区地带性的土壤类型主要为栗钙土,现多已退化为风沙土.研究区典型的固定沙丘和流动沙丘表层(0~20 cm)土壤有机碳和养分含量见表1.主要优势植物有猪毛菜(Salsolacollina)、差巴嘎蒿(Artemisia halodendron)、沙米(Agriophyllum squarrosum)、扁蓿豆(Melissilusruthenicus)、虫实(Corispermumhyssopifolium)、防风(Saposhnikoviadivaricata)和狗尾草(Setariaviridis)等.
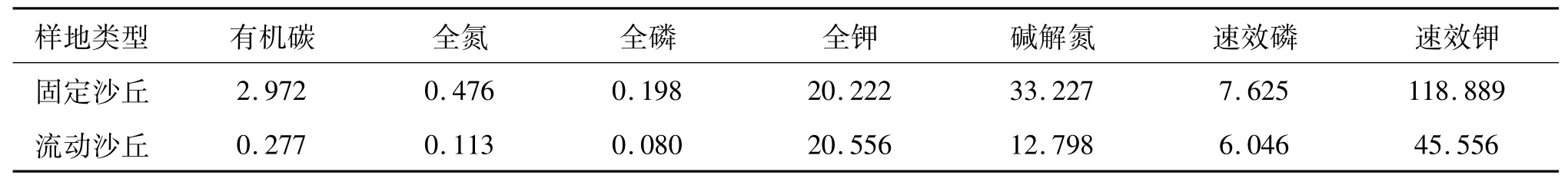
表1 两类沙丘土壤有机碳和养分质量分数特征Tab.1 Characteristics of organic carbon and nutrients content in two sandy dune types g/kg
1.2 研究方法
选取研究区具有代表性的固定沙丘和流动沙丘,分别随机建立5个1 m×1 m的样方,挖取表层0~20 cm的土壤,带回实验室后过2 mm筛后,按照沙丘类型分别将5个样方的土壤混合均匀,然后装入PVC管(内径10 cm,高33 cm,底端用透气膜封闭,风沙土填至30 cm处),每一类型土壤装填20个管,5种实验处理(栽植4种植物+1个不栽植物的空白对照),每种处理4个重复.选取狗尾草、差巴嘎蒿、沙米和扁蓿豆4种植物的当年生幼苗,每种植物的幼苗大小尽可能一致,初始栽入每个PVC管中的相应植物均为2株,将其埋入户外沙地(PVC管露出地表3 cm).栽后前10天视降雨情况不定期浇水,土壤水分保持在田间持水量的70%左右.等植物稳定成活后,每管保留1株植物,其后不再人工浇水,水分来源于自然降雨.生长30天后,齐地表剪去植物地上部分,测定48 h的土壤呼吸动态,测定频率白天每2 h一次,夜间每3 h一次.
测定仪器为美国Li-cor公司生产的LI-6400-09土壤呼吸测定系统测定,测定土壤呼吸的同时,用仪器自带的温度探针测定土壤表层10 cm的温度.在土壤呼吸测定结束后立即挖出PVC管,分0~10 cm和10~30 cm两层测定根系生物量.与此同时取0~10 cm土壤样品,用烘干法测定土壤含水量,用氯仿熏蒸浸提法测土壤微生物量碳.
1.3 数据处理
不栽植物的空白对照(CK)测定值即为土壤异养呼吸(Rm),栽植植物的测定值即为土壤总呼吸(RS).RS与Rm之间的差值可认为是根系呼吸(Rb).利用SPSS22.0对所得数据进行方差、标准误差和相关性分析,采用Orign2018作图.
