
叶面喷施钙、镁肥对‘妃子笑’荔枝果肉有机酸跨膜运输的影响
2022-08-06 08:57廖海枝林晓凯杨成坤杜婧加彭俊杰周开兵
西南农业学报 2022年6期
廖海枝,林晓凯,杨成坤,杜婧加,彭俊杰,周开兵
(1. 热带作物新品种选育教育部工程研究中心,海口 570228; 2. 海南大学园艺学院, 海口 570228)
【研究意义】荔枝(LitchichinensisSonn.)为无患子科常绿乔木,与香蕉、菠萝、龙眼一同号称“南国四大果品”,其在华南区具有重要的经济和社会效益[1]。其中,‘妃子笑’荔枝(LitchichinensisSonn. cv. Feizixiao)是我国荔枝主栽品种之一,是海南产区栽培面积最大和总产量最高的品种[2],其提质增效栽培是海南省荔枝产业可持续发展的必然途径,因此必须开展荔枝果实品质发育研究。‘妃子笑’荔枝果肉富含苹果酸、酒石酸、草酸、柠檬酸等多种有机酸组分[3],荔枝果实风味品质正是由糖和有机酸的组分与含量影响的[4],然而植物果实有机酸的积累与有机酸的跨膜运输有关[5],目前关于‘妃子笑’荔枝果实的有机酸代谢及其生理机制问题的研究报道较少,因此有必要着手研究荔枝果肉有机酸跨膜运输问题,以期为荔枝的提质增效栽培提供理论依据。【前人研究进展】植物果实有机酸代谢很复杂,有机酸参与糖酵解反应、三羧酸循环、糖异生作用等途径[6]。有机酸在代谢中也存在液泡膜的跨膜运输问题,其关键酶主要包括ATP酶(H+-ATPase)和焦磷酸酶(V-PPase)等两个酶[7]。植物液泡膜H+-ATPase和液泡膜V-PPase是液泡膜上含量丰富的两个蛋白,在植物的正常生长和胁迫条件下起到重要作用[8-9]。前人报道,液泡膜H+-ATPase和V-PPase等2种类型的质子泵会影响果实液泡有机酸的贮藏[9]。跨膜运输和分解可能会影响柠檬酸积累[10]。柠檬酸的转运需要质子流的流动,液泡膜上的的H+-ATPase和V-PPase负责调节,保持液泡内的酸度pH,防止液泡被碱化[11]。【本研究切入点】果实中的质子泵基因密切影响着有机酸含量,尤其是苹果酸和柠檬酸[12]。有机酸转运过程可能影响有机酸含量,此外,营养元素如钾、氮、磷等也会对果实酸度有较大影响[12-13]。【拟解决的关键问题】基于课题组前期已明确叶面喷钙镁混合肥处理能缓解妃子笑荔枝果肉“退糖”现象[14],本文通过在‘妃子笑’荔枝树冠上喷施钙、镁肥对其进行处理来探讨不同处理是否对‘妃子笑’荔枝果实有机酸跨膜运输存在影响,进而了解是否引起有机酸积累而影响果肉呼吸作用,从而缓解果肉“退糖”现象,也为果实有机酸代谢研究提供一定理论支持。
1 材料与方法
1.1 材料
试验地点位于海南省临高县金牌农场五队荔枝园,地形为琼北台地,属于热带季风气候区,年平均温23~24 ℃,年平均日照时数2175 h,年平均降雨量1100~1800 mm,且雨热同季;土壤为肥沃砖红壤。在该果园选取长势一致、无病虫害的20株16年生‘妃子笑’荔枝树作为试验材料。花期为2—3月,生理落果期为4月初开始,4月下旬进入果实膨大期,5月中旬进入果实成熟期,试验于4月下旬开始进行叶面喷肥处理。
1.2 试验设计方法
对树冠进行叶面喷肥处理,即以叶面喷施0.3%氯化钙水溶液(Ca)、0.3%氯化镁水溶液(Mg)、0.3%氯化钙和0.3%氯化镁混合水溶液(Ca+Mg)等为处理,以叶面喷施同等体积的清水为对照(CK)。单株小区,5次重复。
