
ITS-nrDNA 片段分析确认安妮块菌Tuber anniae 是泛北极分布物种
2022-08-31 07:07刘虹李素玲李婷郭尚
山西农业大学学报(自然科学版) 2022年4期
刘虹,李素玲,李婷,郭尚
(1.山西农业大学 山西功能食品研究院 山西 太原 030031;2.首都师范大学 生命科学学院,北京 100048)
2015 年8 月至2017 年9 月,在中国北部河北省雾灵山地区和山西省吕梁山脉的中北部地区调查当地温带森林区地下真菌物种多样性时,采集到部分块菌菌根样本和一种白色的块菌子囊果,经鉴定为原产于美国的安妮块菌Tuber anniaeW.Colgan&Trappe[1-2]。块 菌 属 物 种分布 表 现 为 高度的洲际特有化,目前基于现代分子系统学基础上的块菌物种在自然界还没有同时分布在两个大洲的记录,之前许多欧洲种或北美种在亚洲或其它大洲的分布报告均被近期研究证实或源于标本的错误鉴定[3],或由于人类在洲际间引种树木等行为随共生树木被动带入,或由人工种植主动引入[4]。安妮块菌是第1 个被现代分子系统学证实的间断分布于2 个大洲以上、均可与当地本土树种共生的块菌属物种。深入分析安妮块菌,对进一步研究块菌属Tuber的起源演化、生物地理、种群传播乃至引种驯化等方面具有重要参考意义。
本研究以中国样本为基础,结合NCBI 数据库和相关研究采用的分子数据,对安妮块菌在中国的形态学特征及其变化特点、全球地理分布、共生树种等方面进行分析,为菌根食用菌开发、名贵商业块菌品种的引进以及森林生态经济模型的建立等方面提供新的思路和理论依据。
1 材料和方法
1.1 试验材料
1.1.1 菌根采集
本研究所用样本于2015 年8 月从离目标树木华北落叶松Larix principis-rupprechtiiMayr 和油松Pinus tabulaeformisCarr. 周边1~2 m 处,挖取15 cm×15 cm×10 cm 左右大小的土块,从中捡出小的细根,挑选表面有菌根样的根尖部位,用自来水将表面泥土冲洗去除,分类包好。将上述样本放置在4 ℃下冰箱内保存备用,存储时间不得超过7 d。
1.1.2 子囊果采集
本研究依据的子囊果于2016 年10 月采自中国河北省东北部雾灵山和2017 年9 月采自山西省吕梁山脉中北部的华北落叶松Larix principis-rupprechtiiMayr 树下土壤中。新鲜的子囊果标本经45 ℃烘干后保存于首都师范大学标本馆(BJTC)。
1.2 试验方法
1.2.1 形态学鉴定
