
秋葵AeMYB1R1转录因子基因克隆及对胁迫的响应
2022-08-19 00:37朱志鹏刘慧玲吴可鑫虞健翔王博文
西北植物学报 2022年6期
朱志鹏,刘慧玲,吴可鑫,虞健翔,王博文,孙 淼,2*
(1 江苏省盐土生物资源研究重点实验室,盐城师范学院海洋与生物工程学院,江苏盐城 224002;2 南京农业大学,作物遗传与种质改良国家重点实验室,南京 210095)
秋葵[Abelmoschusesculentus(L.) Moench]为锦葵科(Malvaceae)秋葵属(AbelmoschusMedicus),又称黄秋葵,在世界范围内广泛种植,具有观赏价值、食用价值和药用价值[1]。秋葵的茎为圆柱形,疏生散刺;其叶呈掌状,裂片阔至狭,托叶线形,被疏硬毛;其蒴果为筒状尖塔形。秋葵性喜暖,耐热力强,多生长于热带和亚热带地区[2]。秋葵中含丰富的钾、钙、铁、锌、锰等元素,具有极高的营养价值,已逐渐成为一种新型蔬菜;它还具有果胶类物质、膳纤维、酚类化合物等多种生物活性物质,可以有效防治心血管疾病、消化疾病等[3]。
植物在自然生长发育过程中会面对各种生物或非生物胁迫影响,面对这些逆境条件,植物体内通常会通过一系列的生理、生化反应作为自身的防御保护措施[4]。在植物逆境响应系统中,转录因子对功能基因表达的调控是植物逆境响应的关键环节[1]。MYB基因家族是与生理代谢、细胞的形态和模式建成等生理过程有关的一类转录因子,广泛参与了植物器官的生长发育以及对非生物胁迫的特异性响应[5]。迄今为止,研究者已在拟南芥(Arabidopsisthaliana)[6]、棉花(GossypiumhirsutumL.)[7]、大豆(GlycinemaxL.)[8]、漆树(Toxicodendronvernicifluum)[9]等多种植物中扩增获得MYB类基因,且过度表达MYB基因的拟南芥[10]、番茄(SolanumlycopersicumL.)[11]等均表现出了较强的非生物胁迫抗性。但秋葵MYB基因的功能研究尚无相关报道。
本研究以‘北海道1号’秋葵为实验材料,扩增获得了1个MYB类转录因子基因AeMYB1R1,基于生物信息学技术分析其核苷酸序列和编码的氨基酸序列。并采用实时荧光定量PCR(qRT-PCR)技术分析了AeMYB1R1基因在秋葵不同器官(叶、茎、根)和非生物胁迫(低温、干旱、盐和高温胁迫)中的表达特异性,为进一步开展秋葵MYB类转录因子的分子调控机制研究提供理论依据和实验基础。
1 材料和方法
1.1 试验材料
本试验研究对象为秋葵(图1)品种‘北海道1号’(A.esculentus, cv ‘Hokkaido No.1’),待秋葵幼苗长至三叶一心时移入培养箱进行胁迫处理。胁迫处理试验分为4组,分别为38 ℃的高温胁迫处理(A组),200 mmol/L NaCl溶液的盐胁迫处理(B组),200 g/L PEG6000溶液的干旱胁迫处理(C组)和-7 ℃低温胁迫处理(D组),每组6株用于采样,每个处理设置3个生物学重复。在处理6、12和24 h时分别取秋葵相同位置的叶、茎、根等组织,液氮速冻后保存于-80 ℃冰箱以用于后续实验。
1.2 方 法
1.2.1 RNA的提取与cDNA的合成使用RNA提取试剂盒(RNAsimple Total RNA Kit试剂盒,购自北京天根生化科技有限公司,目录号:DP419)分别提取秋葵的不同时期和不同组织的Total RNA,提取完成后,参照反转录试剂盒(PrimeScriptTMRT Master Mix试剂盒,购自北京TaKaRa生物医药科技有限公司,目录号:RR036A)的使用说明,将提取的总RNA反转录为cDNA。
