
Myo1h基因敲除小鼠模型的建立与初步观察
2022-09-15 02:12温士强孙榕榕李永明
口腔医学 2022年8期
温士强,孙榕榕,时 函,李永明
Ⅰ类肌球蛋白基因(Myo1h)作为非常规型肌球蛋白1H的调控基因,对颌骨的生长发育尤为重要,成为近年来的研究热点[1-2]。首先,Tassopoulou-Fishell等通过研究发现位于12q24.1的Myo1h基因与下颌前突相关,其中rs10850110的G等位基因在下颌前突病例中显著多于对照组,且于不同种族背景中无差异,为Myo1h基因可能有助于下颌前突提供了遗传证据[3]。Cruz等通过主成分分析的研究发现Myo1h(rs10850110 A
目前已知人Myo1h基因第3、25、30号外显子与小鼠存在高度同源性,且有研究表明小鼠Myo1h同源基因第3号、25号外显子的突变能够导致一种罕见的隐性先天性通气不足,并具有明显的致死效应[6],但小鼠Myo1h同源基因对颌骨生长发育有何影响仍未知。而Myo1h基因第30号外显子rs3825393多态位点的突变能够增加汉族人群中下颌后缩的风险[7],也进一步说明了位于Myo1h基因尾域结构的第30号外显子对颅颌面骨生长发育中发挥着重要作用。
研究表明Myo1h基因是颌骨生长发育畸形的易感基因,本课题组前期研究也证实Myo1h基因为下颌前突的易感基因,并发现Myo1h基因可能通过对下颌软骨发育和形态形成产生影响,进而参与下颌前突等颅颌面发育异常的发生。为进一步研究Myo1h基因对哺乳动物颅颌面骨骼发育的影响,构建Myo1h基因敲除实验动物模型十分必要,目前国内外对此部分的研究尚处于起步阶段。基于Cripr/Cas9技术的诸多优势[8-10],本研究采用Cripr/Cas9技术构建Myo1h-/-小鼠模型,为进一步探索Myo1h基因对骨生长发育中的影响,尤其是为探究位于哺乳小鼠Myo1h同源基因尾域结构的第30号外显子对于颅颌面骨生长发育的作用提供研究基础。
1 材料与方法
1.1 实验动物
3~4周龄SPF级C57BL/6JMyo1h基因敲除的嵌合体F0小鼠3只,雄鼠1只雌鼠2只,均由同济大学生科院提供。所有小鼠均饲养在同济大学医学院实验动物中心SPF级动物房,温度控制在25 ℃左右,湿度保持在70%左右,明暗循环12 h/d,自由饮水和进食。小鼠笼盒、垫料、饲料、饮用水均经高温高压消毒灭菌处理。饲养过程中,每2 d进入动物房观察和记录小鼠的生长情况。每3~4 d更换1次小鼠垫料,每天补充饲料和饮用水。所有操作均符合同济大学实验动物伦理学要求(伦理号:TJLAC-019-150)。
1.2 主要仪器与试剂