1.3 取样与样品处理方法
在第一次处理前,先在树冠中部外围四方选择生长健壮、大小中等的果实作为以后取样的参照果,并挂牌标记。叶面喷肥处理时间分别于谢花后35、42、50 d(9: 00—10: 00 am),每次施肥处理前对各单株按照取样参照果大小取好样果;此后分别继续于谢花期后56、63、69和73 d对各单株分别按照取样参照果大小、着色状态取果样;每次对各单株取样果30个,取好样果后立刻速冻于液氮罐,并于实验室超低温冰箱(-80 ℃)中储存。
1.4 试验指标及其测定方法
有机酸含量测定:有机酸标样包括苹果酸、酒石酸、柠檬酸、草酸和琥珀酸等,由北京坛墨质检科技有限公司出产。参考胡志群等[15]与王芮东[16]等方法并略有改动,采用高效液相色谱法鉴定和检测上述单体有机酸种类和含量。称取1 g荔枝果肉,用0.2%的偏磷酸冰浴研磨,超声20 min后10 000 r/min离心15 min,使用滤膜(0.45 μm)过滤上清液后待测。流动相为用磷酸调节pH至2.8的0.1%的磷酸二氢钠溶液,流速为0.6 mL/min,柱温为室温,波长210 nm,进样量10 μm。
跨膜运输酶活性测定:使用江苏科特生物公司试剂盒测定H+-ATPase和V-PPase两种跨膜运输酶活性,依据产品说明书介绍的双抗体夹心法进行测定。具体方法参考马俊兴等人在牦牛肺上的研究方法[17]。
水溶性钙、镁含量测定:称取1 g荔枝果肉,烘干至恒重,加入去离子水研磨至匀浆,震荡过夜后采用火焰原子吸收法测定水溶性钙、水溶性镁含量[18]。
使用仪器:配有VWD紫外检测器的高效液相色谱仪(型号:ALLianceE26954),5 μm色谱柱(型号:SinoChromODS-BP,Nucleodur 250 mm×4.6 mm),原子吸收分光光度计(型号:NOVAA400P),离心机,酶标仪,震荡培养箱等。
1.5 数据统计分析方法
采用SAS软件进行数据统计分析,分别采用ANOVA过程、DUNCAN法和REG过程作方差分析、多重比较分析、多元线性相关分析。
2 结果与分析
2.1 有机酸含量动态变化
2.1.1 草酸 不同处理和CK的草酸含量动态变化趋势和同一时间点不同处理之间均具有明显的差异(图1-A)。CK、Mg和Ca+Mg处理经历两次“上升、下降”交替过程,随后CK和Ca+Mg处理再次上升,之后CK维持稳定,Ca+Mg处理则下降;Ca处理表现为“上升、下降、平缓、上升、下降”的变化趋势。在花后42 d,Ca处理显著最高,Ca+Mg处理显著低于CK;在花后50 d,Mg和Ca处理显著低于CK;在花后56 d,Ca和Ca+Mg处理显著低于CK;在花后63 d,CK和Ca+Mg处理显著低于Mg和Ca处理,Ca处理显著低于Mg处理;在花后69 d,CK和Mg处理显著低于Ca和Ca+Mg处理,Ca处理显著低于Ca+Mg处理;在花后73 d,Mg处理显著最低。可见,CK果肉草酸含量在前期处于较高水平,在后期则低于Ca、Mg和Ca+Mg处理,说明不同处理在前期抑制果肉草酸积累,在后期则反之。