2 结果与分析
2.1 不同土壤条件下土壤呼吸日变化特征
不同实验处理下的土壤呼吸日动态变化如图1所示.土壤温度的测定结果为图1(a),固定沙丘土壤温度在18.9℃~34.4℃范围内波动,温差为15.5℃.流动沙丘土壤温度在17.8℃~34.0℃范围波动,温差为16.1℃.固定沙丘土壤平均温度为25.1℃,流动沙丘土壤平均温度为25.2℃,二者温差为0.1℃.两种土壤条件下,土壤温度昼夜变化呈现“多峰型”,峰值总体出现在11∶00~17∶00,最低值出现在22∶00~4∶00.两种沙丘土壤人工种植植物后土壤呼吸均显著高于对照,表现为狗尾草>差巴嘎蒿>扁蓿豆>沙米>CK,前24 h与后24 h土壤呼吸变化趋势相同,呈现“多峰型”.四种植物土壤呼吸峰值总体出现在11∶00~17∶00,最低值出现在4∶00~7∶00.同一植物,固定沙丘中土壤呼吸速率动态变化整体大于流动沙丘,差巴嘎蒿表现则相反.沙米、狗尾草和扁蓿豆在固定沙丘土壤呼吸速大于流动沙丘(见图1(b)、1(c)、1(d)).差巴嘎蒿则在流动沙丘中土壤呼吸速率峰值大于固定沙丘,分别为6.583μmol·m-2·s-1和4.446μmol·m-2·s-1(见图1(e)).
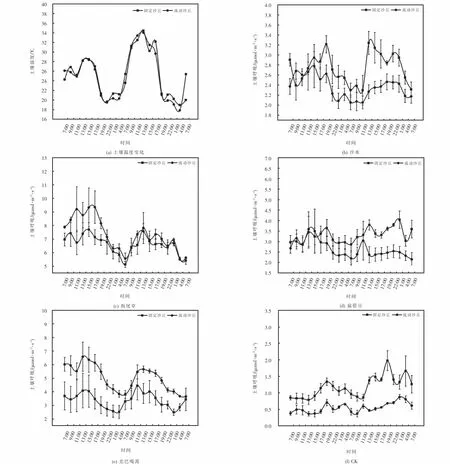
图1 不同实验处理下的土壤呼吸状态Fig.1 Soil respiration under different experimental treatments
2.2 不同植物根系呼吸对土壤呼吸的贡献率
四种植物根系呼吸对土壤呼吸的贡献率不同(见图2),如图2所示,固定沙丘中植物根系呼吸贡献率分别为:狗尾草82.33%、差巴嘎蒿65.23%、扁蓿豆64.59%、沙米57.16%;流动沙丘中植物根系呼吸贡献率分别为:狗尾草92.50%、差巴嘎蒿89.07%、扁蓿豆79.33%、沙米76.83%;四种植物分别作用于固定沙丘和流动沙丘时,狗尾草对土壤呼吸的贡献率显著高于沙米和扁蓿豆,但沙米和扁蓿豆根系呼吸贡献率相比无显著差异;四种不同植物作用下,同种植物根系呼吸贡献率表现为在流动沙丘中高于固定沙丘,两种土壤条件下植物根系呼吸贡献率由高到低均表现为狗尾草>差巴嘎蒿>扁蓿豆>沙米.

图2 植物根系呼吸对土壤呼吸的贡献率Fig.2 Contribution of the plant root respiration to soil respiration
2.3 不同土壤条件下根系生物量、微生物量碳及土壤含水量的变化
根系生物量测定结果如图3所示.两种土壤条件两个土层根系生物量由高到低依次均为:狗尾草>差巴嘎蒿>扁蓿豆>沙米.固定沙丘中,狗尾草0~10 cm土壤深处根系生物量为6.113 g,10~30 cm土壤深处根系生物量3.178 g,二者比值为1.924.差巴嘎蒿0~10 cm土壤深处根系生物量为5.490 g,10~30 cm土壤深处根系生物量1.983 g,比值为2.768.扁蓿豆和沙米不同土壤深处根系生物量比值分别为1.598和2.424.流动沙丘0~10 cm土壤深处根系生物量分别为14.0 g、5.248 g、1.728 g和1.003 g;10~30 cm土壤深处根系生物量分别为9.243 g、1.670 g、0.833 g和0.368 g,不同深度根系生物量的比值分别为:1.515、3.143、1.034和2.727.
