
亚硝态氮对虾肝肠胞虫感染脊尾白虾的影响
2022-07-20 01:59胡吉卉段健诚陈姝含张庆起赖晓芳
水产科学 2022年4期
胡吉卉,段健诚,高 阳,陈姝含,张庆起,牟 华,3,4,赖晓芳,3,4,高 焕,3,4
(1.江苏海洋大学 海洋科学与水产学院,江苏省海洋生物技术重点实验室,江苏 连云港 222005;2.连云港赣榆佳信水产开发有限公司,江苏 连云港 222100; 3.江苏省海洋生物产业技术协同创新中心,江苏 连云港 222005; 4.江苏省农业种质资源保护与利用平台,江苏 南京 210014)
虾肝肠胞虫(Enterocytozoonhepatopenaei)是一种具有高度传染性的专性细胞内寄生虫,感染后造成虾类生长缓慢或停滞,是影响全球虾类养殖生产的重要病原之一[1-2]。目前普遍认为,极管是虾肝肠胞虫感染宿主细胞的主要细胞器,当孢子进入宿主体内时,由于外界环境的刺激,孢子内部压力急剧增大,极管外翻并弹出,刺破宿主细胞膜,然后将孢原质注射进宿主细胞内部。孢原质在新的细胞中增殖循环,最终产生新的孢子,从而实现孢子的繁殖和遗传物质的复制[3-5]。虾肝肠胞虫的主要靶器官为肝胰腺,而肝胰腺也是慢性亚硝态氮中毒的虾类中病变最严重的器官[6],同时虾肝肠胞虫的传播也受水质条件的影响。虾肝肠胞虫的传播有两种途径,一种是母体宿主传递到子代宿主的垂直传播,另一种是通过摄食携带虾肝肠胞虫的肝胰腺组织或养殖水体的水平传播[7]。
近年来,虾肝肠胞虫的感染率逐年上升,给养殖户造成了极大的经济损失,严重制约了我国虾类养殖业的发展。然而,目前针对虾肝肠胞虫的研究仍停留在定性或定量检测阶段,尚未研发出有效的治疗药物,通常是通过调节水质、降低养殖密度、增强池底排污、控制饲料投喂等方式加以控制[7]。
亚硝态氮作为氮代谢的中间产物,是水产养殖环境中主要的污染物之一[6,8],尤其是在高密度养殖条件下,亚硝态氮是诱发虾类暴发性疾病的重要因素之一[9]。目前,亚硝态氮在虾类中的研究主要集中在其急性或慢性毒性方面[10-13],虾肝肠胞虫在虾类中的研究主要集中在其对虾类的生长状况的影响方面[5,14-15]。已有研究表明,亚硝态氮胁迫显著影响甲壳动物的生理代谢、抗氧化能力和免疫功能,增加甲壳动物对病原体的易感性,抑制其生长和变态发育[6,16-19]。徐胜威等[20-21]研究表明,一定剂量的亚硝态氮和氨氮在抑制虾肝肠胞虫传播的同时能促进凡纳滨对虾(Litopenaeusvannamei)的生长。而亚硝态氮、脊尾白虾(Exopalaemoncarinicauda)和虾肝肠胞虫三者之间的内在关系鲜见报道。因此,笔者以脊尾白虾为试验对象,探讨不同质量浓度的亚硝态氮对脊尾白虾感染虾肝肠胞虫过程中的生长情况及体内虾肝肠胞虫携带量的影响,以期为脊尾白虾养殖过程中控制虾肝肠胞虫的养殖水质管理提供参考依据。
1 材料与方法
1.1 试验材料
试验用健康脊尾白虾和病虾均取自江苏省南通市如东县养殖池溏,感染用病虾用常规PCR检测,确认虾肝肠胞虫阳性后,-20 ℃冻存。在实验室进行适应性暂养1周后,选取300尾体长(4.1±0.1) cm、体质量(1.02±0.05) g、健康活泼的脊尾白虾用于试验。