多聚甲醛(上海碧云天科技有限公司,中国);乙二胺四乙酸钠脱钙液(上海碧云天科技有限公司,中国);琼脂糖粉末(上海博晗生物科技有限公司,中国);TBE粉末(上海碧云天科技有限公司,中国);DNA聚合酶(南京诺唯赞生物科技股份有限公司,中国);DL1000 marker(宝日医生物技术(北京)有限公司,中国);6X DNA凝胶加样缓冲液(宝日医生物技术(北京)有限公司,中国);核酸染色剂Gold View(白鲨生物科技有限公司,中国);PCR逆转录试剂盒(宝日医生物技术(北京)有限公司,中国);荧光定量PCR SYBR Green(翌圣生物科技(上海)股份有限公司,中国);PCR扩增仪(伯乐生命医学产品(上海)有限公司,中国);荧光定量PCR仪(瑞士罗氏公司,瑞士);凝胶成像系统(伯乐生命医学产品(上海)有限公司,中国);电泳仪(天津赛力斯自动化科技有限公司,中国);DAPI染色液(上海碧云天生物技术有限公司,中国);电子天平(赛多利斯科学仪器(北京)有限公司,中国);高速冷冻离心机 (赛默飞世尔科技(中国)有限公司,中国);低速离心机(日本TOMY公司,日本);纯水仪(威立雅水处理技术(上海)有限公司,中国);正置荧光显微镜(Nikon,日本)
1.3 Crispr/Cas9技术构建Myo1h基因敲除小鼠
1.3.1Myo1h基因敲除位点的设计及sgRNA的获取 通过NCBI和Ensembl网站查询小鼠Myo1h基因信息(NCBI Gene ID:231646;Ensembl,Gene:Myo1hENSMUSG00000066952),确定敲除序列片段为第30号外显子后设计向导RNA(gRNA),利用Cas9/gRNA靶点效率检测试剂盒将设计的gRNA靶点体外转录成单向导RNA(sgRNA)并检测转录产物活性,为了防止脱靶效应,分别在30号外显子前后紧邻的内含子上设计获得两对有效的sgRNA,序列如下,sgRNA-1:5′-CGAGGGAGTTTCTCTCGCAG-3′,sgRNA-2:5′-TTACAGTTCTACGTCAGTCC-3′,sgRNA-3:5′-GTGGTAAGTGGTGCGTGCGG-3′,sgRNA-4:5′-AAATCGACCACTCCTAGCCG-3′。
1.3.2 体外获取受精卵 向预先准备好的4周龄左右C57BL/6雌鼠腹腔注射血促性素与人绒毛膜促性腺激素诱导排卵,另备一只8周龄左右C57L/6雄鼠采精子。将在体外获取的精子与卵细胞结合,显微镜下收集受精卵。
1.3.3 显微注射与胚胎移植 通过显微镜注射的方式,将Cas9 mRNA与sgRNA导入到小鼠受精卵中,之后将存活状态良好的受精卵移植到与结扎雄鼠合笼后阴道见栓的代孕雌鼠生殖系统,伤口缝合后饲养于独立通气笼盒环境中。