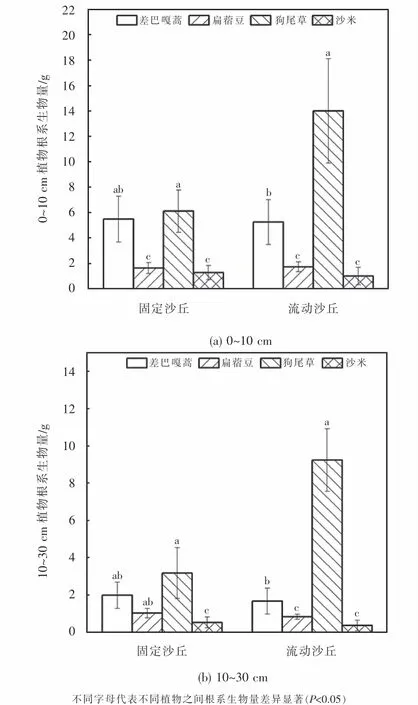
图3 两类沙丘土壤的根系生物量Fig.3 Root biomass in two sand dune types
土壤微生物量碳测定结果如图4(a)所示.两种土壤条件下,种植植物后显著提高了土壤微生物量碳含量(见图4(a)),在固定沙丘中依次为:差巴嘎蒿>扁蓿豆>狗尾草>沙米>CK;在流动沙丘依次为:扁蓿豆>狗尾草>差巴嘎蒿>沙米>CK.土壤含水量测定结果如图4(b)所示.流动沙丘土壤含水量均低于固定沙丘,植物种植后土壤含水量较对照组均显著降低,种植狗尾草使土壤含水量降幅最大,固定沙丘和流动沙丘土壤含水率分别为1.8%和0.38%.
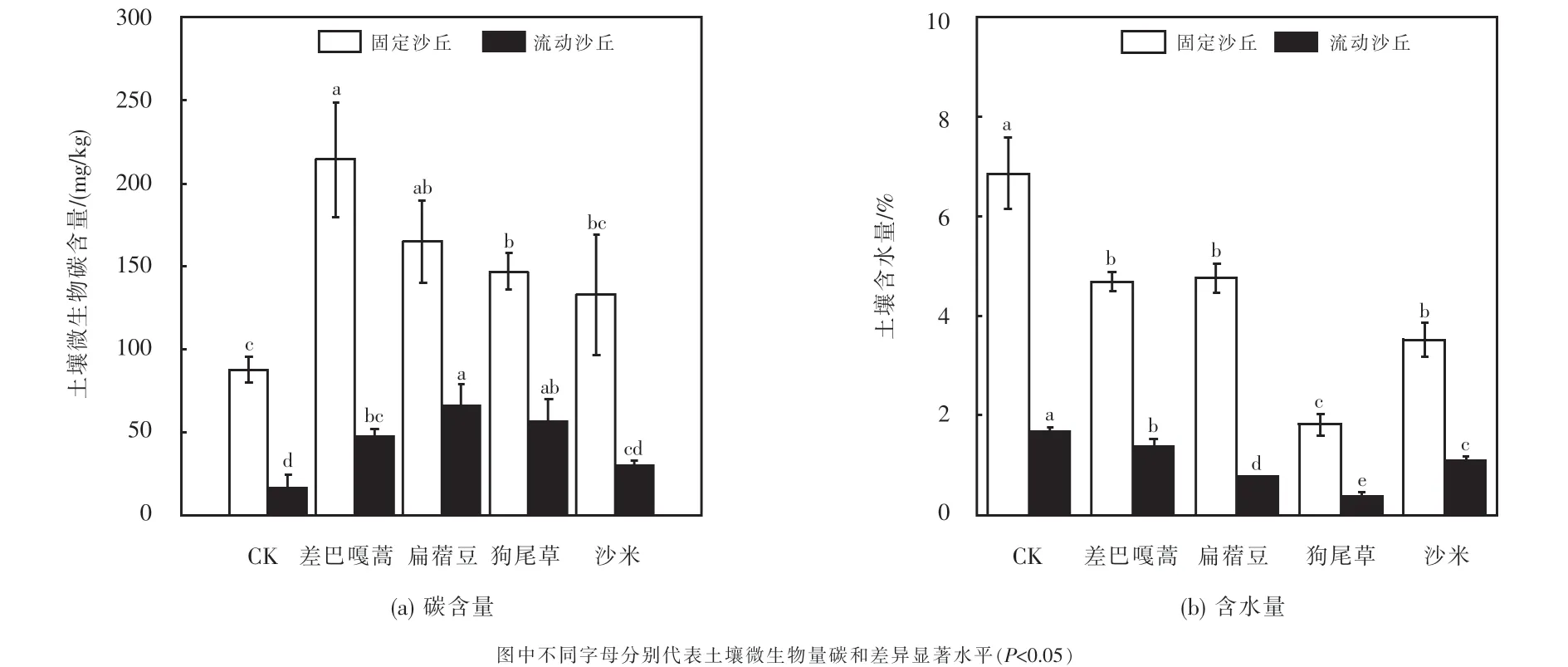
图4 土壤微生物碳含量和土壤含水量测定结果Fig.4 Soil microbial biomass carbon and soil water content among planted species
2.4 不同土壤条件下土壤呼吸与影响因子的相关性分析
不同土壤条件下土壤呼吸各因素综合相关性分析结果如表2所列.从相关性来看,不同土壤条件下土壤呼吸与土壤温度相关性不显著(P>0.05),与土壤含水量呈极显著负相关(P<0.01),与根系生物量呈极显著正相关(P<0.01),土壤呼吸与根系生物量的相关性随土壤深度的增加而降低.固定沙丘中土壤呼吸与土壤微生物量碳相关性不显著(P>0.05),流动沙丘中土壤呼吸与土壤微生物量碳呈显著正相关(P<0.05).

表2 土壤呼吸与各因素相关性分析Tab.2 Correlation between soil respiration and various factors
2.5 植物类型和沙丘类型对土壤呼吸的交互效应分析
