
适宜隧道发酵栽培草菇的培养料配方筛选试验
2022-08-05 08:19:56林金盛曲绍轩李辉平李宏斌顾鲁同
食用菌 2022年4期
蒋 宁 林金盛 曲绍轩 李辉平 骆 昕 徐 平 李宏斌 马 林* 顾鲁同
(1江苏省农业科学院蔬菜研究所/江苏省高效园艺作物遗传改良重点实验室,江苏南京 210014;2泗洪绿康食用菌科技有限公司,江苏宿迁 223942;3 阳泉职业技术学院,山西阳泉 045000;4江苏省农业技术推广总站,江苏南京 210036)
草菇Volvariella volvacea,隶属于担子菌纲Basidiomycetes 伞菌目Agaricales 小苞脚菇属Volvariella[1],是一种高温型食用菌,具有较高的保健价值,在我国南方地区和东南亚国家栽培较广[2]。据中国食用菌协会统计,2019 年中国草菇总产量为23.43万t。
草菇栽培常用的原料有棉籽壳、废棉渣、稻草、中药渣等,近年来利用金针菇、杏鲍菇等木腐类食用菌菌渣栽培草菇量不断上升[3]。草菇规模化栽培分几个阶段,首先从20 世纪60 年代的室外生料堆式栽培,到80 年代后期的保温房床架式栽培,之后进入21 世纪实现了机械化、周年化栽培,再到目前开始探索全方位的工厂化栽培阶段[4]。
草菇工厂化栽培,通过隧道发酵有效降解培养料是实现草菇高产的关键。在同样需要经过培养料隧道发酵过程的双孢蘑菇Agaricus bisporus生产中,培养料木质纤维素的降解率与其酶活性呈正相关[5]。利用不同基质栽培大球盖菇Stropharia rugosoannulata时,胞外纤维素酶、半纤维素酶、漆酶等胞外酶活性均存在明显差异[6]。已有文献报道,菌渣中加入稻草或麦草有利于草菇培养料的发酵[7],但未对最适比例进行深入研究,尤其是适宜隧道发酵的最佳配方鲜有报道。培养料配方对隧道发酵过程中基质胞外酶、产量等都有影响。因此,笔者从草菇生理生化的角度出发,以杏鲍菇菌渣和麦草培养料为研究对象,探究隧道发酵过程中草菇基质的pH 和部分胞外酶活性的变化,以期为稳定草菇产量,优化生产条件,进一步推进草菇工厂化栽培提供基础数据支撑。
1 材料与方法
1.1 试验材料
草菇菌种:V23,江苏省农业科学院蔬菜研究所提供。
培养料配方1:杏鲍菇菌渣100%;配方2:杏鲍菇菌渣90%,麦草10%;配方3:杏鲍菇菌渣80%,麦草20%;配方4:杏鲍菇菌渣70%,麦草30%。杏鲍菇菌渣的原始配方为杂木屑25%,玉米芯20%,甘蔗渣15%,麸皮25%,玉米粉5%,豆粕粉7%,轻质碳酸钙2%,石灰1%。
1.2 试验方法
1.2.1 发酵前准备
杏鲍菇采收结束后,挑选无杂菌感染的菌包,用脱袋机去除塑料袋,并将菌渣粉碎成直径小于5 mm 的颗粒物;将麦草切割成长度20 cm 左右的小段。菌渣与麦草按比例备好,分别加入4%的熟石灰,混合均匀后准备发酵。
