
火龙果的抗旱生长及生理响应
2022-08-06 09:05李莉婕赵泽英黎瑞君冯恩英岳延滨聂克艳
西南农业学报 2022年6期
李莉婕,赵泽英,黎瑞君,冯恩英,岳延滨,袁 玲,聂克艳
(1.贵州省农业科技信息研究所,贵阳 550006;2.西南大学资源环境学院,重庆 400716)
【研究意义】植物应对干旱胁迫的响应是生态学研究的重要领域[1]。火龙果(Hylocereuspolyrhizus)于20世纪90年代初引进我国种植,因其生长快,经济效益高,在川、黔、滇、桂等地的北盘江流域、红水河流域、金沙江干热河谷地区的种植规模迅速扩展[2-3]。这些区域季节性干旱严重,水分是作物生长的主要限制因素。因此,研究火龙果对干旱胁迫的响应,对于指导其合理灌溉具有重要意义。【前人研究进展】火龙果叶片退化为刺,茎蔓呈肥厚肉质状,含有能够贮存可溶性物质和水的大液泡,以及大量能够提高细胞渗透压和保水性的晶细胞和粘液物质[4],可在土壤缺水的情况下长时段供给植株光合作用所需水分[5]。作为景天酸代谢途径植物,良好的灌溉能够提高火龙果夜晚CO2吸收速率[6],在商业化生产中需要充足的水分供应才能满足其生长发育需求。火龙果每3 d需要摄入0.5 kg水分才能满足火龙果生长的需求[7],70%~80%的土壤相对含水量适宜火龙果生长发育和产量形成[8]。火龙果适宜生长在土壤较为潮湿的环境,对淹水胁迫有一定的适应能力[9],但淹水胁迫会导致根系缺氧烂根,枝条长斑,严重影响其生长发育及产量。在模拟干旱条件下,火龙果组培苗和实生幼苗生长和生理指标都受到明显的影响[10-11]。【本研究切入点】在气候变暖背景下,干旱呈发生频率高、分布地域广、持续时间长、时空改变大等特点,严重影响农业生产[12]。实施精量灌溉是提高作物水分利用率和生产效率的重要途径之一[13]。而目前有关火龙果长期抗旱生长及生理响应的报道较少。【拟解决的关键问题】通过盆栽试验,研究长期干旱、复水交替环境下火龙果1年生扦插苗生长及生理指标的影响,以期为阐明火龙果抗旱机制、确定火龙果需水阈值提供参考,为科学制定抗旱策略,精准灌溉提供一定的数据支持。
1 材料与方法
1.1 试验地概况
试验在贵州省惠水县好花红镇贵州省农业科学院数字农业试验基地的温室中进行。试验区属亚热带季风气候,北纬26°17′45″,东经107°05′14″,试验地海拔956.0 m,年降水量1200~1300 mm,年均气温16.0 ℃,无霜期290 d。
1.2 材料
试验土壤为酸性黄壤(pH 5.50,有机质35.03 g/kg,全氮2.33 g/kg,碱解氮157.1 mg/kg,有效磷19.40 mg/kg,速效钾180.0 mg/kg,缓效钾227.0 mg/kg。火龙果品种为紫红龙,一年生红皮红肉火龙果扦插苗,主茎高50.5 cm。栽植容器为塑料盆,盆高38 cm,直径34 cm。
1.3 方法
1.3.1 试验设计 每盆装土13 kg,施入复合肥20 g(15-15-15),种植1株火龙果扦插苗,竹条支架固定,滴灌至充足水分。火龙果扦插苗种植4周后(4月1日起),模拟自然状态,进行降雨—干旱交替处理,设正常供水(CK,土壤含水量保持在田间持水量的80%)、轻度干旱胁迫(LD,土壤最低含水量为田间持水量的60%)、中度干旱胁迫(MD,土壤最低含水量为田间持水量的40%)和重度干旱胁迫(SD,土壤最低含水量为田间持水量的20%)4个处理,每个处理3盆,3次重复。各处理先浇足水分,利用TZS-5X土壤水分传感器(浙江托普公司生产)每3 d测量1次土壤含水量(10 cm深),当土壤水分降至临界值时行滴灌至水从盆底流出时结束。6月灌水时将尿素(N 46.0%,15 g/盆)溶于水滴灌,其他管理按常规。各处理的灌水量及灌水次数见表1。