1.2.2 秋葵AeMYB1R1基因克隆根据本课题组上传的秋葵转录组数据库(NCBI登录号:SRR13983395)[12],检索获得编码秋葵MYB转录因子的基因AeMYB1R1 (GenBank登录号:OM963000),利用DNAMAN设计1对引物(正向引物:5′-CTGCGGAGGAGTGAAACTGT-3′,反向引物:5′-TCTCACCGCGATGATTAGCC-3′)。以反转录得
到的cDNA为模板,使用20 μL体系(PrimeSTAR®Max DNA Polymerase 10.0 μL,cDNA 1.0 μL,正、反向引物各1.0 μL,ddH2O 7.0 μL)进行AeMYB1R1基因PCR扩增(高保真PCR酶购自北京TaKaRa生物医药科技有限公司,目录号R045A)。PCR反应程序为:98 ℃预变性10 s,98 ℃变性10 s,58 ℃退火10 s,72 ℃延伸10 s,共35个循环,最后72 ℃延伸10 min。PCR扩增产物使用1.2 %琼脂糖凝胶进行电泳检测。
1.2.3AeMYB1R1基因的生物信息学分析采用ExPASy-Translatetool可对AeMYB1R1核酸序列进行翻译,将AeMYB1R1基因序列翻译成相应的氨基酸序列。使用ExPASy-Protparamtool分析蛋白质的理化性质。信号肽分析利用SignalP 5.0 Server软件完成,跨膜结构分析利用TMHMM Server v.2.0软件完成。通过NCBI网站进行BLAST对比,得到AeMYB1R1蛋白的保守域以及其他物种MYB蛋白的氨基酸序列。选用SOPMA软件进一步分析蛋白质二级结构,蛋白质三级结构模型由Swiss-Model在线建立。DNAMAN 6.0软件被用于物种间MYB蛋白的多重对比和AeMYB1R1蛋白的亲/疏水性分析。AeMYB1R1蛋白糖基化位点的预测采用NetOGlyc软件。使用Netphos3.1 Server PredictProtein对AeMYB1R1蛋白的磷酸化位点进行分析。使用Plant CARE识别转录因子识别并特异结合的目标基因旁侧的DNA序列(顺式作用元件)。本研究所用生物信息学分析在线网址及软件如表1所示。

表1 生物信息学分析在线网址及软件Table 1 Bioinformatics analysis online URL and software
1.2.4 实时定量PCR反应基于PCR扩增且经过测序分析的AeMYB1R1基因序列,设计qRT-PCR荧光定量引物,AeMYB1R1基因表达正向引物为5′-CTGCGGAGGAGTGAAACTGT-3′,反向引物为5′-TCTCACCGCGATGATTAGCC-3′。以秋葵AeActin7为内参基因,正向引物为5′-TCGCAGACCGTATGAGCAAG-3′,反向引物为5′-GGTGCTGAGTGATGCCAAGA-3′。使用qRT-PCR检测系统分析该基因在不同组织中和不同非生物胁迫下的表达情况。反应体系20 μL,包括TB Green Premix Ex Taq Ⅱ(Tli RNaseH Plus)(2×)10.0 μL,cDNA 2.0 μL,正、反向引物各0.8 μL,ROX Reference Dye Ⅱ(50×)0.4 μL,ddH2O 6.0 μL(TB Green®Premix Ex TaqTMⅡ试剂盒购自北京TaKaRa生物医药科技有限公司)。使用GraphPad Prism 8.0软件,采用2-ΔΔCt方法对‘北海道1号’秋葵AeMYB1R1基因在不同器官和不同胁迫条件下的相对表达量进行分析。
2 结果与分析
2.1 秋葵AeMYB1R1基因的克隆及序列分析