植物类型和沙丘类型对土壤呼吸的交互效应如表3所列.植物类型对土壤呼吸影响差异极显著(P<0.01),沙丘类型对土壤呼吸影响无显著差异(P>0.05),植物类型和沙丘类型对土壤呼吸存在极显著的交互效应(P<0.01).各因素对土壤呼吸变异的贡献是植物类型>植物类型和沙丘类型交互效应.

表3 植物类型和沙丘类型对土壤呼吸的交互效应分析Tab.3 Interaction effect of plant species and soil types on soil respiration
3 讨论
3.1 不同土壤条件下土壤呼吸动态变化
土壤呼吸不仅受土壤温度、土壤水分和土壤养分等非生物因素的影响,而且受地面植被和土壤微生物等生物因素的影响[14].影响土壤呼吸的诸多因素在不同时间尺度上的变化不尽相同,如空气温度与湿度、植物根系活动等具有随季节变化和日变化特征.这些因素在时间尺度上的变化可能会使土壤呼吸的季节间变化和昼夜间变化产生差异[15].因此在不同时间尺度上,土壤呼吸的动态变化可能不同,不同生态系统的土壤呼吸的季节性变化也会不同.本实验不同土壤条件下,四种植物土壤呼吸均高于空白对照组,总体上从7∶00起土壤呼吸速率逐渐升高,17∶00起土壤呼吸速率逐渐降低,呈“多峰型”昼夜变化,这一变化的主要原因从7∶00起由于太阳照射的原因,土壤温度明显升高,相应的土壤呼吸速率升高,此时间段由于地面红外辐射导致土壤温度降低,土壤呼吸速率也随之降低.土壤呼吸速率的增长与植物生理活动、根系生物量及土壤微生物的活性紧密相关.土壤温度通过改变植物地上部分的生理活动、地下根系呼吸和土壤微生物呼吸等方式来影响土壤呼吸[16].土壤温度随时间的变化,造成植物根系呼吸、微生物活性等随时间变化,从而引起土壤呼吸呈现明显的日变化特征.多数研究表明土壤呼吸与土壤温度有显著相关性,土壤温度使得土壤呼吸呈现日、月、季、年的变化特征[17].也有研究表明沙地生态系统中,土壤呼吸与土壤温度没有显著的相关性,土壤温度和其他因素对土壤呼吸起共同作用,影响错综复杂,关系尚不明确,但土壤呼吸呈现日、季变化特征[2].