试验容器为55.2 cm×41.5 cm×30.5 cm的蓝色塑料箱。以配制的人工海水为试验用水,盐度24±1,水温(25±1) ℃,pH 8.0±0.2,溶解氧(6.5±0.5) mg/L。亚硝态氮母液由分析纯亚硝酸钠[生工生物工程(上海)股份有限公司]配制而成。
1.2 试验方法
1.2.1 试验设置及日常管理
根据预试验结果,设置对照组(0.008 mg/L)和4个亚硝态氮试验组,即3.000、6.000、9.000 mg/L和12.000 mg/L,每组2个平行,每个平行放置30尾脊尾白虾,试验周期为21 d。试验期间每日按体质量的3%投喂虾肝肠胞虫病虾2次(8:00、20:00),投喂2 h后吸污,每24 h换10%的等质量浓度亚硝态氮的海水。每日监测养殖水体亚硝态氮质量浓度,并用亚硝酸钠母液稳定试验所需的亚硝态氮质量浓度。
1.2.2 生长指标测定
试验期间每隔1周从每个平行组中随机取3尾脊尾白虾进行体长和体质量的测定,并取肝胰腺组织于-20 ℃保存备用;记录每组的脊尾白虾死亡数量,试验结束后计算每组样品的存活率。
1.2.3 样品肝胰腺总DNA的提取
取感染虾肝肠胞虫的脊尾白虾肝胰腺组织约25 mg,使用Ezup柱式动物基因组DNA抽提试剂盒[生工生物工程(上海)股份有限公司]提取脊尾白虾总基因组DNA,用超微量核酸蛋白测定仪(Q5000,Quawell)测定样品DNA浓度后于-20 ℃保存。
1.2.4 常规PCR扩增
1.3.4 福山区政府政策优势。福山区政府确立了以特色樱桃产业带动经济发展、以优秀电商模式推动特色樱桃产业的双驱动模式,不断提升樱桃特色产业的水平,使大樱桃的电商模式驶入“快车道”。自2006年以来,在面对其他大樱桃地区的快速发展时,福山政府推动了樱桃品牌的创立,大樱桃产业发展需要品牌意识,以实现利益与品牌价值的捆绑提升,在提高水果质量的同时,也提升了品牌认证的力度,不断地巩固福山区大樱桃的优势地位[1]。
以总基因组DNA为模板,参考刘珍等[1]设计的定量特异引物(EHP-F:5′-GTA GGG GAA CGG ATA GGG-3′,EHP-R:5′-CAA GCA TTG TCG GCA TAG-3′)进行PCR扩增。采用2×Taq Plus Master Mix Ⅱ(Dye Plus)(南京诺唯赞)的反应体系。反应程序为:94 ℃预变性3 min;94 ℃ 20 s,60 ℃ 20 s,72 ℃ 30 s,共35个循环;72 ℃延伸5 min。
1.2.5 标准曲线的制备
采用1%琼脂糖凝胶电泳技术对PCR产物进行检测,观察其结果并拍照,使用SanPrep柱式胶回收试剂盒[生工生物工程(上海)股份有限公司]切胶回收。将得到的目的片段用T3载体在大肠杆菌(Escherichiacoli) DH5α中克隆,克隆的片段送生工生物工程(上海)股份有限公司测序验证。目的产物的克隆经扩大培养后,用质粒提取试剂盒[生工生物工程(上海)股份有限公司]提取质粒DNA。采用超微量核酸蛋白测定仪(Q5000,Quawell)测定所提质粒的浓度,换算成载量为1.15×1010拷贝/μL。将质粒标准品进行10倍梯度稀释,共得到1.15×106~1.15×1010拷贝/μL的5个梯度,每个梯度3个平行。以循环阈值为纵坐标,质粒标准品拷贝数为横坐标,绘制标准曲线。
