
个体亲近程度对藏酋猴攻击行为的影响
2022-07-25 09:54张通王希张启信李进华
兽类学报 2022年4期
张通 王希 张启信 李进华*
(1安徽大学资源与环境工程学院,合肥 230601)(2黄山生物多样性与短尾猴行为生态学国际联合研究中心,合肥 230601)(3合肥师范学院生命科学学院,合肥 230601)
群居动物在空间利用上存在差异(Guanet al.,2013),研究表明,不同动物个体间距离是友好行为和冲突行为的折中体现,表现为个体间距的差异(Wey and Blumstein,2010),而友好行为和冲突行为共同决定了社会结构及其表现形式(Schuelkeet al.,2020)。
在非人灵长类中,友好行为和冲突行为之间的关系是理解社会组织形式及社会维持机制的关键。友好行为主要包括近距和社会理毛行为(Arseneau-Robaret al.,2018),基于近距和理毛行为联合量化的社会联系强度可以表示个体间亲近程度(Silket al.,2010;Kalbitzet al.,2016),而量化的攻击行为则可以用于衡量冲突程度。动物个体间不同程度的亲近关系对攻击行为的影响不尽相同。雌性草原狒狒(Papio cynocephalus)社会联系越强的个体,理毛行为越多,攻击次数越少,攻击支持的次数越多(Seyfarth and Cheney,2012);在日本猕猴(Macaca fuscata)的研究中,社会联系越强的个体,攻击行为越少(Kawazoe,2021)。但在另一些研究中得到了相反的结论,在黑猩猩(Pan troglodytes)群体中,社会联系越强的个体间发生攻击行为的概率越高(Reddy and Mitani,2020);在恒河猴(Macaca mulatta)的研究中也发现社会联系强度越高的个体,攻击行为发生频次越多(Seyfarth and Cheney,2012)。此外,黑猩猩的近距行为可以单独表达亲近程度(Nishida,1968),近距虽然增加了个体间友好接触的机会,但也增大了个体间发生冲突的概率(Arseneau-Robaret al.,2018),有研究发现,雌性恒河猴近距时间越长,个体间攻击行为越多(Krawczaket al.,2002)。社会理毛行为和攻击行为之间也存在一定联系。黑猩猩在群体间发生攻击,会使群体内的理毛行为增加,增强了群体的凝聚力(Samuniet al.,2020)。在雄性藏酋猴(Macaca thibetana)、僧帽猴(Cebus apella)群体内,低顺位个体给高顺位个体理毛,会减少来自高顺位的攻击 (Xiaet al.,2012;Duboscqet al.,2014),但长尾叶猴(Semnopithecus entellus)的理毛和攻击行为呈正比(Gill,2017),卷尾猴(Cebus capucinus)发起理毛频次和受到攻击频次的社会网络也呈正相关关系(Crofootet al.,2011)。有研究表明,雌性卷尾猴中受到高顺位雌性攻击最多的是为其理毛最多的个体(Perry,1996)。由此可见,亲近关系对攻击行为会产生影响,但这种影响在物种间,甚至物种内都有差异,还有待深入研究。
动物会根据个体间熟悉程度、社会关系和性别来调节自己的行为,例如在绿猴(Chlorocebus sabaeus)的研究中发现,来自同一母系、亲近关系程度高和高顺位个体间的攻击行为中非接触型的攻击较多(Chalian and Meishvili,2006)。斑纹隐小鱂(Kryptolebias marmoratus)更喜欢与熟悉的个体或者近亲待在一起,并会相应地调节攻击性(Edenbrow and Croft,2012)。但拟寄生蜂(Parasitoid wasp)与竞争对手的熟悉程度不会影响攻击的程度 (Innocentet al.,2011)。绒毛猴 (Lagothrix lagotricha)雄性个体间比雌性个体间行为互动较少,且攻击大多都是象征性的,很少有接触型攻击(De Filippiset al.,2009;Shanee,2014)。由此可见,非人灵长类的攻击强度受亲近关系程度、顺位和性别的影响,但这些因素对于攻击强度的作用很少有研究报道,此外,近距、理毛行为和攻击频次的复杂关系尚未得到清晰的回答。