1.3.4 F0代小鼠获得与后续繁殖策略 代孕雌鼠产下的仔鼠1周龄后剪下脚趾并编号,提取脚趾DNA,经基因型鉴定及测序后筛选出Myo1h突变小鼠F0代嵌合体小鼠。之后利用筛选出的F0(The founder)代Myo1h基因敲除小鼠与野生型小鼠进行繁殖,获得杂合F1(the first filial generation,子1代)代,利用F1代雌雄小鼠进行交配,经过基因型鉴定后获得纯合F2(the second filial generation,子2代)代小鼠(Myo1h-/-)作为实验鼠,并以野生型小鼠(基因型为Myo1h+/+)作为对照。
1.4 Myo1h基因敲除小鼠的基因型鉴定
待F2代仔鼠10 d龄时,将小鼠对应脚趾剪取于经高温高压消毒灭菌的1.5 mL EP管中,在EP管中加入100 μL Buffer A(0.04 mmol/L NaOH溶液),轻轻涡旋混匀。恒温金属浴100 ℃处理30 min,离心(12 000 r/min、5 min),转移上清液至新的EP管中,加入9 μL Buffer B(pH=6.8的Tris-HCl),混匀,4 ℃或-20 ℃放置备用或者直接用于PCR扩增。基因型鉴定引物设计如下,Myo1h正向:5′-AGGGTAGAGAGAGCAGGTGG-3′;Myo1h反向:5′-ATCTTGGCTGGGTTCTGAGC-3′。按PCR说明书Vzyme Mix(10 μL)、引物Myo1h正向(1 μL)、引物Myo1h反向(1 μL)、ddH2O(7.5 μL)、DNA模板(0.5 μL)的体系依次加样,置于PCR反应仪,开启预设程序。配制2%浓度的琼脂糖凝胶,电压设定120 V,时间30 min,电泳鉴定。
1.4.1 肺组织及脑组织的提取 分别取F2代基因型为Myo1h+/+和Myo1h-/-的实验小鼠(n≥3),1%戊巴比妥钠(80 mg/kg)麻醉下,打开胸腔,暴露左心尖,行PBS灌流后,解剖提取肺部组织;解剖颅顶部皮肤及软组织,小心剔除颅骨,提取脑部组织,并置于焦碳酸二乙酯处理水(diethylpyro-carbonate-treated water, DEPC)(上海碧云天生物技术有限公司)预浸泡的RNA free研磨碗中,倒入液氮速冷,研磨棒充分研磨。