1.2.2 培养料隧道发酵
将混匀备用的培养料放入一次发酵隧道中,按照8 min/20 min的通风频率持续发酵2 d。一次发酵结束后将培养料转入封闭的二次发酵隧道,加大通风量将培养料料温降至45 ℃(空间温度为43 ℃),随后降低通风量,保持每小时1~1.5 ℃的速度增温,至料温达到60 ℃(空间温度为58 ℃),恒温保持10 h;加大通风量降温8 h,料温降至50 ℃(空间温度为48 ℃),保持通风2~3 d,待氨气下降到5 mg/L 以下,温度40 ℃以下时培养料隧道发酵结束。
1.2.3 样品采集
分别在一次隧道发酵前(A)、二次隧道发酵前(B)和二次隧道发酵结束后(C)采集培养料样品。采用随机取样法,每处理取5 个点采集50 g,混匀,三次重复。每重复分两份样品:一份用于测定pH,另一份用于测定胞外酶活性。
1.2.4 培养料pH测定
称取通过2 mm 孔径筛的供试样品10 g,按水样比2.5∶1加入去除CO2的蒸馏水,将样品充分搅拌分散,放置30 min后采用玻璃电极法测定样品pH[8]。
1.2.5 粗酶液的提取
称取10 mg 供试样品,加入100µL pH 7.2~7.4的磷酸缓冲盐溶液(phosphate buffer saline,PBS),充分混匀。4 ℃、3 000 r/min 离心20 min,收集上清即为粗酶液。
1.2.6 胞外酶活性测定
采用双抗体夹心法酶联免疫吸附试验试剂盒法,分别按照上海茁彩生物科技有限公司微生物总蛋白酶(t-Pro)、半纤维素酶(Hce)、纤维素酶(CE)和木质素过氧化物酶(LIP)ELISA 检测试剂盒说明书操作,Rayto RT-6100 酶标分析仪在450 nm 波长处测定各处理的OD 值,根据说明书计算公式计算酶活性。
1.2.7 草菇产量统计
培养料二次发酵结束后出仓移入出菇房内,播种后进行常规的工厂化出菇管理,分别记录4 个配方的两潮草菇产量。
1.3 统计分析
数据分析采用SPSS 20 软件,计量资料用平均数±标准差(±SD)表示,多组间比较采用单因素方差分析(ANOVA),两两比较采用Duncan-Dunnett’s T3,P<0.05为差异有统计学意义。
2 结果与分析
2.1 pH测定
由图1 可知,培养料经过一、二次隧道发酵后pH 发生了变化。4 个配方均表现出相同的pH 变化趋势,即一次发酵后pH 下降明显,pH 全部下降至8.15~8.50。二次发酵后pH 变化较小,配方1 和配方2仍为下降趋势,但配方3和配方4却略有上升。
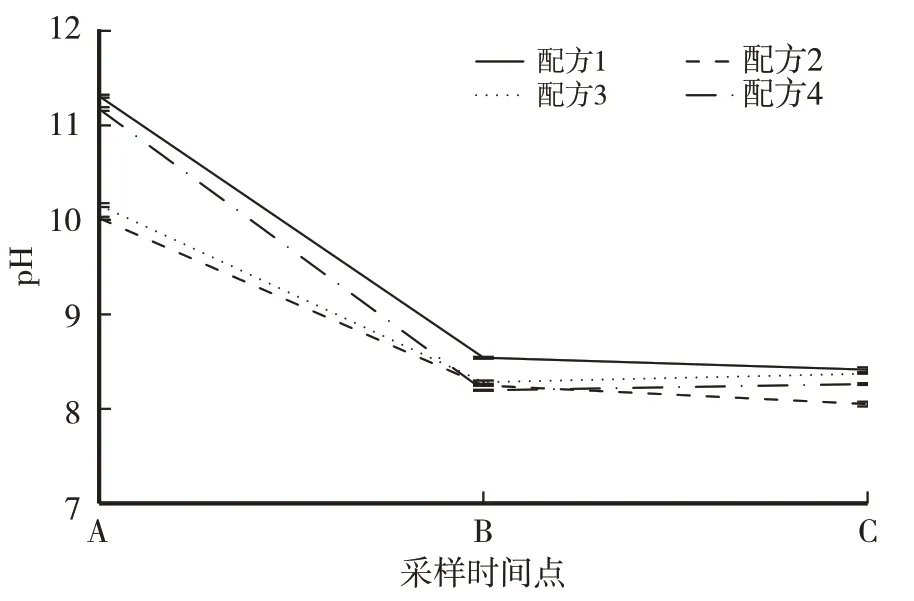
图1 供试配方料发酵过程pH变化
2.2 胞外酶活性分析
2.2.1 总蛋白酶活性的变化