表1 试验各处理的灌水量和灌水次数Table 1 Irrigation amount and times of different treatments
1.3.2 测定项目及方法 ①植株生长和生物量。试验开始后每间隔7 d测量新生茎蔓数量(以新芽长出5.0 cm计)和茎蔓长度(当茎蔓长度增长量连续3次测量低于0.5 cm即认为该枝条生长已达最大)。试验结束后,用游标卡尺测定一级分枝(从主茎上直接发出的侧枝)和二级分枝(从侧枝上发出的新枝)的棱厚(茎蔓中部的棱边缘至中央维管束之间中点的厚度)。将火龙果植株地上部分和根系称鲜重后,用刀片将火龙果茎蔓肉质皮层划开,与测定根系形态后的地下部分一起经105 ℃杀青30min,再80 ℃烘干至恒重,计算含水量和根冠比。②植株养分。植株烘干粉碎后,用H2SO4-H2O2消煮,氮用凯氏法,磷用钒钥黄比色法,钾用火焰光度计法测定。③根系形态特征。将根系装入70目的尼龙网袋中,浸泡30 min后用流水缓慢冲洗干净。将根系样本分解后放入装有水的专用无色透明根盘,用镊子调整根的位置尽量避免交叉重叠,用根系扫描仪(Epson Expression 11000XL,Epson company,日本,分辨率为300 dpi)对根系进行扫描后,用根系扫描分析系统(WinRHIZO Pro LA 2400,Regent Instruments,加拿大)测量根系总长度、根系总表面积、平均根系直径和总根体积。④气孔形态特征。剪下无任何表观损伤的成熟肉质茎,立刻用刀片将茎蔓切成4 mm×4 mm ×1 mm的组织块,将其放入盛有2.5%戊二醛的离心管中固定24 h,然后经PBS 缓冲液(0.1 mol/L,pH 6.8)清洗3次,分别用55%、65%、75%、85%、95%和100%的乙醇梯度脱水,每次脱水15 min。将脱水处理过的样品冷冻干燥后,用导电胶粘在样品台上,离子溅射仪镀金膜,然后用扫描电子显微镜(S3400,Hitachi,日本)进行观察和拍照。每个处理选取10个视野,用Image J软件计算气孔密度、气孔大小(长度×宽度)等。⑤生理指标。将采集的茎蔓样品经液氮(-196 ℃)处理后置于超低温冰箱(-80 ℃)中保存。采用苏州科铭生物技术有限公司试剂盒进行超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)、丙二醛(MDA)、脯氨酸(Pro)的测定;采用露点水势仪(WP4,Decagon,美国)测定火龙果茎蔓水势。以上指标均重复3次,取平均值。
1.4 数据分析
采用Microsoft Office Excel 2016对试验数据进行统计和计算,SPSS 20.0 软件进行方差分析和显著性测验,LSD法进行多重比较,显著水平为P≤0.05。
2 结果与分析
2.1 干旱胁迫对火龙果生长的影响
2.1.1 对地上部生长的影响 从表2可见,各干旱胁迫处理火龙果的新枝总长均低于对照(CK)其中,MD处理下降6.22%,差异不显著;MD和SD处理茎蔓生长受到严重抑制,分别比CK显著下降25.61%和66.82%。轻、中度干旱对新枝数和二级分枝棱厚无显著影响,但重度干旱使之显著降低;在轻度干旱处理中,一级分枝棱厚和地上部干重与CK间无显著差异,但中、重度干旱则显著降低。说明中度和重度干旱严重影响火龙果地上部分的正常生长。

表2 干旱对火龙果地上部分生长的影响Table 2 Effect of drought on the growth of above ground part of H. polyrhizus