3.2 不同植物根系呼吸对土壤呼吸的影响及贡献率
本次实验选取的四种不同植物对土壤呼吸速率有显著影响,四种植物之间存在显著的差异性.造成差异的主要原因是不同的植物类型构成不同的生态环境,影响土壤理化性质并改善土壤肥力,已有研究表明不同植被类型的土壤养分含量具有明显差异[18].马立群等研究发现在差巴嘎蒿的覆盖后土壤呼吸发生了显著变化,这与本研究结果一致.不同植被类型对地表覆盖和土壤特性的影响不同,使植物根系生长和土壤生物活动发生改变,进而影响土壤呼吸.差巴嘎蒿根系的水平分布呈现以根基为中心向外辐射的趋势,主要以直径1~2 mm的根系为主,呈“伞”型分布,在1~10 cm土层中根系发达[19];沙米根系具有浅根性,主根短小,侧根发达,向四周延伸,多分布于沙表层;狗尾草根系主要生长在0~30 cm的土层内,是比较典型的浅层根系植物;扁蓿豆为轴根性植物,根系入土深度达60~140 cm,其中有80%左右的根系分布在0~30 cm范围内[20-21].不同植被类型根系的生长过程中,根系对土壤水分、养分的需求不同,根系微生物群落也有差异,最终导致土壤呼吸的差异,这与王忆慧等[22]的结果研究一致.现有的研究表明,根系呼吸对土壤呼吸贡献率可达30%~70%[23],贡献率也因区域气候和生态环境的不同而存在较大差异,北方干旱和半干旱区域根系呼吸贡献率较高,为50%~93%[24],与本研究植物根系对土壤呼吸的贡献率结果相符.
3.3 根系生物量、土壤微生物、土壤含水量对土壤呼吸的影响
不同沙生植物条件下根系生物量和土壤水分含量存在显著差异.根系生物量是影响土壤呼吸的重要因素之一,研究发现土壤微生物群落参与土壤养分循环和物质代谢过程,通过降解植物凋落物和改善土壤理化性质等方式影响植物生长[25],根际微生物生长和活动强烈,养分循环速度快,因此土壤中微生物呼吸的主要来源是根际微生物的呼吸[26].温度和降水通过调控土壤养分供给,微生物群落数量及活性来影响土壤微生物呼吸[27].种植不同植物的土壤呼吸受根系微生物群落的影响不同,由土壤温度、养分、水分及微生物群落多样性等诸多因素决定.
土壤水分也是影响土壤呼吸的重要因素之一,研究发现土壤水分通过影响根系呼吸和微生物呼吸来改变土壤呼吸,当土壤含水量小于田间持水量时,土壤含水量对土壤呼吸起到促进作用,土壤呼吸速率会随着土壤含水量的增加而增加,这是因为随着土壤含水量增加,土壤中的微生物和动物新陈代谢所需要的激发能减少,导致土壤呼吸迅速增加[28-29].崔骁勇等[30]研究发现,当水分过少时,土壤根系呼吸和微生物呼吸将受到抑制,也会减少土壤呼吸通量,与本实验流动沙丘土壤含水量和根系生物量对土壤呼吸的影响相一致.