1.4.2 实时定量PCR检测Myo1h基因的表达 研磨后的肺及脑组织细粉末加1 mL Trizol,消化1 min,转移至1.5 mL RNA free EP管中,加入200 μL氯仿,摇匀后静置10 min, 4 ℃离心(12 000 r/min,20 min)。小心吸取最上层无色水相液,加入等量异丙醇,摇匀后静置10 min,4 ℃离心(12 000 r/min、20 min)。弃上清,加入适量75%乙醇(DEPC水配制),4 ℃离心(8 000 r/min、5 min),重复洗涤2次弃上清,无香纸上干燥。用10 μL DEPC水充分溶解沉淀,在超微量紫外分光计上机测定RNA浓度,反转录至cDNA。RT-qPCR引物设计如下,Myo1h正向:5′-TTCACATCTCACCAGCGGAC-3′,Myo1h反向:5′-TAACTGGAGGGGTCCATGCT-3′。按qPCR酶5 μL,引物Myo1h正向(0.5 μL),引物Myo1h反向(0.5 μL),cDNA模板(0.5 μL),DEPC水(3.5 μL)的体系依次加样,每样设3个复孔,离心后置荧光定量PCR仪,开启预设程序,分析数据。
1.4.3 免疫印迹(Western Blot,WB)检测Myo1h蛋白的表达 两组蛋白样品在浓度为7.5%的SDS-PAGE胶孔中加样,接通电源,浓缩胶恒定电压80 V电泳 30 min,分离胶恒定电压100 V电泳60 min,恒定电流200 mA湿法转膜120 min,置于5%脱脂牛奶室温封闭2 h,TBST洗膜后,Anti-Myo1h antibody-C-terminal一抗(艾博抗上海贸易有限公司)稀释液4 ℃过夜孵育;TBST洗膜后,加入辣根过氧化物酶标记的驴抗山羊IgG(H+L)二抗(赛默飞世尔科技(中国)有限公司)稀释液室温孵育2 h,TBST洗膜后,运用ECL化学发光试剂盒显影(内参蛋白为β-actin)。
1.4.4 不同组织中Myo1h阳性细胞免疫荧光染色 ①组织处理:实验组和对照组小鼠提取下颌髁突组织后,经4%多聚甲醛固定48 h、10%乙二胺四乙酸钠脱钙21 d、组织脱水、石蜡包埋,以4 μm的厚度进行切片,60 ℃烘烤30 min。实验组和对照组小鼠脑组织与肺组织取样后行常规4%多聚甲醛固定48 h,组织脱水、石蜡包埋,以4 μm的厚度进行切片,60 ℃烘烤30 min。②组织染色:将切片脱蜡、浸水,95 ℃抗原修复,兔抗-Myo1h 多克隆抗体一抗(北京博奥森生物技术有限公司)4 ℃过夜孵育;PBS清洗后,孵育辣根过氧化物酶标记的山羊抗兔IgG(H+L)二抗1 h(上海碧云天生物技术有限公司);PBS 清洗后,滴加DAPI染色液,洗去后封片。在正置荧光显微镜下拍照,随机选取每只小鼠第一磨牙根分叉区域牙槽骨、脑及肺组织3个高倍镜下视野,计数Myo1h阳性细胞数。
1.5 Myo1h基因敲除小鼠的生长发育变化观察