2.1.2 对根系生长的影响 由表3可知,与CK相比,除平均根系直径外,随干旱胁迫的加重,火龙果的总根长、根系总表面积、总根体积和根干重均呈逐渐减少趋势。干旱对根系平均直径无显著影响,轻、中、重度干旱条件下降幅为1.09~1.18 mm。轻度干旱处理火龙果的总根长、根系总表面积、总根体积和根干重降幅未达显著水平,而中度和重度干旱火龙果的总根长、根系总表面积、总根体积和根干重均显著低于CK。表明中度和重度干旱严重影响火龙果根系的生长和扩展。

表3 干旱对火龙果根系性状的影响Table 3 Effect of drought on root characteristic of H. polyrhizus
2.1.3 对火龙果茎气孔参数的影响 从图2和表4可知,气孔形态和气孔参数对干旱胁迫有不同程度的响应。轻、中度干旱胁迫下气孔形态变化不明显;重度干旱胁迫下气孔形态发生显著变化,气孔明显下陷,周围分布较多的针晶或棒状晶体。重度干旱胁迫下气孔密度显著增加,而外部气孔长度、宽度、气孔大小、内部气孔长宽、气孔孔径均显著减小;轻、中度干旱胁迫下气孔密度、外部气孔长度和内部气孔长度在变化不明显,但气孔外宽、内宽和气孔孔径均显著小于对照。轻度干旱胁迫下气孔大小与对照无显著差异,中度干旱胁迫下气孔大小与对照有显著差异。
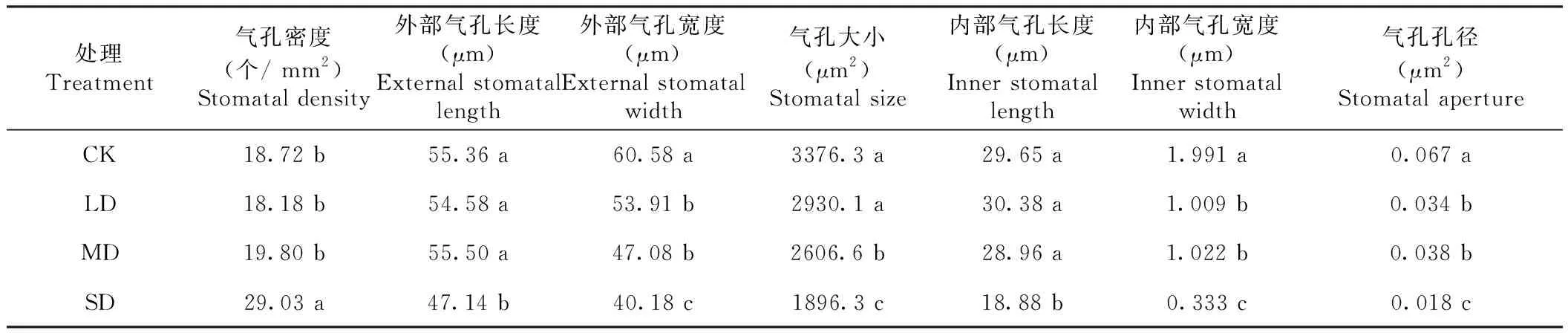
表4 不同干旱胁迫处理火龙果的茎蔓表皮气孔参数Table 4 Stomatal parameter of H. polyrhizu stem and epidermis treated with different drought levels
2.2 干旱胁迫对火龙果植株养分吸收的影响
由表5可知,植株养分含量中, N含量随着干旱胁迫的加深呈先增加后降低趋势,各干旱处理均高于对照,除轻度干旱处理显著高于CK外,其余处理与CK间差异不显著;植株P含量和K含量随着干旱胁迫的加深均呈增加—降低—增加的趋势,P、K含量除中度干旱低于CK外,轻度和重度干旱处理均显著高于CK;说明轻度干旱有利于氮和磷吸收,钾含量则是随着干旱程度增加整体显著升高,与其他处理间差异显著。火龙果植株氮、磷、钾吸收量以轻度干旱处理(LD)的最高与其他处理间差异显著;其次是中度干旱处理(MD),与CK间差异不显著,其中氮、钾吸收量显著高于重度干旱处理(LD),磷吸收量与LD处理差异不显著;重度干旱处理(SD)火龙果的氮、磷、钾吸收量均最低比LD处理分别低21.23%、4.17%和17.55%。

表5 不同干旱胁迫处理火龙果的养分含量和吸收量Table 5 Nutrient content and absorption of H. polyrhizus treated with different drought levels
2.3 干旱对火龙果含水量和抗旱生理指标的影响
2.3.1 含水量和水势 由表6可见,一级分枝和二级分枝含水量均以轻度干旱胁迫(LD)处理的最高,其中,一级分枝各处理含水量表现为LD≥CK≥MD≥SD,二级分枝为LD>CK>MD>SD,说明轻度干旱(LD)处理有利于火龙果茎储存水分;随旱情加重,含水量呈下降趋势,表现出明显的缺水现象;与CK相比,中度缺水(MD)的一、二级分枝含水量虽有降低但无显著差异,但重度缺水的二级分枝与MD已存在显著差异,说明火龙果虽是耐旱植物,但严重干旱条件下新枝的水分蓄积受影响。一、二级分枝的水势前LD、CK和MD处理间无显著差异,但SD处理的水势明显降低,与CK相比,一、二级分枝的水势分别降低31.87%和13.31%。