2.1.2 酒石酸 不同处理和CK的酒石酸含量动态变化趋势基本一致,在某些时间点不同处理间存在明显差异(图1-B)。不同处理和CK在初始时相对稳定,在花后50 d迅速上升后下降。在花后42 d,Mg处理显著最低,其余处理与CK间均无显著差异;在花后56 d,Ca处理显著低于Ca+Mg处理;在花后69 d,CK和Mg处理显著低于Ca和Ca+Mg处理;在花后73 d,Mg处理显著低于Ca处理;除上述显著差异外,不同处理和CK在果肉酒石酸含量上均无显著差异。可见,在果肉酒石酸含量上,CK呈低于Ca和Ca+Mg处理趋势,而呈高于Mg处理趋势,说明Ca和Ca+Mg处理呈促进酒石酸积累的趋势,而Mg处理则反之。
2.1.3 苹果酸 不同处理和CK的果肉苹果酸含量具有一致的“L”型动态变化趋势,均先急剧下降至花后50 d,随后趋于稳定;在某些时间点不同处理间则存在显著差异(图2-A)。在花后42 d,CK显著最低,Ca处理显著低于Mg处理;在花后73 d,CK和Mg处理显著低于Ca处理;除上述显著差异外,不同处理和CK间均无显著差异。可见,前期的所有处理呈促进苹果酸积累的趋势,仅Ca处理促进后期苹果酸的积累。
2.1.4 柠檬酸 不同处理和CK的柠檬酸含量的动态变化均呈“M”型趋势,在同一时间点不同处理间则存在明显差异(图2-B)。在花后42 d,Ca处理显著最低,CK和Mg处理显著高于Ca+Mg处理;在花后50~63 d,CK均最低,除了在56 d时与Ca处理无明显差异外,均显著低于其余3个处理;在花后50和56 d,Ca处理显著低于Mg和Ca+Mg处理;在花后63 d,Ca+Mg处理显著低于Ca和Mg处理;在花后69 d,CK显著低于Ca+Mg处理,Mg处理明显最低;在花后73 d,Ca+Mg处理显著最高,CK和Mg处理明显低于Ca处理。可见,CK持续处于较低水平,Ca和Ca+Mg处理可提高柠檬酸含量,Mg处理则呈先促进、后抑制的趋势。

图2 不同处理下苹果酸和柠檬酸的含量变化Fig.2 The content changes of malic acid and citric acid under different treatments
2.2 有机酸跨膜运输酶活性动态变化
2.2.1 H+-ATPaseV 不同处理和CK的果肉H+-ATPase活性具有不同的变化趋势,且在同一时间点不同处理间也存在明显差异(图3-A)。CK经历3次的“下降、上升”交替;Mg处理经历3次“上升、下降”交替;Ca处理先无明显变化,随后呈“下降、上升、平缓”的趋势;Ca+Mg处理先下降,随后保持稳定,再经历2次“上升、下降”交替过程。在花后42 d,CK和Ca+Mg处理显著低于Mg和Ca处理;在花后50 d,CK显著最高;在花后56 d,Ca处理显著低于CK和其余处理;在花后63 d,CK、Ca、Mg、Ca+Mg处理依次降低,且任意两者间均显著;在花后73 d,亦为任意两者间均显著,由高至低排序为Ca、Ca+Mg、Mg处理。可见,CK的高峰与其余处理相反,且69~73 d,唯有CK显著上升,表明不同处理呈抑制果肉H+-ATPase活性的趋势。