3.4 不同土壤条件下土壤呼吸与影响因子的相关性
本研究中土壤呼吸与土壤温度无相关性,与土壤含水量极显著负相关.多数控制性实验证明,土壤呼吸通常对首要限制它的因素产生响应[30].在受水分胁迫的干旱、半干旱地区,土壤含水量较低,日变化不明显,对植被根系生长和微生物活动产生限制,土壤水分是土壤呼吸最主要的胁迫因子.研究表明,科尔沁沙丘生境短时间的温度变化对土壤呼吸没有显著的影响[2].本研究中土壤呼吸与土壤温度相关性不显著,主要原因可能是本实验中土壤水分在实验前后的变化且土壤呼吸测定时间尺度较短.土壤含水量对土壤呼吸的影响表现为时间尺度上的特征,并与温度等其他因素有密切关系,过高和过低的水分状况都会限制温度对土壤呼吸的作用,且土壤含水量对土壤呼吸的限制随着温度的升高而增强[31].土壤含水量与土壤呼吸大多呈正相关,但也有负相关的报道,Dorr等[32]的相关研究结果发现土壤含水量与土壤呼吸呈负相关,与本研究结果相一致.马立群等[33]研究了差巴嘎蒿群落对土壤呼吸的影响,土壤呼吸与土壤优势微生物种群受到差巴嘎蒿的影响,进而影响土壤呼吸.本研究中不同植物根系生物量存在显著差异,且与土壤呼吸显著正相关,这与上述植物根系对土壤呼吸影响的研究一致.本研究中土壤呼吸与微生物量碳在固定沙丘中无相关性,在流动沙丘中显著相关,主要原因是固定沙丘中有机质相对较为丰富,土壤环境相对稳定,微生物群落结构稳定,且活性较高,即便较少的微生物量也能够快速高效的进行有机质转化.相反,在流动沙丘中有机质薄弱,土壤环境稳定性较差,土壤微生物活性较低,土壤微生物量直接影响土壤呼吸通量.此外,本研究中植物类型和土壤类型对土壤呼吸产生交互作用,因此,科尔沁沙地土壤呼吸受土壤温度、土壤含水量、土壤微生物量、根系生物量等多重因素共同影响,各个影响因子之间相互促进、相互制约.
4 结论
1)人工栽植植物后土壤总呼吸显著高于无植物的土壤,但不同植物间的土壤呼吸存在差异,两种沙丘土壤条件下均表现为:狗尾草>差巴嘎蒿>扁蓿豆>沙米.
2)四种不同植物作用下,同种植物根系呼吸贡献率表现为流动沙丘中高于固定沙丘,两种沙丘中植物根系呼吸贡献率可以达到57.16%~92.50%,由高到低依次表现为狗尾草>差巴嘎蒿>扁蓿豆>沙米.
3)四种不同植物作用下,土壤呼吸与土壤温度相关性不显著(P>0.05),与土壤含水量呈极显著负相关(P<0.01),与根系生物量呈极显著正相关(P<0.01);在固定沙丘中土壤呼吸与土壤微生物量碳相关性不显著(P>0.05),在流动沙丘中土壤呼吸与土壤微生物量碳呈显著相关(P<0.05).
4)植物类型对土壤呼吸影响差异极显著(P<0.01),土壤类型对土壤呼吸影响无显著差异(P>0.05),植物类型和土壤类型对土壤呼吸存在极显著的交互效应(P<0.01).
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
中国农业气象(2022年8期)2022-08-24
农业工程学报(2022年10期)2022-08-22
现代园艺(2022年6期)2022-04-08
土壤(2022年1期)2022-03-16
作文周刊·小学一年级版(2020年40期)2020-10-19
安徽农学通报(2020年7期)2020-05-26
江苏农业科学(2017年21期)2017-12-13
广东第二课堂·小学(2017年2期)2017-02-20
小学生作文选刊·低年级版(2016年6期)2016-06-24