培养料中含有丰富的蛋白质,在隧道发酵过程中总蛋白酶活性的变化呈现出不同的规律。由图2可以看出,配方1 培养料中总蛋白酶的活性经过两次隧道发酵变化很小,差异不显著;其他3个添加麦草的配方,培养料经过一次发酵后总蛋白酶活性均呈上升趋势,且差异显著,经过二次发酵后,配方2的酶活性依然上升,而配方3、配方4 酶活性却下降至2 IU/g;二次发酵结束后,配方2 的总蛋白酶活性显著高于其他3个配方。
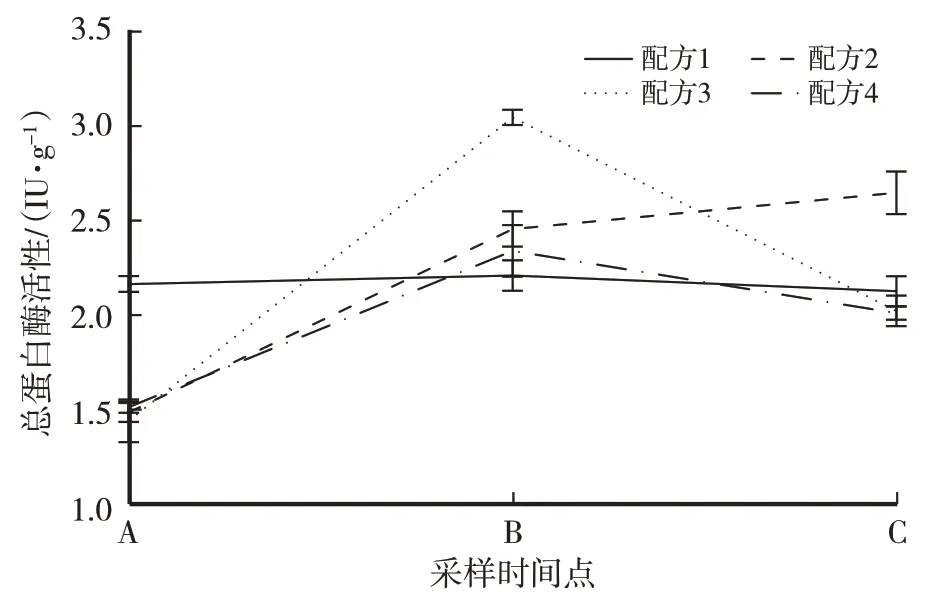
图2 供试配方培养料发酵过程总蛋白酶活性变化
2.2.2 木质素酶活性的变化
培养料在隧道发酵过程中木质素酶活性的变化趋势如图3 所示。麦草含量最高的配方4 在两次发酵中木质素酶活性都呈上升趋势,一次发酵前和二次发酵结束后的酶活性差异显著;其他3 个配方的培养料酶活性呈先上升后下降的趋势,其中无麦草的配方1培养料经过两次隧道发酵后酶活性从最高(0.58±0.02)U/g 降至最低(0.51±0.03)U/g,配方2和配方3培养料发酵过程中各时间点木质素酶活性的差异不显著。
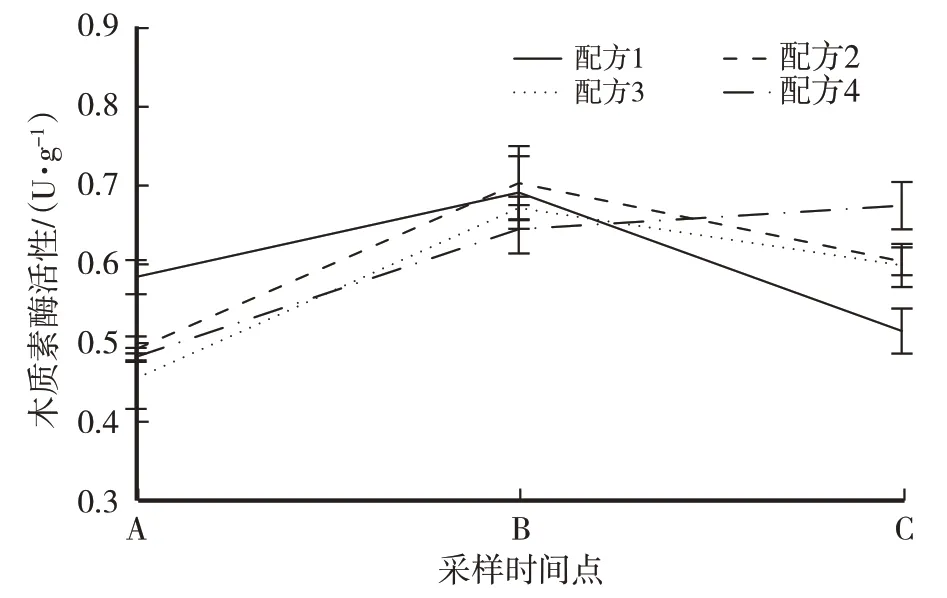
图3 供试配方培养料发酵过程木质素酶活性变化
2.2.3 半纤维素酶活性的变化
隧道发酵过程中降解培养料中半纤维素的主要酶是内切木聚糖酶,其变化趋势如图4 所示,4 个配方表现出3 种不同的变化趋势:配方1 培养料的酶活性变化为先降后升,且二次发酵结束后与一次发酵前的差异不显著;配方4 培养料经过每一次发酵后酶活性均明显地上升;配方2 和配方3 的培养料酶活性为先升后降,且每个取样点麦草含量较少的配方2培养料的酶活性都显著高于麦草含量较多的配方3。

图4 供试配方培养料发酵过程半纤维素酶活性变化
2.2.4 纤维素酶活性的变化
草菇培养料隧道发酵过程中纤维素酶活性变化如图5 所示,4 个配方的酶活性变化趋势一致,均在二次隧道发酵前出现了酶活性最高峰,其中配方3 的培养料酶活性最高,为(4 414.47±184.80)U/g。在二次隧道发酵结束后,除配方4 外,其他3 个配方的培养料酶活性都显著高于一次隧道发酵前。

图5 供试配方培养料发酵过程纤维素酶活性变化
2.3 供试配方培养料栽培草菇产量
4 个配方培养料栽培草菇采收两潮后记录草菇总产量。由表1可知,配方1和配方2的总产量显著高于配方3、配方4,配方1的出菇表现为第1潮菇产量最高,第2 潮菇产量显著降低;配方2 的两潮菇产量相当。
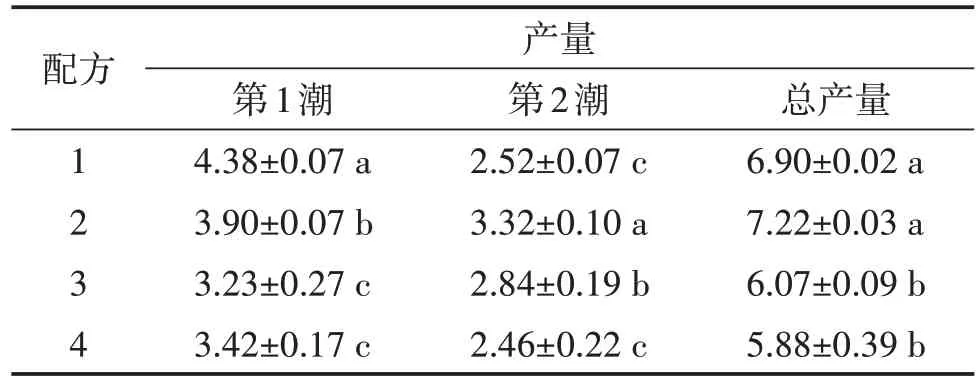
表1 供试配方培养料栽培草菇产量统计 单位:kg/m2
3 小结与讨论