1.2.6 实时荧光定量PCR的测定
参照Cham Q Universal SYBR qPCR Master Mix试剂盒说明书准备反应体系,反应体系为10 μL,包括含有SYBR荧光染料的预混液(SYBR qPCR Master Mix)5.2 μL,0.4 μL正向引物(EHP-F,10 μmol/L),0.4 μL反向引物(EHP-R,10 μmol/L),2 μL模板,2 μL双蒸水。扩增反应在实时荧光定量PCR仪(StepOne-PLUS)中进行。反应程序为:95 ℃预变性30 s;95 ℃ 10 s,60 ℃ 30 s,40个循环。
将不同样品中虾肝肠胞虫基因的表达量所对应的循环阈值与标准曲线相对照,即可计算出样品的载量。
2 结 果
2.1 虾肝肠胞虫定量PCR标准曲线
质粒标准模板在1.15×106~1.15×1010拷贝/μL内有明显扩增,每个质量浓度的3个平行循环阈值的差异符合qPCR要求(图1)。由图1可见,r2为0.99656,表明标准曲线线性关系较好,可用于虾肝肠胞虫载量检测。
2.2 亚硝态氮对感染虾肝肠胞虫的脊尾白虾的存活率的影响

图1 虾肝肠胞虫实时荧光定量PCR标准曲线Fig.1 Standard curve of E. hepatopenaei (EHP) real-time quantitative PCR
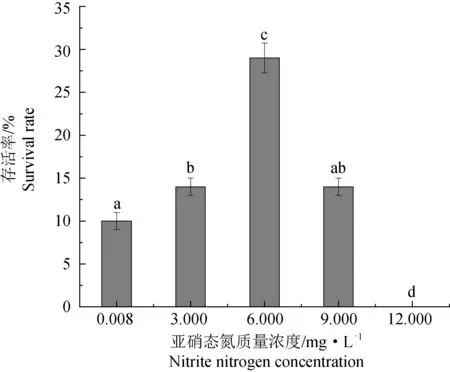
图2 亚硝态氮对感染虾肝肠胞虫的脊尾白虾存活率的影响Fig.2 The effect of nitrite nitrogen on the survival rate of ridgetail white prawn infected with EHP
2.3 亚硝态氮对感染虾肝肠胞虫的脊尾白虾的体长的影响
亚硝态氮胁迫下,各质量浓度组的体长变化基本一致,均呈现单调递增的趋势(图3)。随着亚硝态氮质量浓度的升高,各质量浓度组间呈现先升后降的趋势,其中亚硝态氮6.000 mg/L组的脊尾白虾体长增长最为明显,在胁迫21 d后体长达到最大值,平均为4.9 cm,平均增量0.8 cm。在处理7 d后,9.000 mg/L和12.000 mg/L的体长增长较低质量浓度组快,但随着处理时间的延长,高质量浓度组的体长增长比低质量浓度组慢。结果表明,一定质量浓度范围内的亚硝态氮对感染虾肝肠胞虫的脊尾白虾的体长增长有一定的促进作用,而质量浓度过高则会抑制其体长增长。

图3 亚硝态氮对感染虾肝肠胞虫的脊尾白虾体长的影响Fig.3 The effect of nitrite nitrogen on the body length of ridgetail white prawn infected with EHP标有不同小写字母者表示组间有显著性差异(P<0.05),标有相同小写字母者表示组间无显著性差异(P>0.05),下同.Means with different letters are significant differences (P<0.05), and means with the same letter are not significant differences (P>0.05), et sequentia.