1.5.1 小鼠下颌骨Micro-CT扫描分析 分别取F2代基因型为Myo1h+/+和Myo1h-/-的实验小鼠(n≥3),1%戊巴比妥钠(80 mg/kg)麻醉下解剖提取双侧下颌骨,4%多聚甲醛固定24 h,剔除下颌骨附着的肌肉等软组织,选取合适的装管方向,用泡沫塑料固定,扫描精度10 μm,开机扫描。选取下颌第一磨牙根分叉的颌骨,连续30层作为骨密度分析的感兴趣区(ROI);经三维重建后的下颌骨,用Image J软件测量下颌中切牙与牙槽骨交界处至髁突顶的直线距离,测量3次后取平均值作为有效下颌骨长度。
1.5.2 小鼠有效身长的测量 实验组与对照组分别取4周龄小鼠(n≥3),经1%戊巴比妥钠(80 mg/kg)麻醉,在小鼠全身放松的情况下,以左手食指、中指与无名指轻压小鼠使其全身舒展开仰卧于实验台,用直尺测量小鼠鼻尖部到肛门的距离为身长。
1.6 统计学方法
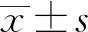
2 结 果
2.1 小鼠培育与基因型鉴定
经繁殖培育的F2代小鼠PCR扩增后,琼脂糖凝胶电泳基因型片段分3种:①扩增长度为430 bp左右,为纯合敲除型(Myo1h-/-)小鼠;②扩增长度为650 bp左右,为野生型(Myo1h+/+)小鼠;③扩增长度介于430 bp和650 bp之间且存在两个电泳条带的为杂合(Myo1h+/-)小鼠。经过琼脂糖凝胶电泳实验鉴定,本研究成功获得Myo1h+/+,Myo1h+/-,Myo1h-/-三种基因型的F2代小鼠(图1 ),其中基因型Myo1h-/-小鼠为实验鼠(图1 箭头所示)。
箭头示基因型Myo1h-/-
2.2 mRNA表达水平比较
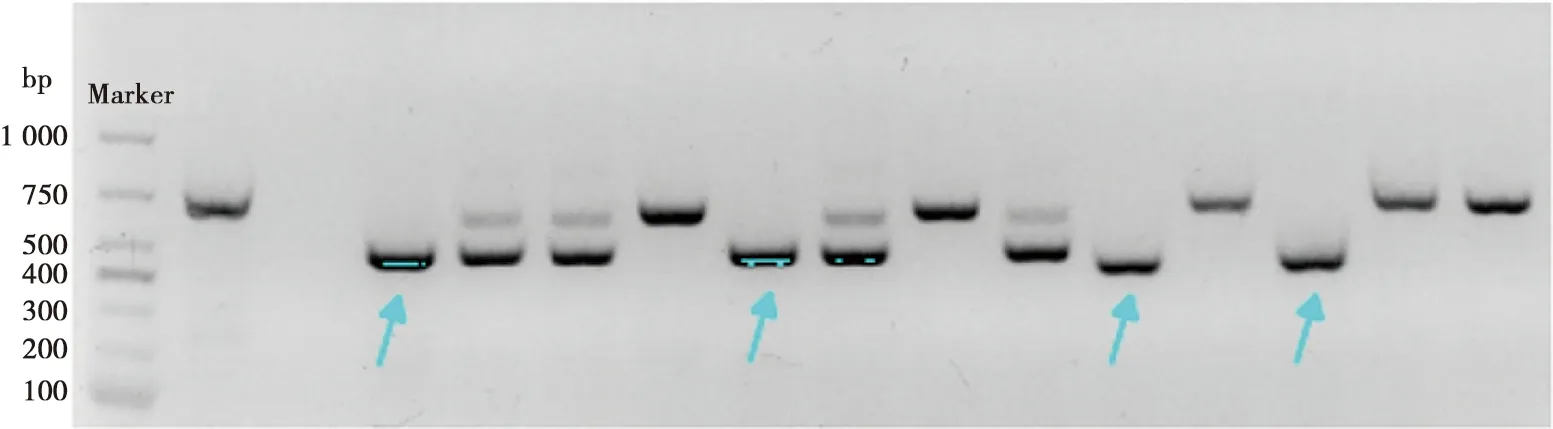
为了进一步确认PCR结果的可靠性,选取经PCR鉴定获得的Myo1h+/+小鼠和Myo1h-/-小鼠,经实时定量PCR的方法检测其大脑及肺部组织Myo1h基因的表达情况,结果如图2示,相较于Myo1h+/+小鼠的正常表达,Myo1h-/-小鼠脑及肺部组织mRNA表达水平显著降低,差异有统计学意义(P<0.05),结果与PCR方法鉴定结果一致,验证了Myo1h基因的敲除效果。
A:脑组织实时定量PCR结果;B:肺组织实时量定PCR结果;**:P<0.01,****:P<0.000 1
2.3 脑组织Myo1h蛋白表达水平对比
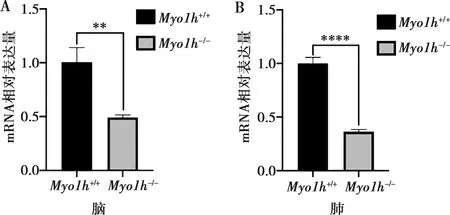
分别提取两种基因型小鼠的脑组织蛋白,Western Blot实验结果显示,在保持β-Actin内参蛋白基本一致的情况下,Myo1h+/+小鼠脑组织的Myo1h蛋白表达正常,而Myo1h-/-小鼠脑组织的Myo1h蛋白表达降低(图3),说明经基因敲除后的小鼠在蛋白水平上可实现Myo1h的敲除。