藏酋猴营群居生活,其社会群体由多个雄性、多个雌性以及其后代组成,雄性之间没有亲缘关系,雌性构成为母系群体以及少数迁入个体,社会关系复杂多样(Liet al.,2020)。雄性和雌性个体间遵循着严格的线性等级制度,攻击行为频发且攻击类型多样(Liet al.,2020)。这些特征十分有利于研究非人灵长类中不同社会关系个体间的攻击行为差异以及影响因素。
本文以安徽黄山的藏酋猴为研究对象,通过观察、收集并分析攻击行为、近距和理毛行为数据,探讨不同社会关系个体间的攻击行为差异以及影响因素,探究亲近关系、性别和顺位差等因素对藏酋猴个体间攻击强度的影响程度,近距、理毛行为和攻击频次之间的关系。提出以下预测:(1)个体间亲近程度越高,攻击频次越高,但攻击强度较低;(2)雄性间攻击频次低于雌性间,但攻击强度较高;(3)近距矩阵和理毛矩阵、攻击总次数、轻度攻击和重度攻击次数矩阵均呈正相关关系。本研究将为理解藏酋猴社会关系及结构形式提供一定的理论支持。
1 研究方法
1.1 研究地点与研究对象
研究地点位于中国安徽省黄山野生猴谷(北纬30°29′,东经118°11′),海拔600 ~ 1 200 m(Liet al.,2020)。研究群体为栖息于该研究地点的野生藏酋猴鱼鳞坑A1群(YA1)。对该群体自1986年持续跟踪观察,群体内全部个体均可以根据其自然特征准确识别,并且可以确定所有个体的母系亲缘关系。研究期间(2020年9月—2021年5月)猴群个体总数为58只,根据Li等(2020)进行年龄段的划分,其中成年个体24只,亚成年个体6只,未成年个体28只。本研究选取24只成年个体作为研究对象(表1)。

表1 研究期间藏酋猴YA1群成年个体种群组成Table 1 Composition of adult individuals in YA1 group during study period
1.2 取样方法
研究期间,每天08:00—17:00采用目标动物取样法(Focal sampling method)对目标动物所有行为进行观察和收集,目标动物的顺序用随机抽签的方法确定,取样时间为15 min。采用行为取样法(Behavioral sampling)结合全事件记录法(All-occurrence sampling method)记录猴群攻击行为全过程数据(Altmann,1974)。当攻击行为发生时,记录攻击行为的类型、发起者、接受者、接受者的反应行为类型以及有无第三方加入。根据Berman等(2004)的定义,将瞪眼、击地、短冲、长冲、追赶、抓、咬7种攻击行为按照有无身体接触分为轻度攻击和重度攻击,追赶行为发生后一般都会演变成有身体接触的攻击(Luet al.,2008),故将追赶也定义为重度攻击(表2)。

表2 藏酋猴友好和攻击行为参数定义Table 2 Definitions of affiliative and aggressive behaviors in Tibetan macaques
1.3 等级顺位
将猴群的攻击行为数据分别构建雄性和雌性的攻击—屈服矩阵(Aggression-submission matrix),然后基于此矩阵计算David’s score(DS)值,通过DS值确定雄性和雌性的个体等级顺位(Gammellet al.,2003)。具体的计算公式为:

其中,Pij代表个体i战胜个体j的次数占个体i和个体j攻击屈服总次数的比例,,aij代表个体i战胜个体j的次数,nij代表个体i和个体j之间发生攻击屈服行为的总次数。∑Wj×Pji代表个体i的所有Pji加权和,Wj代表个体j的所有Pji之和;∑Pji代表个体i的所有Pji之和;∑Ij×Pji代表个体j的所有Pji加权和,Ij代表个体j的所有Pij之和。