对AeMYB1R1基因的PCR扩增结果显示,得到一条长约 1 100 bp 长度的目的条带(图2),目的片段大小基本符合预期。本研究从‘北海道1号’秋葵中克隆得到AeMYB1R1基因,结果显示AeMYB1R1基因的开放阅读框(open reading frame, ORF)长度为 1 056 bp,编码352个氨基酸(图3)。
2.2 AeMYB1R1蛋白的氨基酸组成及理化性质
ExPASy-Protparamtool对AeMYB1R1蛋白序列和理化性质分析表明,其分子量为37 891.57 Da,理论等电点为8.75,分子式为C1647H2615N489O515S12,总原子数为5 278,脂肪指数为70.37,总平均亲水性为-0.495,不稳定指数(The instability index Ⅱ)为40.22。根据不稳定指数标准,当指数<40时为稳定蛋白,指数>40时为不稳定蛋白,因此由不稳定性指数可判定AeMYB1R1蛋白为不稳定蛋白质[13]。在碱基组成方面,AeMYB1R1基因序列由A、C、G、T四种碱基组成,其中含量占比最高的为腺嘌呤A,其数量为306,占比29.0%;胞嘧啶C的数量为254,占比24.1%;胸腺嘧啶T的数量为250,占比23.7%;鸟嘌呤G的数量为246,其占比为23.3%。表2显示,在氨基酸组成方面,组成AeMYB1R1蛋白的20种氨基酸中丝氨酸(Ser)含量最高,色氨酸(Trp)数量最少,包括其中带负电荷残基总数(Asp+Glu)32个以及带正电荷残基总数(Arg+Lys)36个。
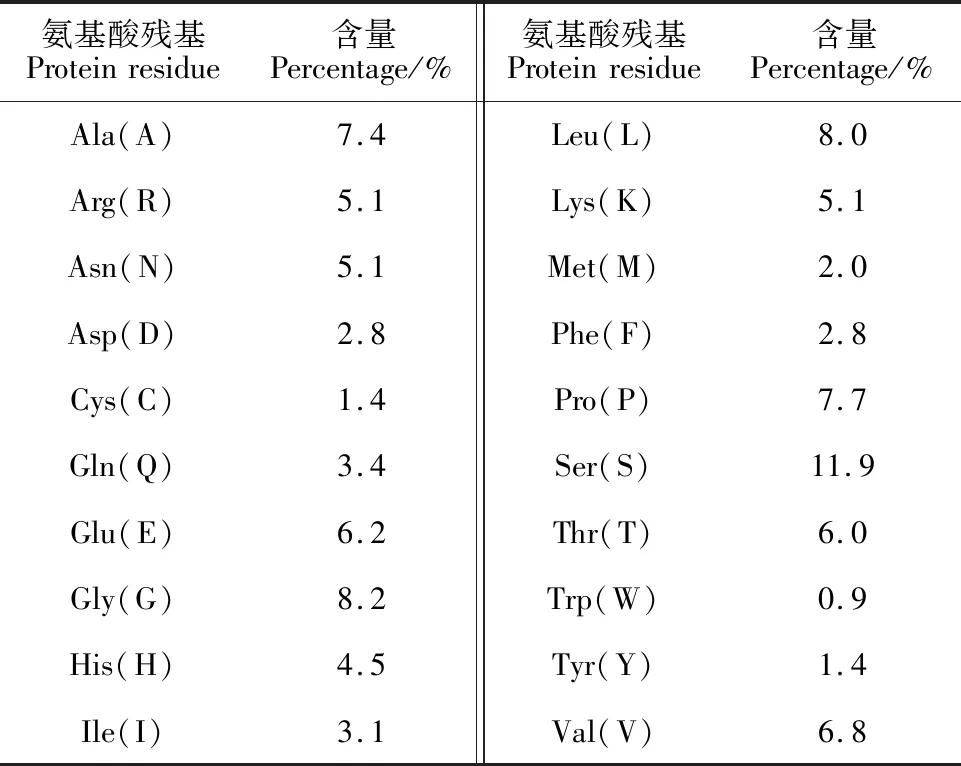
表2 AeMYB1R1蛋白中的20种氨基酸含量表Table 2 The list of 20 amino acids in AeMYB1R1 protein
2.3 AeMYB1R1蛋白的保守结构域分析
NCBI-CDD在线工具分析结果显示,AeMYB1R1的保守结构域为第104~156位氨基酸的序列区域,特定匹配(Specific hits)得分最高的是myb_SHAQKYF,该匹配所属SANT超家族(SANT superfamilies),而SHAQKYF类MYB家族转录因子(域架构ID 10019308)在植物的发育过程和防御反应中起着核心作用。
利用NCBI在线网站(https://www.ncbi.nlm.nih.gov/)检索木槿(Hibiscussyriacus)、陆地棉(Gossypiumhirsutum)、榴莲(Duriozibethinus)、可可(Theobromacacao)等19种植物的MYB1R1氨基酸序列,将其与秋葵AeMYB1R1氨基酸序列共同导入MEGA-X进行同源性比对。结果(图4)表明,AeMYB1R1的氨基酸序列与其他物种中MYB1R1的氨基酸相似度均超过65%,具有较高的一致性,说明MYB1R1在不同植物物种间的保守性较高。
2.4 AeMYB1R1蛋白结构和信号肽预测及其跨膜结构分析
利用SOPMA对AeMYB1R1蛋白进行二级结构分析。结果显示,AeMYB1R1编码蛋白由α-螺旋、延伸主链、β-转角、无规则卷曲4种结构组成,分别占比为21.31%、9.94%、2.27%和66.48%。其中,α-螺旋和无规则卷曲是AeMYB1R1主要的二级结构(图5,A)。Swiss-Model对蛋白质三维结构的预测分析结果与二级预测结果基本相符(图5,B)。以Entamoeba histolytica EhMybS3(SMTL ID:6nvz.1)为模板,所构建的模型与其序列一致性为41%,覆盖率为15%,氨基酸序列范围为104~158。