图 3 Myo1h基因敲除后,Myo1h蛋白表达情况
2.4 敲除后组织中Myo1h基因表达降低
Myo1h+/+小鼠与Myo1h-/-两组小鼠的Myo1h基因免疫荧光染色结果显示基因敲除小鼠髁突软骨层中的Myo1h表达降低(图4 A、D);同时观察到Myo1h基因主要表达在髁突软骨表层及肥大细胞层(图4 A)。Myo1h-/-小鼠脑组织中的Myo1h表达降低(图4 B、E示);Myo1h-/-小鼠肺组织中的Myo1h表达也降低(图4 C、F)。该结果表明,构建的Myo1h基因敲除小鼠的敲除效果实现了体内验证。

A:下颌髁突组织( ×200); B:脑组织( ×200); C:肺组织( ×200);D:髁突组织Myo1h阳性细胞表达比例; E:脑组织Myo1h阳性细胞表达比例;F:肺组织Myo1h阳性细胞表达比例;**:P<0.01,***:P<0.001
2.5 下颌骨Micro-CT骨密度分析
利用Micro-CT对4周龄Myo1h+/+小鼠与Myo1h-/-小鼠右侧第一磨牙根分叉处颌骨扫描分析,并对BV/TV、BS/BV、Tb.N以及Tb.Th等骨密度相关参数进行统计学分析后发现Myo1h基因第30号外显子的敲除并未引起下颌骨密度的改变(图5)。
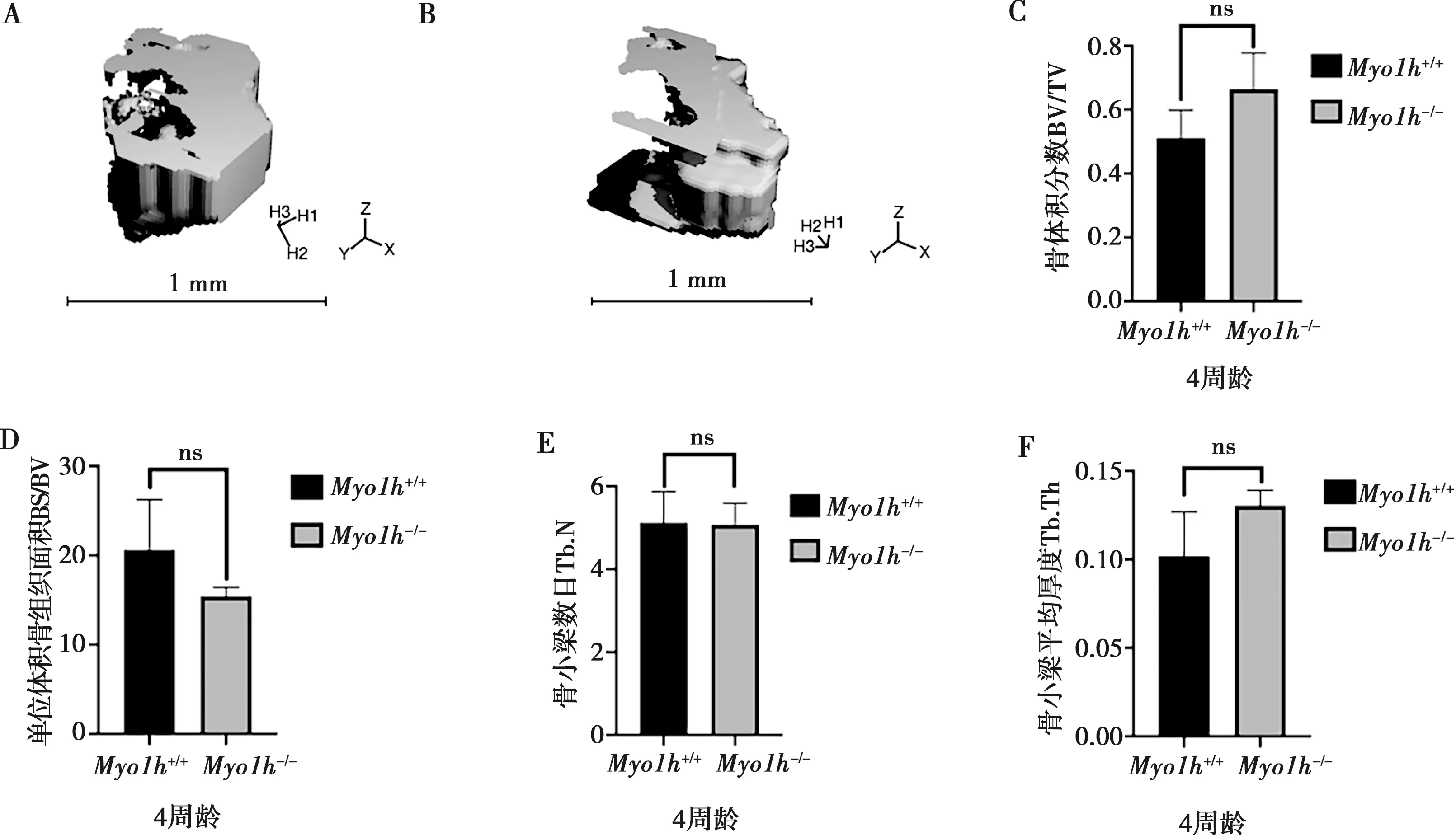
A:Myo1h+/+小鼠下颌骨Micro-CT扫描结果;B:Myo1h-/-小鼠下颌骨Micro-CT扫描结果;C:骨体积分数BV/TV分析;D:单位体积骨组织面积BS/BV分析;E:骨小梁数目Tb.N分析;F:骨小梁平均厚度Tb.Th分析;ns:无显著差异
2.6 有效下颌骨长度及身长的测量
利用Micro-CT对4周龄Myo1h+/+小鼠与Myo1h-/-小鼠右侧下颌骨进行扫描重建,并对下颌骨的有效颌骨长度(EfML)进行测量后发现野生型小鼠有效下颌骨长度为(8.980±0.605)mm,纯合敲除型小鼠有效下颌骨长度为(9.436±0.314)mm,经独立样本t检验统计学分析组间差异得知,Myo1h基因第30号外显子的敲除并未引起有效下颌骨长度差异性改变(图6 A、B);但测量4周龄小鼠的有效身长后发现野生型小鼠身长为(8.033±0.146)cm,纯合敲除型小鼠身长为(7.610±0.139)cm,经独立样本t检验统计学分析组间差异得知,Myo1h基因的突变能够导致纯合Myo1h小鼠身长的下降(图6 C,D)。
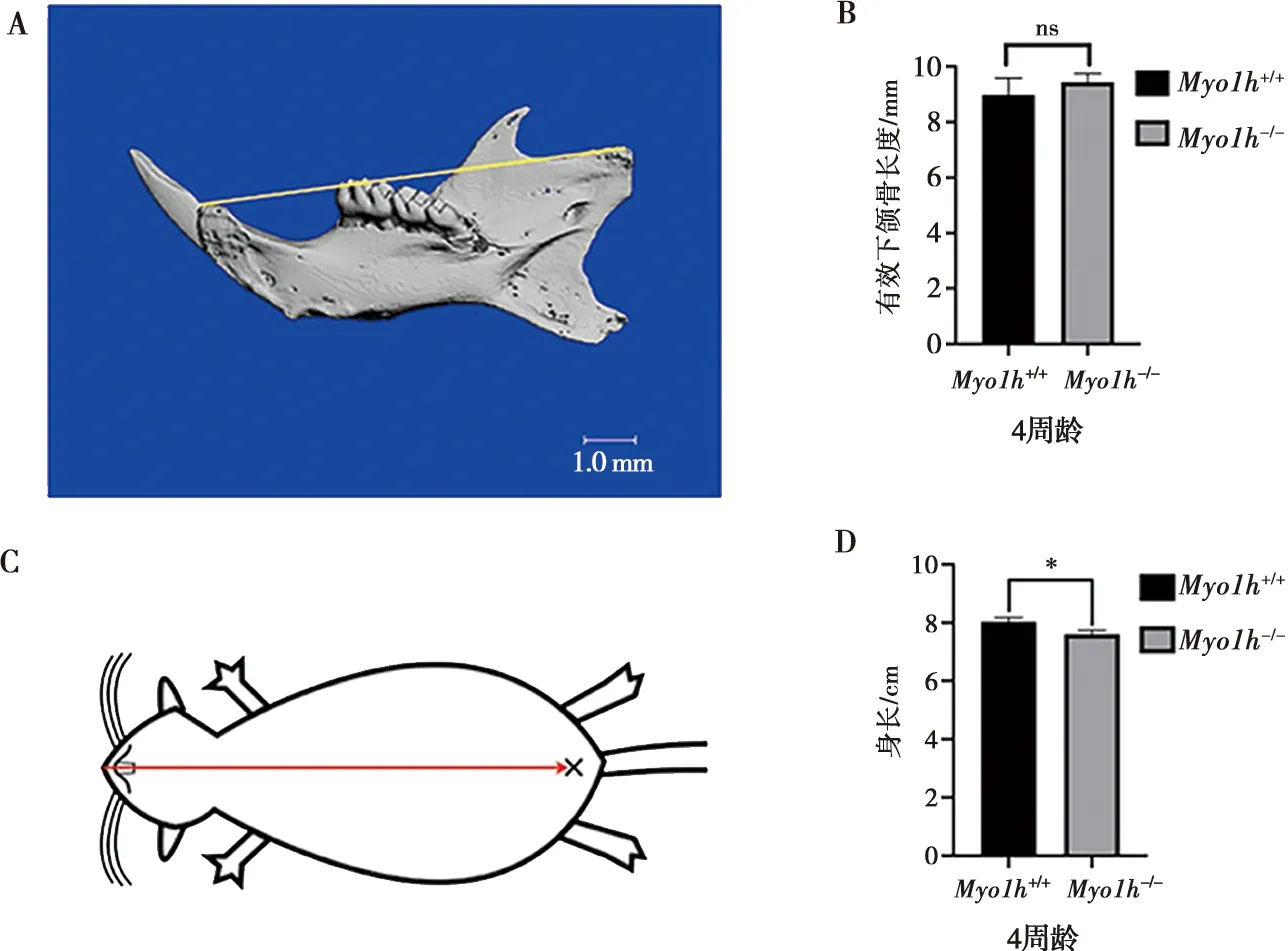