1.4 个体间近距和理毛与成对关系指数
个体间的近距行为是非人灵长类群体反应亲近关系的重要指标之一(Arseneau-Robaret al.,2018),研究采用成对联系指数(Dyadic association index,DAI)来表示个体间亲近关系程度。计算公式:

其中,∑A表示个体A与群体内其他个体的总近距时间;∑B表示个体B与群体内其他个体的总近距时间;∑(A+B)表示个体A和个体B单独近距的总时间。DAI值的前25%(0.110~0.570)为亲近关系,中间50%(0.024~0.110)为一般关系,后25%(0.000~0.024)为疏离关系(Nishida,1968)。
相互理毛行为既可以增加个体间的亲近关系,也可以修复被攻击破坏的关系,对于非人灵长类群体的稳定性十分重要(Xiaet al.,2012)。基于个体间理毛时间计算理毛指数(GI)(Hashimotoet al.,1996),公式为:

其中,fA(B)表示当观察个体A时,个体B给予或接受个体A理毛的总时间;同理,fB(A)表示当观察个体B时,个体A给予或接受个体B理毛的总时间,F(A)和F(B)分别代表个体A和B被观察取样的总时间。同理,也依据此公式计算个体间近距指数。
1.5 数据分析与处理
使用克鲁斯卡尔-沃利斯检验(Kruskal-Wallis)分析不同关系类型中成对个体的总攻击次数均值之间的差异性,以及不同关系类型中轻度攻击百分比和重度攻击百分比差异。利用Spearman相关系数分析顺位差和攻击次数以及强度的关系、相同性别个体间近距指数和顺位差之间的相关性关系。利用二项式检验(Binomial test)分析轻度攻击频次和重度攻击频次占比的差异性。基于近距和理毛指数构建近距和理毛矩阵。基于总攻击、轻度攻击和重度攻击次数构建攻击次数矩阵,采用Mantel test检验分析近距矩阵和理毛矩阵以及攻击次数矩阵的相关性,理毛矩阵与攻击矩阵的相关性。
采用广义线性混合模型(Generalized linear mixed model,GLMM)分析可能影响攻击次数和强度表达的因素(Nakagawa and Schielzeth,2013)。攻击强度用平均攻击分数表征,平均攻击分数的计算过程:将轻度攻击、重度攻击分别赋值1和10,个体间发生的轻度和重度攻击次数乘以赋值数,得到的总数除以总攻击次数,然后将得数进行对数转换以更符合模型(Cowlet al.,2021)。将以下因素作为模型预测变量:个体关系、理毛指数、顺位差(1~23)、不同亲缘系数 (母子0.5、兄妹0.25、表亲0.125)(Bermanet al.,2008)、性别差异(雌性对、雄性对和异性对)。雌性和雄性的身份作为模型随机作用变量以消除伪重复。
模型的构建过程如下:首先,利用似然比(Likelihood ratio test)对全模型和空模型(只包含随机项与截距项)之间的差异进行显著性检验(Forstmeier and Schielzeth,2011)。然后利用R包(car)对所有预测变量的潜在共线性进行检验(Healy,2005)。如果VIF值为0~10,说明变量间不存在共线性(Stapleton,2016),两个模型的VIF值均小于2。模型采用R Core Team 2020,version 4.0.3中的lme4 package运行,另外采用R语言中的ade4 package进行Mantel test的实现(Zhanget al.,2011)。
其他数据均利用SPSS 22.0进行分析,Originpro(9.6.5.169)进行制图。统计结果均为双尾检验,显著性水平设置为α=0.05,数据以平均值±标准差(mean±SD)表示。
2 结果
2.1 藏酋猴攻击行为