对子囊果的宏观特征在体视显微镜下进行观察和描述,主要包括子囊果的形状、大小、颜色、基部是否具凹陷、子囊果表面有无毛、光滑、具疣或具瘤、产孢组织颜色、菌脉的类型等。子囊果的显微结构特征观察在光学显微镜下进行。首先采用手术刀进行徒手切片,以蒸馏水、3%KOH 作为浮载剂制作水封片。观察、测量与记录的项目主要包括子囊果包被的厚度、组织类型,包被表面毛状菌丝的形状、长度和宽度,子囊的形状、大小、子囊孢子的数目,子囊孢子的形状、大小、颜色、表面纹饰的类型、纹饰的高度等。
1.2.2 DNA 序列测定和比对
DNA 提 取、PCR 以 及 序 列 测 定 参 照Fan 和Huang[3,5]方 法,DNA 提 取 采 用 改 良 过 的CTAB 方法提取担子果总DNA 后,扩增核糖体DNA 内转录间隔区(internal transcribed spacer,ITS)片段,PCR 扩 增 引 物 为ITS1-F/ITS4(Gardes & Brun 1993)[6],ITS 的 扩 增 条 件 为:94 ℃预 变性3 min,94 ℃变性30 s,55 ℃退火45 s,72 ℃延伸1 min,35个循环,最后72 ℃延伸10 min;将PCR 产物送至北京中科西林生物技术有限公司(中国北京)进行纯化、测序和编辑,将序列提交到NCBI 数据库(http://www.ncbi(nlm.nih.gov/.)检索获取相似性较高的同源序列,与本研究自测序列相似度高于95%的序列汇总(表1)。
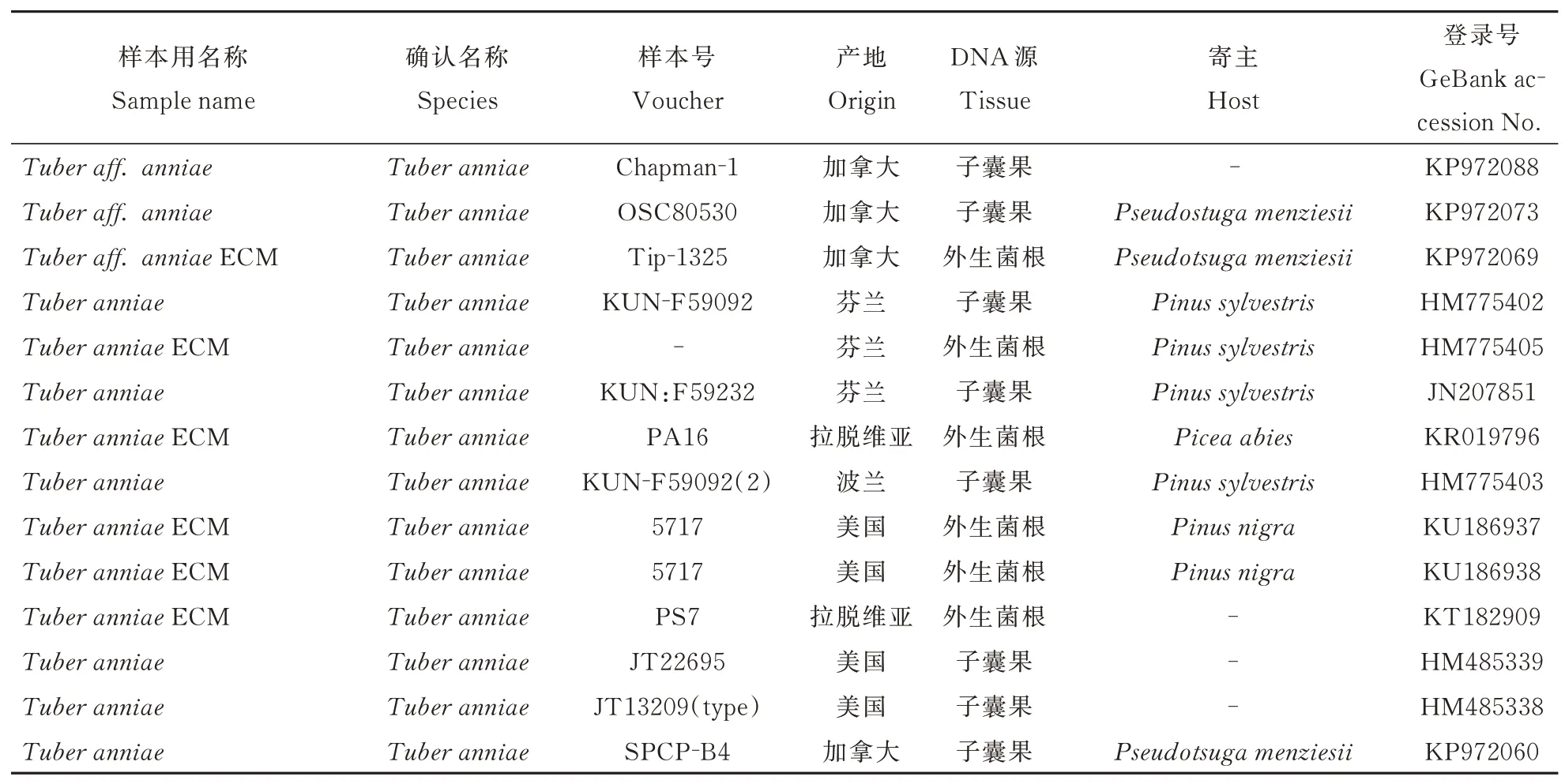
表1 本研究用样品名称、来源ITS 序列登录号Table 1 Scientific name,locality and ITS sequence information of the samples in this study
系统发育树的建立和分析参见Fan 和Huang[3,5]方 法。采 用ITS 序 列 数据集将供试标本序列于GenBank 中进行Blast 比对,用EditSeq 软件将供试标本序列和从GenBank 下载的序列进行比对,用Gblocks(http://molevol. cmima. csic. es/castresana/Gblocks_server. html)在线剪切软件进行剪切以及用Se-Al v.2.0[7]手工调整。以最大简约 法(Maximum parsimony, MP)用 PAUP*4.0b10.[8]构 建MP 树,采 用Clustal X 软件对各基因的序列进行比对;自举检验1000 次,分析获得的支 持 度。用TreeView32[9]查看树的 拓 扑 结 构,支持值大于70(MPBS≥70%)视为可靠的支持值。