A、B、C、D分别为标尺1000 SE 下:CK, LD, MD, SD的气孔器形态;a、b、c、d分别为标尺100 SE下CK, LD, MD, SD气孔整体面观A. B, C and D were stomatal apparatus morphology of CK, LD, MD and SD under 1000 SE scale; a,b,c and d were the overall view of H. polyrhizu stomatas of CK, LD, MD and SD under 100 SE scale图2 扫描电镜下火龙果茎蔓表皮气孔0: 00的形态Fig.2 Stomatal micro-morphological features of stem and epidermis of H. polyrhizu at 0: 00 under SEM
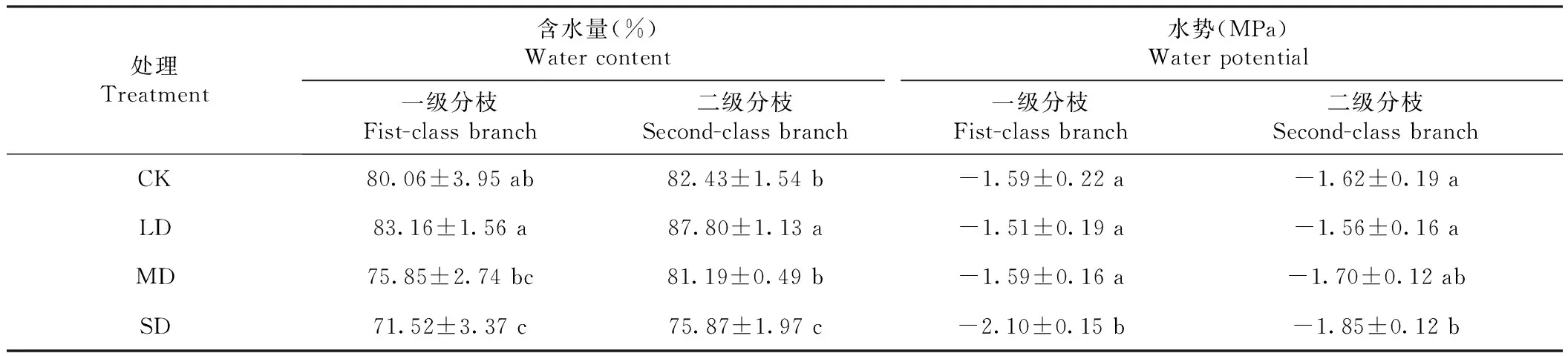
表6 不同干旱胁迫处理火龙果茎蔓的含水量和水势Table 6 Water content and water potential of H. polyrhizus stem and epidermis treated with different drought levels
2.3.2 抗氧化酶活性 由表7可见,4个处理中,火龙果茎蔓的SOD和POD活性表现为MD>LD>CK>SD,说明轻度干旱对抗氧化指标无影响,中度干旱启动了SOD和POD抗氧化保护机制,其酶的活性显著升高,但在重度干旱胁迫下,SOD和POD活性明显降低。CAT表现出在重度干旱条件下显著升高,其他处理无差异。说明抗氧化酶系对干旱程度的响应存在差别。干旱胁迫处理丙二醛(MDA)含量均显著增加,但各干旱处理间无显著差异,MDA含量表现为MD>SD>LD>CK,与CK相比,增幅35.31%~56.60%。各处理脯氨酸(Pro)含量表现为MD>SD>LD>CK,中度干旱脯氨酸含量显著增加,但严重干旱又下降,与对照相比,增幅为8.95%~113.11%。

表7 干旱胁迫下火龙果的抗氧化酶活性变化Table 7 Changes in activities of antioxidant enzymes in H. polyrhizus under drought stress
3 讨 论
季节性干旱是农业生产的主要限制因子之一,其微观上抑制细胞生长,宏观上降低植株的生长速率[14]。因此,生物量和产量的变化是植株对干旱胁迫最直观的反映。火龙果是贵州省干热河谷新兴的主栽经济作物,在植株一年的生长过程中,5月中旬后生长速率逐渐加快,6月初的新生枝条生长速率达最大值,前期的营养生长为后期的生殖生长奠定基础。本研究发现,在充足供水条件下,5—6月火龙果茎枝生长速率很快,类似前人研究[15-16]。中、重度干旱胁迫下,火龙果的新枝总长、一级分枝棱厚及地上部干重显著下降,而轻度干旱的火龙果地上部分干重和一级分枝棱厚均为4个处理的最大值,可能轻度干旱有益于火龙果的生长。对于重度干旱处理(土壤含水量为田间持水量的20%),普通植物一般都难于忍受,但火龙果却未出现死亡现象,说明火龙果抗旱性极强,适宜种植在西南干热河谷地区,但中、重度干旱显著抑制火龙果的正常生长,必然影响后期的生殖生长和果实产量。因此,在干热河谷种植火龙果的过程中,科学灌水可促进火龙果营养生长、茎发育及干物质积累,既能搭建植株骨架,也有利于增加结果量。