研究期间,共记录藏酋猴攻击行为380例,其中交配期15例,非交配期365例。在非交配期,轻度攻击229例 (62.74%),重度攻击136例(37.26%)。其中,轻度攻击频次占比与重度攻击频次占比差异性显著(n=365,P<0.001),攻击类型发生次数分别为:瞪眼114例(31.23%)、击地11例 (3.01%)、短冲70例 (19.17%)、长冲34例(9.32%)、追赶49例 (13.42%)、抓49例 (13.42%)和咬38例 (10.41%)。
不同类型攻击次数发生在亲密关系间171次(n=68)、一般关系间155次(n=139)和疏离关系间39次(n=72),3种亲近关系类型中成对个体的攻击总次数差异显著 (χ2=7.829,df=2,P=0.020)。轻度攻击频次百分比均高于重度攻击,但3种关系类型中成对个体的轻度攻击百分比(χ2=0.412,df=2,P=0.814)和重度攻击百分比 (χ2=0.761,df=2,P=0.683)差异均不显著(图1)。
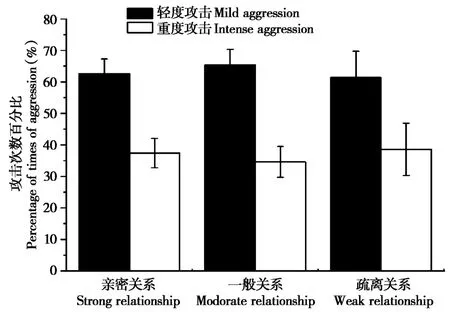
图1 藏酋猴不同亲近关系个体间轻度及重度攻击次数百分比Fig.1 Percentage of mild and intense aggression among individuals with strong,moderate and weak relationships of Tibetan macaques
2.2 藏酋猴近距、理毛和攻击矩阵的关系
近距矩阵与攻击总次数、轻度攻击和重度攻击次数矩阵存在显著正相关关系(图2)。近距矩阵和理毛矩阵显著正相关(r=0.39,P=0.02),理毛矩阵与轻度攻击矩阵和重度攻击矩阵无相关性,理毛矩阵和总攻击次数矩阵呈现正相关的趋势,但显著性未达到统计学水平(r=0.31,P=0.06)。此外,雄性间和雌性间近距指数和顺位差显著负相关(r=-0.181,n=142,P=0.032)。

图2 藏酋猴近距、理毛和攻击矩阵的相关性.A:近距、理毛和总攻击次数矩阵的相关性;B:近距、理毛和轻度攻击次数矩阵的相关性;C:近距、理毛和重度攻击次数矩阵的相关性Fig.2 Correlation diagram of the proximity,grooming and aggression matrix of Tibetan macaques.A:Correlation of proximity matrix,grooming and the total number of times aggressive interaction;B:Correlation of proximity matrix,grooming and the number of times mild aggression;C:Correlation of proximity matrix,grooming and the number of times intense aggression
2.3 藏酋猴攻击频次及强度的影响因素
GLMM分析结果表明藏酋猴个体间的亲近程度对攻击行为的总次数具有显著影响,并且两者之间是正相关关系。在不同的性别对中,雌性对间的总攻击次数最少。而顺位差、亲缘系数和理毛指数对藏酋猴间的攻击行为总次数没有影响(表3)。本研究构建的模型与零模型之间存在显著差异 (χ2=422.40,df=8,P< 0.001)。
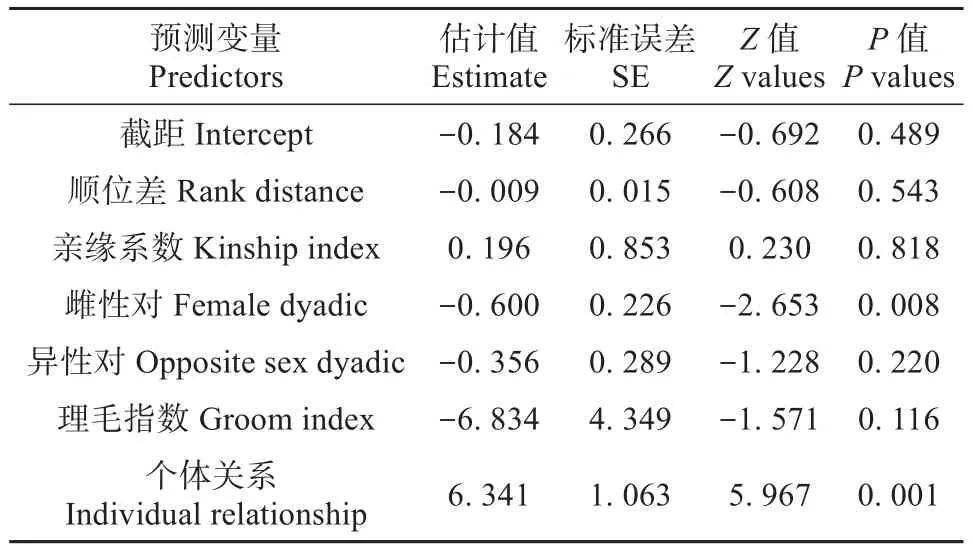
表3 广义线性混合模型测试影响藏酋猴攻击频次因素的结果Table 3 Results of the GLMM to test factor affect the number of times aggression of Tibetan macaques