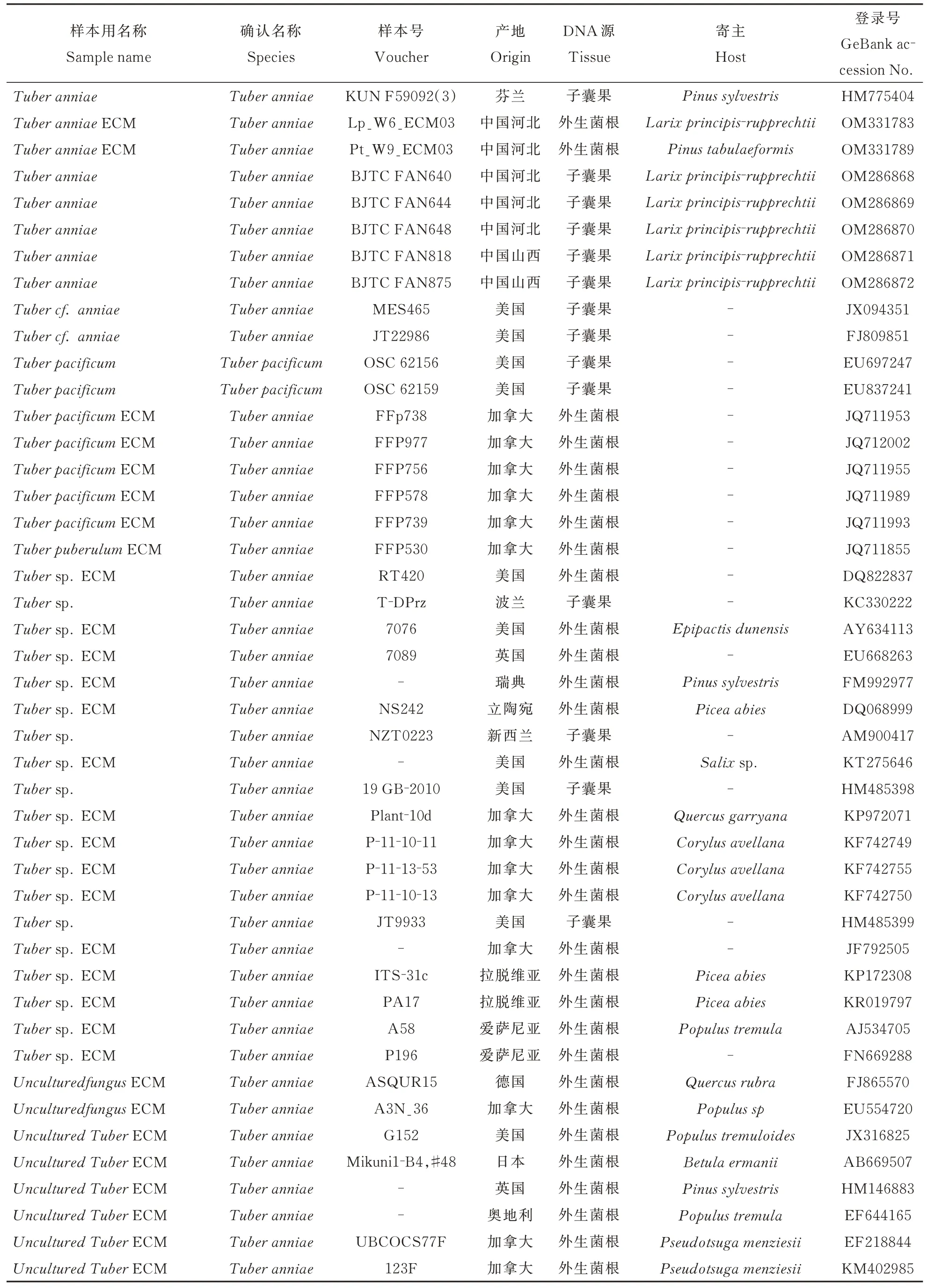
续表

续表
2 结果与分析
2.1 系统发育分析
本研究共采用61 条nrDNA-ITS 序列,其中7条为中国样本,其余54 条下载自NCBI 数据库。2条太平洋块菌Tuber pacificumTrappe,Castellano& Bushnell 和1 条来自美国的Tubersp. 的序列被选作分析用外类群。基于ITS 序列的ML 系统发育树(图1)显示所有内类群聚为在一支并获得100%的支持,这一大支进一步可以识别出3 个亚支,分别标注为A 亚支、B 亚支和C 亚支。其中B亚支自举值较高(BP=95%),A 亚支和C 亚支没有得到支持(图1)。A 亚支由34 条来自亚洲(包括2 条来自中国山西的序列)、欧洲和北美洲的序列组成;B 亚支的13 条序列来自亚洲(包括5 条来自中国河北和1 条日本序列)、欧洲和北美洲;C 亚支有10 条欧洲和北美洲的序列,包括来自安妮块菌Tuber anniae模式标本的序列(HM485338),另外有1 条序列(EF218844)在A 亚支附近,但没有支持值。BLAST 比对分析显示A 亚支序列间的ITS 相似度为98.9%~100%,B 亚支内序列间相似度为98.9%~100%,C 亚支内序列间相似度为99.5%~100%。相对而言,A 亚支和B 亚支之间ITS 序列相似度为98.4%~99.3%,B 亚支和C 亚支之间为97.2%~97.9%,A 亚支和C 亚支之间为97.0%~98.2%。
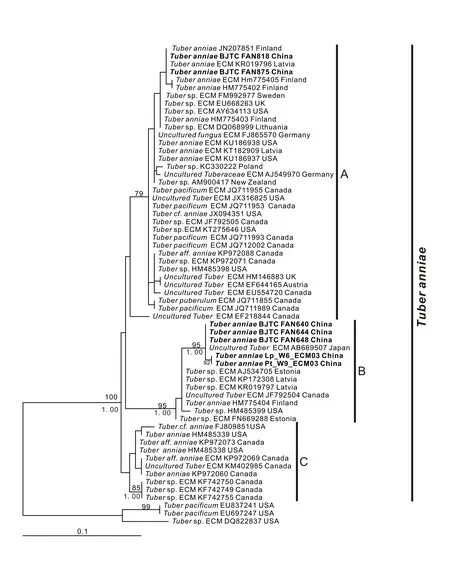
图1 基于nrDNA-ITS 基因片段的安妮块菌系统发育树(ML)Fig.1 Phylogenetic tree(ML)for Tuber anniae based on nrDNA-ITS sequences
2.2 分类学结果
基于中国标本的形态特征描述:子囊果近球形或不规则块状,直径0.8~2.2 cm,新鲜时白色或近白色,老熟后土褐色,表面平坦,光滑,偶尔部分区域有浅沟。孢体幼时白色,以后逐渐褐色,完全成熟时为暗灰褐色,切面观有大理石样白色脉纹,老熟后脉纹不再明显。气味清淡,味道没有记录。包被组织厚150~200 μm,分为2 层;外层为拟薄壁组织,由近球形或不规则的菌丝膨大细胞组成,细胞直径5~25 μm,薄壁或微微增厚到1~1.5 μm;内层为菌丝交织组织,菌丝无色,薄壁,直径2.5~5 μm,分隔,分支。子囊球形、近球形、卵圆形或梨形,60~85 μm×55~70 μm,无色,薄壁,内生1~4 个子囊孢子,多数无柄,偶尔具一短柄。子囊孢子广椭圆至近球形,成熟后褐色至暗褐色,表面具有网格状纹饰,25~45 μm×20~40 μm(不包括纹饰),通常孢子表面横向有3~5 个网格。