A:Micro-CT三维重建后有效下颌骨长度测量;B:有效下颌骨长度分析;C:有效身长测量;D:有效身长统计学分析结果;*:P<0.05,ns:无显著差异
3 讨 论
3.1 Myo1h基因与颌骨发育
肌球蛋白(Myosin)是骨骼肌中重要的结构功能蛋白,由头部、颈部以及尾部三部分共同构成,作为细胞骨架的分子马达,通过水解ATP酶产生能量为肌肉收缩提供动力[11]。根据其来源不同,可分为常规型肌球蛋白与非常规型肌球蛋白两大类[12],由于非常规型肌球蛋白在肌肉细胞中含量极低,故又称非肌肉蛋白[11],并且Myo1h基因在接受过正颌外科手术患者的咬肌中表达极低也被证实[13]。非常规型肌球蛋白在生物体中扮演着重要的角色,尤其在细胞运动、细胞器运输和信号转导过程中发挥着至关重要的作用[14-15]。非常规型肌球蛋白Ⅰ(Myosin 1H蛋白)只有一条重链,尾部较短,但它尾部结构域的多样性却提升了人们对该类肌球蛋白的认知[16]。而Myosin 1H蛋白属于Ⅰ类非常规型肌球蛋白同工酶之一[17],其在视觉、听觉以及平衡功能中有重要作用[15,18]。
研究表明,Ⅰ类肌球蛋白基因(Myo1h)的突变能够在生物体内引起不同的生长发育结果,比如Myo1h基因突变能够导致下颌后缩[7,19],其12q24位点的突变却能够造成下颌前突[3]。da Fontoura等[20]通过对269例受试对象的基因型-表型关联主成分分析研究发现PC2(principle component 2)与Myo1h上游SNP rs11066446相关联,揭示了在骨性错牙合畸形患者中,遗传因素在颌骨矢状位和垂直位发育中发挥重要作用。此外,Cruz等[4]在对54例骨性Ⅲ类患者和120例对照组的10个候选位点进行研究后发现,Myo1h(rs10850110 A Crispr/Cas9技术是通过sgRNA的序列保证基因编辑的特异性,该技术可应用于基因敲除、插入、碱基编辑等操作,免疫原性较低[23],与其他基因编辑方法相比较,Crispr/Cas9技术具有效率高、安全性可靠、适用范围广等优势,是构建基因敲除动物模型的有效工具[24]。基于Crispr/Cas9技术的诸多优点以及Myo1h基因的研究现状,本课题组采用Crispr/Cas9技术构建Myo1h基因敲除F0代嵌合体小鼠,利用F0与纯合野生型小鼠配笼繁殖得到F1代Myo1h杂合小鼠,再利用F1代杂合小鼠进行自交得到F2代Myo1h纯合子小鼠,并按照SPF级标准进行饲养繁殖,随后采用较为简便易操作的琼脂糖凝胶电泳的方法进行基因型鉴定,并对基因敲除后小鼠的下颌骨密度、有效下颌骨长度及有效身长等表型进行初步评估。 