对AeMYB1R1转录因子的信号肽预测分析未发现特异性结果,推测AeMYB1R1蛋白序列中没有信号肽的存在,推断该蛋白无跨膜结构。TMHMM Server v.2.0分析结果显示,AeMYB1R1蛋白的1~352位氨基酸均位于细胞膜表面,无跨膜螺旋区。氨基酸序列不存在跨膜区和信号肽,推测该蛋白属于定位在细胞质基质或细胞器基质中的蛋白,不属于膜蛋白或分泌蛋白。
2.5 AeMYB1R1氨基酸序列的亲/疏水性预测
AeMYB1R1蛋白的亲/疏水性分析结果表明,在AeMYB1R1蛋白的亲水性区域(图6,A),第163位赖氨酸(Lys)的亲水性最强;在AeMYB1R1蛋白的疏水性区域(图6,B),第118位亮氨酸(Leu)的疏水性最强。组成AeMYB1R1蛋白的氨基酸中,35%为亲水性氨基酸,65%为疏水性氨基酸,AeMYB1R1蛋白的疏水区域多于亲水区域,因此推测其属于疏水性蛋白。
2.6 AeMYB1R1蛋白的修饰位点
2.6.1 AeMYB1R1蛋白的磷酸化位点预测NetPhos 3.1 Server预测结果表明(图7),AeMYB1R1蛋白中共发现86处可能存在的磷酸化位点以及47个特异性激酶位点,包含了31个丝氨酸(Ser)、14个苏氨酸(Thr)和2个酪氨酸(Tyr)。同时,AeMYB1R1蛋白涉及到13种特异性激酶,包括以钙和磷脂依赖的蛋白激酶PKC,以环腺苷酸依赖的蛋白激酶PKA,环鸟苷酸依赖的蛋白激酶PKG,以及作用于下游的磷酸化级联系统的p38MAPK、cdc2、cdk5,还包括DNA依赖性蛋白激酶DNAPK,属于丝氨酸/苏氨酸激酶的糖原合成酶激酶-3(GSK-3),参与信号传导的蛋白激酶核糖体S6激酶(RSK),酪蛋白激酶1、2(CKI、CKII),以及其他植物蛋白激酶INSR、ATM。
2.6.2 AeMYB1R1蛋白的糖基化位点预测采用在线网站YinOYang 1.2 Server(http://www.cbs.dtu.dk/services/YinOYang/),本研究对AeMYB1R1蛋白潜在的O-糖基化位点进行预测和分析。结果表明,氨基酸序列的第2、52、58、59、60位等共19处O-糖基化位点,其中位点所对应的氨基酸分别为6个苏氨酸(Thr)和13个丝氨酸(Ser)(图8,A)。NetNGlyc 1.0 Server (http://www.cbs.dtu.dk/services/NetNGlyc/)在线工具主要预测N-糖基化位点,分析结果表明,第50、158、208、340位含有潜在的N-糖基化位点,但此序列不包含信号肽,无信号肽的蛋白质不可能暴露在N-糖基化机制中,也不可能在体内被糖基化(图8,B)。
2.7 AeMYB1R1蛋白的同源性分析
利用MEGA-X对秋葵和其他20种植物的MYBs构建系统进化树,分析其同源进化关系(图9)。结果表明,AeMYB1R1与同属于锦葵科(Malvaceae)的木槿(Hibiscussyriacus)和陆地棉(Gossypiumhirsutum)等植物的MYB蛋白进化关系最近,与木棉科(Bombacaceae)的榴莲(Duriozibethinus)、梧桐科(Sterculiaceae)的可可(Theobromacacao)等植物有着近缘关系,而与茄科(Solanaceae)、芸香科(Rutaceae)、葡萄科(Vitaceae)、豆科(Leguminosae)、胡桃科(Juglandaceae)等植物MYB1R1蛋白的进化关系较远。因此,同属锦葵科的秋葵、木槿、陆地棉、可可等植物中的MYB1R1蛋白在进化中更为保守。
2.8 AeMYB1R1在秋葵不同组织中的表达
通过qRT-PCR检测分析‘北海道1号’秋葵AeMYB1R1基因在不同组织中的相对表达。结果(图10)显示,AeMYB1R1基因在‘北海道1号’秋葵的叶片、茎部和根部都有不同程度的表达,在叶部表达量最高,其次是根部,而在茎部的表达水平最低。其中,秋葵叶部AeMYB1R1基因的表达水平是茎的21.23倍,是根部的7.95倍。