标本研究:中国:河北省兴隆县雾灵山,北纬40°35′,东经117°28′,海拔1801 m,华北落叶松Larix principis-rupprechtiiMayr 树下土壤中,2016年10 月10 日,李 婷LT001(BJTC FAN640);中国:河北省兴隆县雾灵山,闫向远YXY024(BJTC FAN644);同上YXY023(BJTC FAN648)。山西省关帝山,北纬37°50′,华北落叶松树下土壤中,2017 年9 月7 日,李婷LT001(BJTC FAN818);山西省关帝山,闫向远YXY024(BJTC FAN875)。
3 讨论
3.1 形态学讨论
本研究发现的中国标本与美国的安妮块菌模式标本之间的形态学差异主要表现在两个方面,一是子囊孢子形状,二是子囊果颜色。据原始作者发表该物种时的原始描述,其子囊孢子为球形、近球形或广椭圆形[1],而中国的标本为广椭圆形至近球形。根据我们多年来对块菌属物种标本的形态学鉴定观察,块菌属物种的孢子形状的稳定性依不同物种而不同,有些物种其孢子形状非常稳定,如金沙江块菌Tuber jinshajiangL. Fan 和石氏块菌T. shiiL. Fan&Y.W. Wang 这对姊妹种,它们之间形态学方面的唯一区别可能就是它们的子囊孢子形状,即金沙江块菌孢子是正球形,而石氏块菌为球形至近球形,这样的例子尚有欧洲的姊妹种T. sphaerospermum和T. oligospermum之间的形态学差别[10]。而在另一些物种中,子囊孢子的形状会依不同的个体或不同产地而有所不同,如中国西南地区的常见块菌阔孢块菌Tuber latisporumJ. Chen&P. G. Liu,其子囊孢子形状在一些子囊果中是典型的广椭圆形,而在另一些个体中其形状可能会是典型的近球形或以近球形为主,但DNA 分析结果为同一物种。对安妮块菌而言,本研究认为其孢子变化规律与中国的阔孢块菌类似。关于子囊果的颜色,中国的标本为白色或局部有些土褐色,欧洲标本与中国的相似,然而,模式标本的原始描述为暗绿褐色[1],这与中国和欧洲的标本明显不同。