2.2.2 V-P Pase酶 不同处理和CK的果肉V-PPase酶活性具有不同的变化趋势,在同一时间点不同处理间也存在明显差异(图3-B)。CK和Ca处理经历两次“下降、上升”交替过程后又下降;Mg和Ca+Mg处理呈“下降、上升、下降、平缓”趋势。在花后42 d,Ca+Mg处理显著最低;在花后50、63、69 d,CK均显著高于其余处理,在50 d时,按CK、Ca、Mg、Ca+Mg处理排序依次降低,且任意两者间均显著,在63 d时,Ca+Mg处理显著高于Ca处理,69 d时,Ca处理显著高于Mg和Ca+Mg处理;在花后56 d,Mg和CK显著高于Ca和Ca+Mg处理,Mg处理又显著高于CK;在花后73 d,CK和Ca处理显著高于Mg和Ca+Mg处理。可见,CK除在56 d时低于Mg处理外,全程高于其余处理,说明所有处理呈抑制果肉的V-PPase酶活性的趋势。
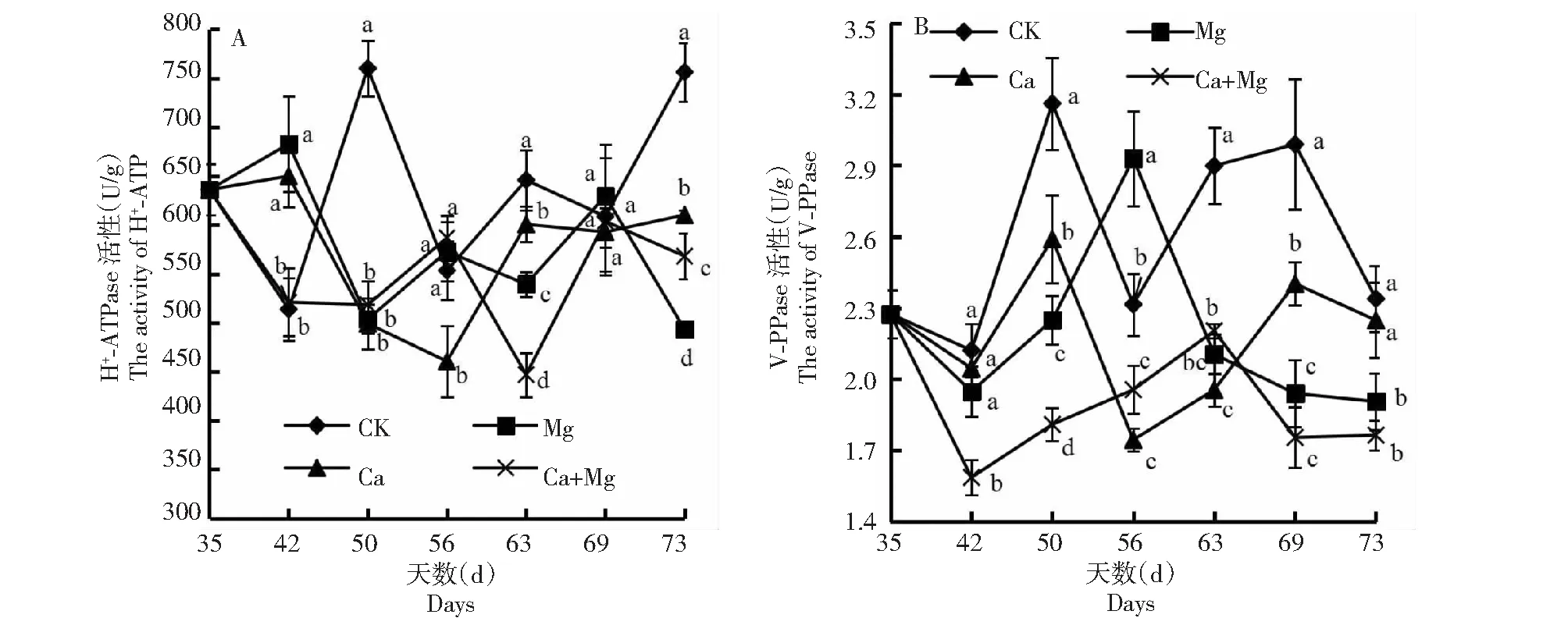
图3 不同处理下H+-ATP和V-PPase的活性动态变化Fig.3 Dynamic changes of H+-ATP and V-PPase activities under different treatments
2.3 水溶性钙、镁含量动态变化
2.3.1 水溶性钙 不同处理和CK的水溶性钙含量在动态变化趋势和同一时间点上均具有明显差异(图4-A)。花后63 d前不同处理和CK的水溶性钙含量均上升,随后CK和Ca处理持续上升,Mg处理继续上升后下降,Ca+Mg处理保持平缓。在花后42 d,各处理间均差异显著,排序从高到低为Ca、Mg、CK、Ca+Mg处理;在花后50~69 d,除63 d外,Mg处理均为显著最高;在花后50和56 d,CK和Ca处理均显著高于Ca+Mg处理;在花后63和73 d,分别为Ca和Ca+Mg处理显著最低。可见,Mg处理高于CK和其余处理,Ca+Mg处理则持续低于CK和Mg处理,表明Mg处理表现出促进水溶性钙含量积累的趋势,Ca+Mg处理则反之,CK、Mg和Ca+Mg处理在73 d后被Ca处理超越。
2.3.2 水溶性镁 不同处理和CK的水溶性镁含量的动态变化趋势和同一时间点均差异明显(图4-B)。所有处理和CK先上升再下降,随后CK维持稳定,Mg处理上升再下降,Ca处理持续上升,Ca+Mg处理保持稳定后降低。在花后42 d,CK显著高于Mg、Ca和Ca+Mg处理,Mg和Ca处理均显著高于Ca+Mg处理;在花后50~56 d,CK和Ca+Mg处理均显著低于Ca处理;在花后50 d,Ca处理还显著低于Mg处理;在花后63 d,Mg和Ca处理显著低于CK和Ca+Mg处理;在花后73 d,Ca处理显著最高,CK显著高于Ca+Mg处理。可见,水溶性镁含量会被Mg和Ca处理所提高,Ca+Mg处理则反之。

图4 不同处理下水溶性钙和水溶性镁的含量动态变化Fig.4 The content changes of water-soluble calcium and water-soluble magnesium under different treatments
2.4 有关指标间多元线性相关性分析
2.4.1 不同有机酸含量与两种酶活性 由表1可见,CK的H+-ATPase活性与草酸、苹果酸、柠檬酸等含量的偏相关系数呈负相关;Mg处理H+-ATPase活性与苹果酸含量的偏相关系数呈正相关;Ca处理的H+-ATPase活性与4种有机酸含量的偏相关系数均呈正相关,V-PPase活性与草酸含量的偏相关系数呈负相关;Ca+Mg处理的H+-ATPase活性与草酸、酒石酸、柠檬酸含量的偏相关系数分别呈正、负、负相关,V-PPase酶活性与草酸含量的偏相关系数呈负相关,与酒石酸、苹果酸等含量的偏相关系数呈正相关。表明不同施肥处理会改变有机酸含量与两种跨膜运输酶活性间的线性相关性,说明叶面喷施钙、镁营养通过影响跨膜运输酶活性与有机酸含量的线性相关性而影响有机酸含量。