雌性对(n=106)、雄性对(n=36)和异性对(n=136)间的总攻击次数分别为1.448±0.020、1.861±0.052和1.037±0.013,不同性别对的总攻击次数间存在显著差异(χ2=422.40,df=8,P<0.001)。
平均攻击强度影响因素的GLMM模型分析结果显示,性别和个体间亲近程度对藏酋猴的平均攻击强度产生显著影响,雌性对和异性对与平均攻击强度呈负相关关系,说明雌性对和异性对间比雄性对间的平均攻击强度小。个体亲近程度和平均攻击强度呈正相关关系,表明藏酋猴间关系程度越紧密,平均攻击强度越大。顺位差、亲缘系数和理毛指数对藏酋猴个体间平均攻击强度没有产生显著影响(表4)。此模型与零模型间具有显著的差异性 (χ2=182.32,df=9,P< 0.001)。
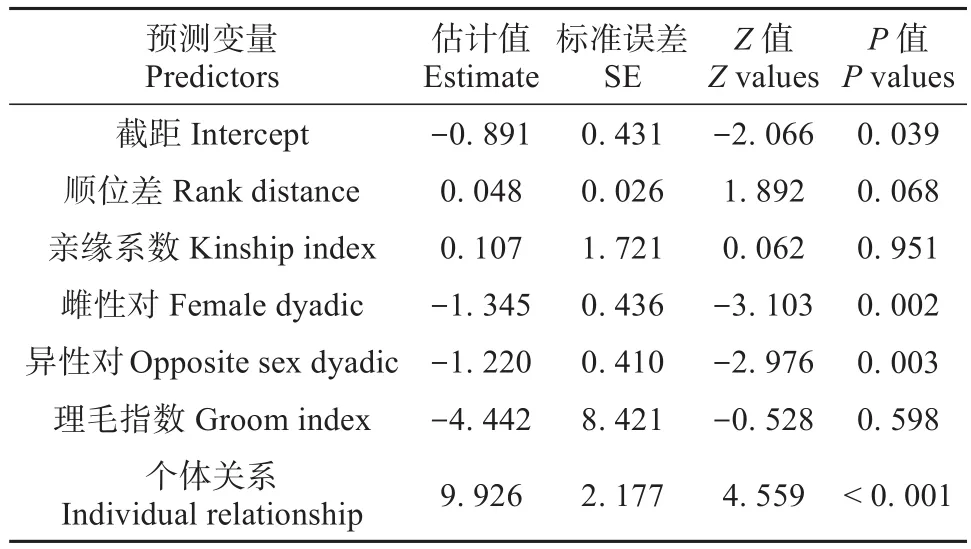
表4 广义线性混合模型测试影响藏酋猴平均攻击强度因素的结果Table 4 Results of the GLMM to test factor affect mean aggression degree of Tibetan macaques
雌性对、雄性对和异性对间的平均攻击强度分别为0.449±0.001、0.507±0.004和0.426±0.001,三者间具有显著差异性(χ2=8.671,df=2,P=0.013)。雄性对间的平均攻击强度显著高于雌性对(P=0.041)和异性对(P=0.003),雌性对和异性对间的平均攻击强度差异不显著(P=0.211)。亲密、一般和疏离关系个体间的平均攻击强度分别为0.496±0.001、0.441±0.001和0.399±0.002,三者间差异性显著(χ2=13.64,df=2,P=0.001),亲密关系个体间的平均攻击强度显著高于一般(P=0.008)和疏离关系个体间(P<0.001),一般关系个体间的平均攻击强度显著高于疏离关系个体(P=0.044)。
雄性对和雌性对间的总攻击次数和顺位差显著正相关(r=-0.012,n=141,P=0.012),平均攻击强度和顺位差无相关性(r=0.026,n=84,P=0.818)。
3 讨论
本研究分析了黄山藏酋猴近距、理毛和攻击行为矩阵之间的相关性,探讨了社会因素和生物因素对攻击行为的影响。发现近距指数和理毛指数呈正相关关系,且与总攻击、轻度攻击和重度攻击次数矩阵均显著正相关;当排除其他因素影响时,个体间顺位差和总攻击次数也呈正相关关系。个体间亲近关系差异和攻击对的性别不同会对攻击行为次数和强度产生影响。雌性对间的攻击次数和攻击强度均小于雄性对和异性对。但个体间亲近程度越高攻击次数和攻击强度均增加,这与假设预期相反。这些结果提示,近距行为可以维持个体和群体间其他成员的关系,但也会增加被高顺位攻击的风险,个体在进行社会交往时要权衡这种短期风险和长期利益。
3.1 近距和攻击行为的关系
本研究发现近距会增加藏酋猴理毛行为,但也引发藏酋猴个体间更多的攻击行为。这与Kummer(1978)提出近距会增加个体间社会行为的理论一致,与绒毛猴(Lagothrix flavicauda)研究中得出的结论类似(Shanee,2014)。近距是群居性动物社会交往的前提,个体之间理毛结束,一般不会立即离开对方,而是保持近距状态,这时候就会有可能因为婴猴、空间、惊扰、第三方行为或压力而对顺位低的一方发起攻击。若个体间发生攻击行为后,永久远离对方,会损害双方的利益(Wellset al.,1991)。根据胁迫理论(Extortion hypothesis),高顺位个体利用攻击的方式迫使从属个体为其理毛或其他行为,并监控从属个体的行为,它们之间会频繁地发生攻击行为,并且理毛行为也比其他个体间多(Rowellet al.,1991)。而低顺位个体主动与高顺位个体交往,维持个体间的亲近关系,可以获得更多的食物容忍和攻击支持。