这一点让我们想起在几个中国特有块菌物种中观察到的一些特别的颜色现象,如曹氏块菌Tuber caoiL. Fan,正常的颜色为浅浅的灰绿色或灰褐色带有较明显的橄榄绿色调,但是我们从中也发现过暗红褐色的个体;同样,在黄绿色的光巨孢块菌Tuber glabrumL. Fan& S. Feng 子囊果中,经常可以发现近乎黑色的个体;同样的情况也出现在阔孢块菌中T. latisporum,据此推测美国的那份模式标本极有可能成熟过度或者颜色异常,否则无法解释ITS 片段的在种一级水平如此高度相似。另外,根据对中国块菌属Tuber物种的观察,安妮块菌所属的块菌属短毛块菌谱系(Puberulumlineage)的物种大多数种类其孢体颜色较浅,山西发现的安妮块菌孢体颜色为褐色,而河北采到的安妮块菌孢体颜色成熟时是深褐色(咖啡色),类似于欧洲的波氏块菌Tuber borchii。芬兰的标本形态描述孢体为浅褐色,但根据其子囊及子囊孢子照片[11],可以判断这些描述明显来自未成熟的子囊果,即安妮块菌Tuber anniae。

图2 安妮块菌成熟子囊果(左图:山西标本,右图:河北标本)Fig.2 Ascomata of Tuber anniae :Shanxi sample(Left),Hebei sample(Right)

图3 安妮块菌显微镜孢子广椭圆形至近球形Fig.3 Ascomata of Tuber anniae
3.2 安妮块菌Tuber anniae 的物种确定
安妮块菌最早发现于美国西北部太平洋沿岸的本土树种Pseudotsuga menziesii(Mirb.)Franco树下土壤中[1],这个物种发表后并没有引起人们太多的注意。2013 年,在北欧芬兰的Pinus sylvestris树下采到了一些白色的块菌[11],并将这些块菌与GenBank 数据库下载的DNA 序列进行了比对分析研究。共采用了24 条有关的ITS 片段来进行系统发育分析,结果显示这些密切相关的DNA 序列聚成了一个自举值很高的大支,这个大支内进一步可以分成3 个有较高自举值的亚支,这3 个亚支被作者定义为安妮块菌复合种Tuber anniaecomplex[11]。这个观点被后来的一些学者所接受[12-13]。本研究中,共收集了包括7 条中国自测序列在内的来自全球范围内的58 条相关序列,基于nrDNAITS 片段的ML 系统发育树显示所有内类群聚为在一支并获得100%的支持。这些序列也可以进一步识别为A、B、C 等3 个亚支,这3 个亚支基本上对应于[11]的cladesI、II、III。BLAST 分析显示,本研究的这3 个亚支内的ITS 相似度分别为98.9%~100%、98.9%~100%和99.5%~100%,而3 个亚支之间的ITS 相似度分别为98.4%~99.3%、97.2%~97.9% 和97.0%~98.2%,其中,A 亚支中来自中国山西的2 条序列和C 亚支中的安妮块菌模式种序列相似度高达99%,这些数值均远高于美国学者[14]和日本学者[15]基于大量数据分析提出的96%或95%的块菌属物种界定的ITS 相似度值。