植物根系是吸收水分和养分的重要器官。随旱情加重,火龙果根系总长度、总表面积和总体积逐渐降低。但值得注意的是,在中度干旱条件下,火龙果根系总长度、总表面积和总体积的降幅达显著水平(与正常供水相比),而只有在重度干旱条件下,火龙果新枝数、新枝长度和生物量等与生长相关的参数才显著降低。此外,火龙果根系生物量仅占总生物量的极少部分(3.22%~4.71%),远低于一般植物[17-19],说明火龙果抗旱能力强非根系原因,而与地上部的生物学形态、水分保持和生理代谢等密切相关。
气孔是CO2固定的主要通道,在干旱条件下,植物为保持水分减少气孔开放程度,增加气孔密度,是适应干旱胁迫环境的常用策略[20]。然而,气孔导度的降低也限制了CO2的固定量,导致光合速率受到抑制和同化减少,并且在形态上表现为生长抑制和产量损失[21]。轻度干旱胁迫下,细胞伸长受到抑制,导致叶面积减少,单位面积气孔数增加,气孔密度增加;严重干旱胁迫下,叶片受到严重伤害,气孔数量显著减少,气孔密度随之降低[22-23]。本研究中,中度干旱胁迫已引起气孔大小显著减小,推测这是影响光合作用,导致生物量下降的原因之一;严重干旱胁迫下,茎表皮气孔周围出现大量针晶,含晶异细胞的形成与干旱环境有关,可以提高植物的吸水和保水能力[24],使火龙果具有强抗旱能力。

火龙果茎由表皮、皮层、维管束和髓组成。其中表皮为多层细胞组成的复表皮,皮层由外部的栅栏组织和内层的储水组织、粘液细胞组成[32]。随干旱胁迫程度的增加,茎表皮厚度、储水细胞数量增加,皮层及茎总厚度减小[4]。尽管干旱使火龙果含水量和细胞水势降低,但降幅远低于一般植物[33-34]。在干旱条件下,较高的含水量和细胞水势有益于维持光合作用和新陈代谢,并减轻伤害[35]。因此,从生理指标看,适度干旱可提高火龙果新枝SOD、POD和CAT的活性。在植物体内,SOD、POD、CAT可清除干旱产生的活性氧,避免膜脂过氧化[36]。干旱使植物体内的细胞膜发生膜过氧化,产生丙二醛[37-38];脯氨酸积累是干旱伤害程度的重要指标之一,伤害愈重脯氨酸含量越高,可增加几十甚至上百倍[39-40]。尽管干旱使火龙果新枝丙二醛和脯氨酸含量增加,但其增幅远低于普通植物[41-42]。因此,在干旱条件下,火龙果抗氧化酶活性提高,伤害程度减轻可解释其抗旱较强的主要原因。今后将结合火龙果气孔运动、光合作用等生理特征的研究,进一步开展火龙果蒸腾规律的探索,以期为火龙果的节水灌溉提供科学依据。
4 结 论
研究发现,水分供应水平显著影响火龙果苗的新梢生长速率、生物量、根系形态和抗性生理指标。火龙果具有较强的抗旱性,适宜种植于贵州省季节性干旱严重、土层浅薄的干热河谷地区。轻度干旱胁迫下火龙果的生物量略高于正常供水处理,中、重度干旱显著影响火龙果苗的新梢生长速率、生物量、根系形态和酶活性,说明间歇性轻度干旱有利于火龙果的茎蔓生长,长期重度干旱抑制其正常生长。生产中,控制土壤相对含水量为60%左右有利于火龙果树体的生长发育。
猜你喜欢
果树实用技术与信息(2022年6期)2022-11-21
航空材料学报(2022年4期)2022-08-04
贵州农业科学(2022年4期)2022-05-06
干旱地区农业研究(2022年1期)2022-01-28
语文周报·教研版(2021年28期)2021-08-19
农民致富之友(2020年33期)2020-12-03
安徽农业科学(2020年18期)2020-10-09
电脑报(2020年12期)2020-06-30
科技创新与品牌(2019年12期)2019-02-06
农民致富之友(2017年15期)2017-08-20