表1 不同处理和CK四种有机酸含量与两种酶活性的显著偏相关系数Table 1 The significant partial correlation coefficients between the contents of four organic acids and two enzymes under different treatments and CK
由表2可见,除Mg处理的草酸含量外,CK和其余处理的草酸含量与两种酶活性的复相关系数均显著或极显著;Mg和Ca处理的苹果酸含量、CK和Ca+Mg处理的柠檬酸含量分别与两种酶活性达到显著或极显著水平。这说明草酸、苹果酸、柠檬酸等3种有机酸的积累均受两种跨膜运输酶活性变化的影响,任意一种酶活性改变均会引起不同有机酸含量的改变,说明不同处理可能通过影响这些酶活性而影响有机酸的跨膜运输,进而影响有机酸含量。
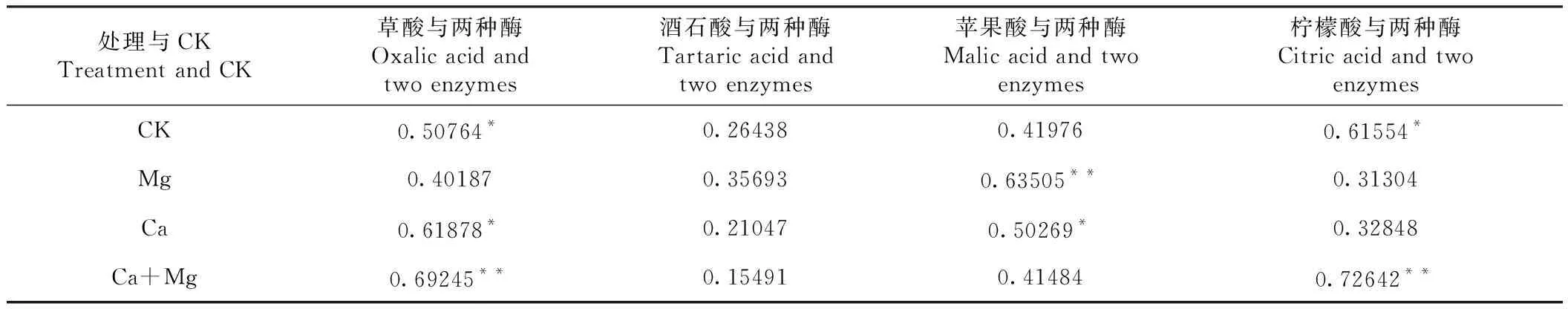
表2 不同处理和CK四种有机酸含量分别与两种酶活性的复相关系数Table 2 The multiple correlation coefficients between the contents of four organic acids with two enzymes respectively under different treatments and CK
2.4.2 水溶性钙、镁含量与两种酶活性 由表3可见,H+-ATPase活性与水溶性钙、镁含量的偏相关系数均为负相关,且仅Mg和Ca处理存在相关性;而Ca和Ca+Mg处理的V-PPase活性与水溶性Mg含量分别呈正、负相关,且叶面喷施钙、镁肥相对于CK均改变了水溶性钙、镁含量与两种酶活性的线性相关性。可见,水溶性钙、镁积累对H+-ATPase、V-PPase活性呈抑制趋势,且不同的施肥处理可能通过对两种跨膜运输酶活性产生不同的调节作用而调节跨膜运输能力,进而调节有机酸的积累。

表3 不同处理水溶性钙、镁与相关酶的显著偏相关系数Table 3 The significant partial correlation coefficients between the contents of the water-soluble Ca and Mg and related enzymes under different treatments
由表4可见,仅Mg处理的水溶性钙含量和Ca处理的水溶性镁含量与两种跨膜运输酶的复相关系数具有显著性,表明叶面喷施钙、镁肥分别可能通过影响水溶性镁、钙含量而影响两种酶活性,进而影响有机酸跨膜运输能力,从而最终影响有机酸的积累。

表4 不同处理和CK水溶性Ca、Mg含量分别与两种酶活性的复相关系数Table 4 The multiple correlation coefficients between the contents of the water-soluble Ca or Mg with two enzymes respectively under different treatments and CK