然而,低顺位个体与高顺位个体近距会增加自身被攻击的风险(Schino and Alessandrini,2015)。如果低顺位个体与高顺位个体近距没有受到攻击,则个体利益增加,在高顺位个体附近可以被保护不受第三方的攻击。但近距与攻击的关系也可能和群体统治风格有关,即在统治风格宽松的群体中,个体间容忍度较高,近距并不会增加攻击频次(Noe,2006)。个体做出这种衡量短期和长期风险及利益的复杂决策,需要从很多方面说明,本研究仅涉及近距的风险方面。
3.2 攻击行为的社会生物学因素
黄山藏酋猴个体之间的关系越紧密,发生攻击的次数越多,并且平均攻击强度也最大。对黑冠猕猴(Macaca nigra)的社会关系研究发现社会联系强度越强的个体会更频繁地攻击对方,但大多数的攻击是轻度攻击(Duboscqet al.,2017),与本研究结果相异,可能本研究的分析单位是成对个体,个体间攻击次数样本量不够大,轻度攻击和重度攻击的差异性不显著。黄山藏酋猴在群体水平上,轻度攻击的占比较大,但与重度攻击的差异没有达到显著水平。藏酋猴关系更紧密的个体间之所以能够承受比其他个体间更多的攻击,是因为它们之间关系更稳定,冲突可调节其社会关系,但并不会破坏其紧密关系(Seyfarth and Cheney,2012);此外,虽然它们之间的攻击次数最多,但是友好交往相较于其他个体也最频繁,当一方与其他个体发生攻击时,另外一个个体会提供攻击支持。在群体水平上,轻度攻击的占比较大,其作用有调节社会关系、维持社会稳定和避免受伤。群体间的冲突更多是轻度攻击,有利于物种的自我保护,由此可见,轻度攻击具有更重要的生物学意义。
另外,藏酋猴雌性间的攻击次数最少,平均攻击强度也最小。在长尾叶猴(Semnopithecus entellus)雄性间冲突的受伤率是雌性间的3倍(Federet al.,2019)。激烈的冲突需要强大的战斗力,消耗更多的能量,受伤的概率也更大,这些情况在雄性中更常见。顺位差较小或亲缘关系较近的雌性间更可能形成稳定而持久的紧密社会关系,社会互动频繁,攻击行为较少且强度低;而雄性之间接触较少(Shanee,2014),且更多的是为了形成联盟,雄性间的竞争也较大(Wanget al.,2009),重度攻击较多。雄性和雌性之间的关系,主要是为了交配。在黑猩猩的研究中发现,社会关系紧密的异性对间性胁迫行为发生更频繁,这有利于提高繁殖的成功率(Reddy and Mitani,2020)。本研究期间观察到,最高顺位雄性藏酋猴会通过连续几天追赶并咬拒绝与其交配的雌性。
综上所述,本研究对黄山藏酋猴近距、理毛和攻击行为之间的关系以及攻击行为的影响因素进行了探讨,认为近距增大了个体间理毛和攻击行为发生的可能性,选择个体保持近距是衡量风险与获利的决策。社会关系紧密的个体间攻击次数和强度均最大,说明藏酋猴个体不会根据与另一个个体之间的关系调整攻击强度。雌性间攻击次数和强度最低,表明雌性之间的关系最稳定。本研究为了解非人灵长类的社会关系、认知水平以及社会结构形式提供了更多的基础资料,但对于群体水平的空间结构、关系质量和攻击行为的其他影响因素的探讨还需要继续深入全面的研究。
致谢:感谢黄山猴谷管理员谢升飞、汪有国对野外采样工作给予的帮助,感谢程海滨家庭在野外生活中的帮助。感谢安徽大学灵长类创新团队全体人员在科研和生活上给予的帮助。
猜你喜欢
中国当代医药(2022年25期)2022-10-20
新传奇(2022年26期)2022-07-22
中国典型病例大全(2022年9期)2022-04-19
小学阅读指南·低年级版(2020年9期)2020-10-12
中国房地产·综合版(2019年9期)2019-10-18
家庭医药(2019年9期)2019-09-23
文萃报·周五版(2019年18期)2019-09-10
小天使·六年级语数英综合(2019年2期)2019-01-12
儿童故事画报·自然探秘(2017年7期)2018-03-14
Coco薇(2015年11期)2015-11-09