2.9 在非生物胁迫下AeMYB1R1的表达特性
通过顺式作用元件分析,在AeMYB1R1序列中预测到了CGTCA-motif(茉莉酸甲酯响应元件)、ABRE(脱落酸响应元件)、MBS(干旱响应元件)、G-box(光响应元件)等多个与植物激素和逆境胁迫相关的响应元件,表明该基因可能在秋葵响应非生物胁迫的转录调控机制发挥作用。
进一步通过qRT-PCR分析发现,‘北海道1号’秋葵AeMYB1R1基因在高温胁迫、盐胁迫、干旱胁迫和低温胁迫下具有表达特异性(图11),在秋葵不同组织和不同处理时间相对表达水平存在明显差异。
高温(38 ℃)胁迫数据表明(图11,A),高温处理6 h时,AeMYB1R1基因在秋葵叶片相对表达水平最高,分别为茎的7.64倍、根的1.45倍;处理12 h,AeMYB1R1基因在叶片中的表达量有略微增加,
而在茎部和根部的表达量显著提高,与6 h数据相比,叶、茎、根中AeMYB1R1的相对表达量分别提高了15.48%、83.34%和110.34%;处理24 h,AeMYB1R1基因相对表达量在叶、茎、根中均有显著提高,相较于6 h,叶、茎、根中AeMYB1R1的相对表达量则分别提高了2.69、3.50和3.28倍。
盐(200 mmol/L NaCl溶液)胁迫试验数据表明(图11,B),盐处理6 h,AeMYB1R1基因在叶中的表达量分别为茎的5.03倍、根的29.80倍;处理12 h时,AeMYB1R1基因的相对表达水平呈现显著上升趋势,与6 h数据相比,叶、茎、根中AeMYB1R1的相对表达量分别提高了1.46、8.45和26.33倍;处理24 h,AeMYB1R1基因在叶和根部的相对表达量明显提升,并在根部显著表达,与6 h数据相比,叶、茎、根中AeMYB1R1的相对表达量分别提高了14.85、10.38和1 339.17倍。
干旱(20% PEG6000溶液)胁迫试验数据表明(图11,C),处理6 h时,AeMYB1R1基因在秋葵的叶、茎和根部的相对表达水平较低,且无显著差异;干旱胁迫12 h时,AeMYB1R1的相对表达水平开始上升,与6 h数据相比,叶、茎、根中AeMYB1R1的相对表达量分别提高了99.20%、400.14%和160.12%;干旱胁迫24 h,AeMYB1R1基因在秋葵的叶片和根部的表达量提升较快,且在根部相对表达量最高,与6 h数据相比,叶、茎、根中AeMYB1R1的相对表达量分别提高了9.38、12.08和125.96倍。
低温(-7 ℃)胁迫试验数据表明(图11,D),低温处理6和12 h时,AeMYB1R1基因在秋葵的叶、茎和根部的相对表达水平缓慢提高,未出现显著变化;24 h时,AeMYB1R1基因在秋葵的叶、茎和根部的相对表达量提高较为显著,相比于12 h时,AeMYB1R1基因在秋葵的叶、茎和根部分别提高了5.43、7.21和2.77倍。
3 讨 论
非生物胁迫可通过破坏系统结构和功能影响渗透调控和离子平衡等,直接或间接地损害植物细胞,从而严重影响植物的生长发育[14]。据报道,植物体内主要存在5类与抗逆性相关的转录因子,它们精准地调控下游功能基因适时适量表达,实现对非生物胁迫的应答,包括NAC、MYB、WRKY、bZIP和ERF/DREB[15]。其中,MYBs成员众多,分布广泛,参与植物次生代谢、激素转导和环境因子反应,并在细胞分化、细胞周期和叶片形态发生中起着重要的调节作用[16]。本研究从‘北海道1号’秋葵中扩增获得了1个AeMYB1R1基因,其氨基酸序列具有较高的保守性。同时,AeMYB1R1属于SHAQKYF类MYB家族转录因子,研究表明,SHAQKYF类MYBs可以调控植物的防御反应机制[17],这也暗示了AeMYB1R1可能参与秋葵的抗胁迫机制。