试验表明,当培养料含有杏鲍菇菌渣90%和10%的麦草时,采收两潮菇的总产量最高,为(7.22±0.03)kg/m2,高于黄桂平采用纯杏鲍菇菌渣栽培草菇的产量[9],同样随着麦草比例的上升,产量逐渐降低。林金盛等[10]研究表明以杏鲍菇菌渣为培养料栽培草菇的产量显著高于以稻草为培养料的产量。这说明在草菇生长过程中菌渣提供了比麦草更多的营养物质,菌渣比例越低,产量越低。纯菌渣中加入少量麦草,在隧道发酵过程中料通气性更好,培养料发酵更加彻底。
草菇培养料隧道发酵时,除配方1外,其他配方一次隧道发酵培养料的半纤维素酶活性全部呈上升趋势。以废棉为培养料栽培草菇时一次发酵培养料的木质纤维素酶变化同样为上升趋势[5],但以秸秆为主要培养料栽培双孢蘑菇时一次隧道发酵的培养料酶活性变化差异较大[11],导致这些不同结果的原因可能与一次发酵的时间及条件相关。二次隧道发酵酶活性呈下降趋势的较多,但在二次隧道发酵结束后,所有酶活性均高于一次发酵前。这说明在隧道发酵时培养料中的微生物开始生长,并对培养料中的蛋白质、木质素、纤维素、半纤维素等分解利用,从而有利于草菇播种后菌丝的萌发生长[12]。杏鲍菇菌渣中存在木质素含量高的杂木屑,麦草的加入不仅能优化培养料结构,提高透气性,同时会额外增加纤维素和半纤维素含量,试验中麦草量与木质素酶、纤维素酶和半纤维素酶活性变化差异相关。
草菇培养料在两次隧道发酵过程中pH 表现出一致的变化规律,即一次发酵显著降低,二次发酵略微降低,与已报道的废棉栽培草菇[5]和菌渣栽培双孢蘑菇[13]规律一致。试验发酵前初始pH 用熟石灰调整后达pH 10~11,但二次发酵结束后的pH 为8.15~8.50,与赵书光等[14]报道的最适pH(7.5 左右)接近,这可能与本试验采用隧道发酵有关。
以菌渣为主要培养料的工厂化栽培草菇模式已是近年来草菇栽培的发展方向,隧道发酵则是工厂化栽培草菇中极为关键的环节之一。试验通过分析供试配方在发酵过程的酶活性变化及栽培草菇的产量,筛选出最适宜配方,为工厂化栽培草菇进一步降本增产、增效提供数据支持。
猜你喜欢
南方农机(2023年3期)2023-01-31 07:29:42
今日农业(2022年3期)2022-11-16 13:13:50
作物学报(2022年2期)2022-11-06 12:08:56
中华环境(2021年10期)2021-11-22 06:47:28
快乐语文(2020年14期)2020-07-04 00:02:00
饮食与健康·下旬刊(2019年3期)2019-03-08 14:33:32
现代食品(2018年10期)2018-02-14 17:27:48
上海农业学报(2017年3期)2017-04-10 12:39:08
临床医药文献杂志(电子版)(2017年98期)2017-03-06 19:43:21
化工管理(2017年29期)2017-03-03 09:19:22