另一方面,本研究分析的ITS 序列系统发育树的3 个亚支中,A、B 两个亚支的样本都来自北美洲、欧洲和亚洲,C 亚支由来自欧洲和北美洲的样本组成,这说明它们彼此之间没有特定的地理对应现象,因而从生物地理角度也不能支持这3 个亚支为不同的块菌物种。
3.3 安妮块菌的共生树种及其在自然界的分布
近年的研究表明块菌属Tuber在种一级水平上表现出高度的洲际特有现象[16],过去几十年来所有在中国的欧美种类报道被证实均源于标本的错误鉴定[3]。据悉,到目前为止安妮块菌Tuber anniae是唯一有DNA 数据支持的欧、亚、北美3 个大陆共有的块菌属物种。在北美洲,安妮块菌最早在美国西北部太平洋沿岸地区的本土树种Pseudotsuga menziesii树下土壤中发现,最近采用DNA技术在美国东部马萨诸塞州Arnold 植物园引进的欧洲松Pinus nigra根部探测到了它的存在[11],此外,在Alaska 和Nebraska 也 发现了它的 分 布[13]。在加拿大东部和西部,尤其在British Columbia,这个物种被证实为当地常见的块菌属物种[13]。在欧洲的波罗的海周边地区,从树木根尖提取的DNA片段证实了安妮块菌在芬兰、爱沙尼亚[10-11]和拉脱维亚(本文)的存在,但仅在芬兰从Pinus sylvestris树下土壤中采到了子囊果[11]。在亚洲的日本,一条来自北海道亚洲本土树种Betula ermanii根尖提取的ITS 序列(AB669507)比对到了这个物种。在中国,安妮块菌在两个中国本土的森林建群树种华北落叶松Larix principis-rupprechtii和油松Pinus tabulaeformis的根尖上检测到了其存在,在华北落叶松下发现其子实体。作者初步调查了本物种发现地河北雾灵山和山西吕梁山产地周边及附近的生物环境,没有发现有大型植物园一类的可能导致外来植物人为引进的场所,也没有查阅到河北雾灵山和山西吕梁山有引进欧美树种的历史文件记录,也就是说没有发现这个和树木形成外生菌根的块菌物种意外或间接被引入该地区的可能因素存在,因此推断这个块菌物种应该是自然分布到本区域的。虽然近来安妮块菌在新西兰松林内被探测到,但它生长在从欧洲引进的外来树种形成的人工林内,据此推测这个物种在南半球的分布可能源于人类从北半球引种树木的结果[11-12]。