经过本研究体内、体外的验证,发现:①构建小鼠Myo1h同源基因第30号外显子全敲除模式动物后,能够获得基因型为Myo1h-/-的纯合小鼠,并能正常存活、繁殖且无明显致死性;②经提取肺组织、脑组织经荧光定量PCR实验,验证了纯合敲除型小鼠组织中Myo1h表达降低;③经髁突组织、脑组织以及肺组织免疫荧光染色表明纯合敲除型小鼠各组织中Myo1h表达降低;且在蛋白水平上也验证了纯合敲除型小鼠脑组织的Myo1h蛋白降低,实现了体内验证。以上三点证明本研究成功构建了Myo1h基因敲除小鼠。 本研究从颌骨长度、身长、下颌骨生长中心髁突中基因表达等方面,观察了Myo1h基因敲除后小鼠的生长发育特点。具体有以下几点:①经髁突组织、脑组织以及肺组织免疫荧光染色表明纯合敲除型小鼠各组织中Myo1h表达降低,本实验也首次在哺乳动物小鼠髁突软中内观察到Myo1h基因主要表达在软骨表层及肥大细胞层,推测该基因可能与髁突软骨发育相关;②Myo1h基因第30号外显子敲除后,F2代野生型小鼠与纯合敲除型小鼠下颌骨密度及有效下颌骨长度并未出现差异性改变,推测Myo1h基因对下颌骨的生长发育可能存在协同基因,具体的作用途径与机制有待进一步的研究;③Myo1h基因第30号外显子敲除后,纯合敲除小鼠的有效身长出现减少。 综上所述,本课题组获得能够正常存活、繁殖且稳定遗传的Myo1h基因敲除的模式小鼠。基于观察时间的限制,尽管暂未发现Myo1h基因敲除小鼠与野生型小鼠在下颌骨密度与有效颌骨长度方面有明显差别,但本研究发现构建的Myo1h突变小鼠的有效身长出现减少,提示Myo1h基因可能在躯干骨的生长发育中发挥着重要作用,同时观察到Myo1h基因主要在髁突软骨表层及肥大细胞层表达,推测该基因可能与髁突软骨发育相关,具体的作用机制还有待进一步研究,本研究填补了Myo1h基因位于尾域结构的第30号外显子在颌骨发育中的研究空白,并期进一步完善Myo1h基因在哺乳动物颅颌面骨骼生长发育中的作用,为后续对Myo1h基因的相关研究提供了稳定且可靠的小鼠动物模型。3.2 Myo1h基因敲除小鼠
3.3 Myo1h基因敲除小鼠的生长发育特点
猜你喜欢
分子诊断与治疗杂志(2022年9期)2022-10-09
中国美容医学(2019年1期)2019-03-01
中国美容医学(2018年3期)2018-05-23
现代仪器与医疗(2017年5期)2018-01-09
今日健康(2016年5期)2017-01-23
中国实用医药(2016年27期)2016-11-30
湖北农业科学(2014年11期)2014-09-10
华西口腔医学杂志(2013年3期)2013-12-23