在柳枝稷(PanicumvirgatumL.)中,PvMYB4在叶片、叶鞘、节间、花序和节间表达量较高,但在根部表达受限[18]。在茶树(Camelliasinensis)中,CsMYB47和CsMYB17在叶和芽中高表达,CsMYB34则在老叶、茎和根部特异性表达[19]。Yang等[20]证明,在白果树(GinkgobilobaL.)中,MYB41在未成熟和成熟果实中表现出最高的转录积累,MYB1在根和茎部高表达,而MYB5、MYB7、MYB21、MYB24、MYB25、MYB49、MYB52和MYB57则在所有组织中几乎无表达。以上研究表明,MYBs具有显著的组织表达特异性。在橡胶(Heveabrasiliensis)的愈伤组织萌发中,HbMYB1R1被鉴定为体细胞胚胎发育的关键标记基因[21]。本研究中,AeMYB1R1在秋葵叶中表达量最高,其次是根部,而在茎部的表达水平最低,说明AeMYB1R1基因在秋葵中具有器官表达特异性,并可能在秋葵的发育过程中起到重要作用。
除了在植物生长发育中的功能,MYBs在植物响应胁迫的作用逐渐被揭示。苹果中的MdMYB308L通过与MdbHLH33的相互作用,增强其与MdCBF2和MdDFR启动子的结合,从而充当苹果耐寒性和花青素积累的正调节剂[22]。黄角牛(Xanthocerassorbifolium)中的XsMYB44通过触发气孔关闭以维持水位和调节ROS稳态,进而抵御干旱和高温的联合胁迫[23]。在本研究中,我们发现AeMYB1R1特异性地应答高温、盐、干旱和低温胁迫;同时,与高温和低温胁迫相比,AeMYB1R1在盐胁迫和干旱胁迫下的相对表达量更为显著。据报道,玉米ZmMYB3R受干旱、盐分和脱落酸诱导,其在拟南芥中的过表达增强了转基因植物对干旱胁迫和盐胁迫的耐受能力[24]。番茄LeMYB49可作为一种正调节剂,通过增强清除活性氧的能力,抑制细胞膜损伤和细胞死亡,保护叶绿体,从而提高对盐胁迫和干旱胁迫的耐受性[25]。芝麻中的SiMYB75受干旱胁迫、盐胁迫、脱落酸和甘露醇强烈诱导,在根部特异性表达,并通过脱落酸介导的途径正向调节干旱、盐分和渗透胁迫反应[26]。以上研究表明,部分MYBs参与了脱落酸介导的活性氧清除途径,从而响应盐碱和干旱环境带来的渗透压胁迫。因此,我们推测AeMYB1R1可能受脱落酸介导,通过调节活性氧清除系统,从而参与秋葵响应盐和干旱胁迫的分子机制。
综上所述,本研究以‘北海道1号’秋葵为试验材料,扩增并获得了1个编码AeMYB1R1转录因子的基因AeMYB1R1,并发现其在叶中特异性表达。通过对该基因在多种非生物胁迫中表达模式的分析,发现AeMYB1R1特异性地响应高温胁迫和低温胁迫,尤其在盐胁迫和干旱胁迫下,该基因的相对表达量显著提高,因此推测AeMYB1R1是调控秋葵抗盐和抗旱的关键转录因子。本研究为了解AeMYB1R1基因在秋葵生长发育和抗逆境机制提供了一定的理论参考,以便为秋葵的育种和抗逆性增强工作奠定基础。
猜你喜欢
中国农学通报(2022年12期)2022-06-01
中国糖料(2022年2期)2022-04-06
中国种业(2021年11期)2021-11-25
西藏文学(2020年5期)2020-09-21
快乐语文(2019年35期)2020-01-06
中国诗歌(2019年6期)2019-11-15
动漫界·幼教365(小班)(2019年10期)2019-10-28
中学生物学(2019年7期)2019-10-17
老友(2019年8期)2019-09-09
决策探索(2017年11期)2017-06-23