进一步的地理分布分析揭示,这个物种明显倾向于高纬度地区。在北美洲,安妮块菌均见于美国和加拿大的北部地区;在欧洲,北部的波罗的海周边可能是它在欧洲的分布中心;在亚洲的日本,只发现于北海道一地;而在中国,仅在北方的河北雾灵山和山西吕梁山脉中北部的高海拔地区发现其分布。就目前数据而言,本种分布最北部的记录是北纬61°35′,位于芬兰[11],中国吕梁山脉中部关帝山的发现地位于北纬37°50′,可能是本种已知分布的最南端。菌根研究证实,安妮块菌能够与多种树木共生形成菌根,其范围包括裸子植物(如Pinus sylvestris,Larix principis-rupprechtii等)和被子植物(如Pseudotsuga menziesii,Betula ermanii等),既可以是本土树种,也可以是外来树种[11-13]。安妮块菌能够在自然界多种不同的土壤[11-13]中产生子囊果证明其有很强的环境适应能力,这在一定程度上解释了它在自然界广泛的地理分布。
4 结论
本研究确认了分布于欧洲、北美洲、亚洲高纬度或高山地区的安妮块菌复合种Tuber anniae complex属于同一个物种。安妮块菌在自然界间断分布于北美洲、欧洲和亚洲,可与多种树木形成外生菌根,广泛分布于北半球高纬度或高山地区,是一个泛北极分布的块菌属物种。对于块菌属这一在洲际间物种高度特有化的地下菌类群而言,安妮块菌对解释块菌属物种在全球范围内的起源演化以及生物地理等方面研究有重要意义。此外,安妮块菌是一种有潜在经济价值的食用真菌,其子囊果成熟后香气浓郁,类似于欧洲的波氏块菌Tuber borchii(仅次于意大利白块菌T. magnatum的著名品种),商业上属于在国际市场上广受欢迎的“白松露”之一种。它在中国北方的发现,尤其是它和油松这一北方温带干旱地区重要造林树种能形成外生菌根的事实为我们下一步的植树造林、菌根食用菌开发、名贵商业块菌品种的引进以及森林生态经济模型的建立等方面提供了一个新的思路和方向。
猜你喜欢
辽宁林业科技(2022年2期)2022-11-24
西南农业学报(2022年8期)2022-10-28
安徽农学通报(2022年10期)2022-06-30
亚热带资源与环境学报(2022年2期)2022-06-30
热带亚热带植物学报(2022年3期)2022-06-10
中国食用菌(2022年5期)2022-06-08
知识窗(2021年2期)2021-08-14
农业科技与信息(2020年20期)2020-11-12
阅读(快乐英语高年级)(2019年4期)2019-09-10
阅读(快乐英语高年级)(2019年3期)2019-09-10