3 讨 论
荔枝果实苹果酸在成熟前含量为下降趋势,酒石酸前期增加成熟后减少,柠檬酸在后期基本检测不到[19],本文结果与此结果基本相同,但与枣[20]、李[21]、菠萝[22]等果实中的各种有机酸含量变化并非一致,这可能与果树种类不同有关。叶面喷施钙、镁肥会影响有机酸含量,其中,Ca和Ca+Mg处理呈促进有机酸积累的趋势,而Mg处理则可能反之。又由于作为呼吸代谢中间产物的有机酸积累会引起果肉呼吸的负反馈调节[22-23],于是导致Ca和Ca+Mg处理果肉呼吸代谢在后期较弱,而Mg处理则相反,从而产生课题组前期研究结果,即Ca和Ca+Mg处理后期积累糖分较多,从而缓解果肉“退糖”现象;而Mg处理后期积累糖分较少,从而与CK一样出现“退糖”现象[25]。
前人报道在甘蓝上钙、镁营养元素的吸收存在协同关系[26],笔者前期研究结果也表明水溶性钙和镁存在相互增益效应[27],而在猕猴桃叶片上Ca和Mg却相互抑制吸收[28],可见,水溶性钙、镁积累问题受遗传差异影响。本文结果表明,单独喷施钙或镁营养分别会促进果肉中水溶性钙、镁积累增多,即水溶性钙、镁存在协同作用,与前人研究一致。
有机酸代谢受跨膜运输酶V-PPase和H+-ATPase的影响,它们都是具有质子泵功能的与液泡膜结合的水解酶[29],可以催化H+从细胞质转移到液泡腔,产生酸性内pH值,对果实酸度调节具有重要作用[30]。本文结果表明,叶面喷施Ca、Ca+Mg会抑制两种跨膜运输酶活性,并将CK表现出的跨膜运输酶活性与有机酸含量的负相关改变为正相关,这意味着叶面喷施Ca、Ca+Mg减弱了有机酸跨膜运输能力[31],致使糖酵解、三羧酸循环等形成的有机酸积累于细胞质、线粒体,推测叶面喷施Ca、Ca+Mg后果肉有机酸并未象CK那样转运进液泡[32],而是主要积累于液泡外细胞质中和线粒体,从而负反馈调节果肉呼吸作用[23-24],导致在成熟期减少糖分的呼吸损耗,这进一步解释了叶面喷Ca、Ca+Mg缓解果肉成熟期“退糖”现象的生理生化原因。

4 结 论
H+-ATPase和V-PPase两种跨膜运输酶会影响‘妃子笑’荔枝果肉有机酸主要组分草酸、酒石酸、苹果酸、柠檬酸等含量;叶面喷施Ca、Mg处理分别通过增加水溶性镁、钙含量而减弱H+-ATPase活性,进而减弱细胞质中有机酸跨膜运输进入液泡的能力,最终可能增加有机酸在液泡外细胞质中的积累。叶面喷Ca、Ca+Mg处理可能通过阻止有机酸跨膜运输进入液泡而积累于液泡外细胞质中,从而提高果肉总酸含量和调节果肉风味营养品质,并抑制果肉呼吸作用而缓解果肉成熟期“退糖”现象。
猜你喜欢
作文新天地(小学版)(2020年3期)2020-04-10
当代旅游(2019年3期)2019-09-19
作文评点报·小学五、六年级(2018年30期)2018-05-18
中学语文(学生版)(2018年11期)2018-03-23
金点子生意(2017年9期)2017-09-13
科学与财富(2016年34期)2017-03-23
江苏农业科学(2016年7期)2016-10-20
农村农业农民·B版(2014年12期)2015-01-04
科技与创新(2014年17期)2014-10-22
晚晴(2014年9期)2014-09-19