2.4 亚硝态氮对感染虾肝肠胞虫的脊尾白虾的体质量的影响
亚硝态氮胁迫下,各质量浓度组的体质量随时间延长均呈增加的趋势。随着亚硝态氮质量浓度的升高,各质量浓度组间呈现先升后降的趋势,其中亚硝态氮6.000 mg/L组的脊尾白虾体质量增长最为明显,在胁迫21 d后,虾的体质量达到最大值,质量增加率为35.33%(表1)。处理7 d后,9.000 mg/L和12.000 mg/L的体质量增长比低质量浓度组快,但随着处理时间的延长,高质量浓度组的增长比低质量浓度组慢。结果表明,亚硝态氮在一定质量浓度范围内对感染虾肝肠胞虫的脊尾白虾的体质量增长有促进作用,质量浓度过高则会抑制其体质量增长。

表1 亚硝态氮对感染虾肝肠胞虫的脊尾白虾体质量的影响
2.5 亚硝态氮对感染虾肝肠胞虫的脊尾白虾的虾肝肠胞虫载量的影响
肝胰腺组织中虾肝肠胞虫的载量见图4。各质量浓度组的虾肝肠胞虫载量随时间延长的变化趋势基本一致,呈现单调递增的趋势。处理21 d后,各亚硝态氮质量浓度组间虾肝肠胞虫载量随亚硝态氮质量浓度的升高呈现先降后升的趋势,其中亚硝态氮6.000 mg/L组的虾肝肠胞虫载量达到最低值,为4.4×103拷贝/mg。处理7 d后,9.000 mg/L和12.000 mg/L的载量增长比低质量浓度组慢,但随着处理时间的延长,高质量浓度组的载量增长比低质量浓度组快。结果表明,一定质量浓度范围内的亚硝态氮会抑制虾肝肠胞虫的增殖,从而促进感染虾肝肠胞虫的脊尾白虾的生长。

图4 亚硝态氮对脊尾白虾感染虾肝肠胞虫载量的影响Fig.4 The effect of nitrite nitrogen on the EHP carrying amount of ridgetail white prawn
3 讨 论
3.1 亚硝态氮对感染虾肝肠胞虫的脊尾白虾的存活及生长情况的影响
亚硝态氮是虾类集约化养殖过程中最常见的毒性污染物之一,主要来源于虾类代谢废物、死亡个体及残余饵料的分解作用[8]。通常认为亚硝态氮在一定质量浓度范围内对水产动物无毒害作用[22],随着养殖周期的延长,亚硝态氮的积累量增多,通常在养殖水体中的亚硝态氮质量浓度超过0.1 mg/L时,会引起虾类免疫功能下降及组织器官损伤,诱发多种疾病[11,23]。研究表明:亚硝态氮胁迫后红螯螯虾(Cheraxquadricarinatus)的免疫酶活性显著降低,肝胰腺和鳃形态学结构受损[24];亚硝态氮与氨氮胁迫可能会抑制日本沼虾(Macrobrachiumnipponense)肝胰腺血红蛋白的合成及血糖调节[25];高浓度亚硝态氮会破坏日本沼虾的肝胰腺结构,影响其体内酶的催化作用和细胞膜的稳定性[26];在高密度养殖后期,池塘中亚硝酸氮的质量浓度高达20 mg/L[24],造成虾类的大量死亡[25]。一般认为,Cl-的浓度越高,竞争吸收进入虾体内的NO2-就越少,因此海水虾类对亚硝态氮的耐受能力要普遍高于淡水虾类[6,27-28]。为使胁迫效应更加显著,笔者选定了较高的亚硝态氮质量浓度,试验过程中发现,随着亚硝态氮质量浓度的升高,感染虾肝肠胞虫的脊尾白虾的存活率呈现先升后降的趋势,其中6.000 mg/L组的存活率最高,这与徐胜威等[21]的研究结果携带虾肝肠胞虫的凡纳滨对虾随亚硝态氮质量浓度升高存活率呈先升后降的趋势一致。本试验中,当亚硝态氮质量浓度达到12.000 mg/L时,脊尾白虾的死亡率急剧增加,分析可能是由于较高质量浓度的亚硝态氮胁迫,毒性作用增强,导致脊尾白虾的耐受性和免疫力降低,从而引起其大量死亡。
3.2 亚硝态氮对感染虾肝肠胞虫的脊尾白虾的虾肝肠胞虫载量的影响
近年来,虾肝肠胞虫已成为制约虾类养殖业健康发展的主要病害之一,造成了巨大的经济损失[29]。刘珍[30]发现,人工投喂冰冻和鲜活病虾的肝胰腺组织均能使健康的凡纳滨对虾感染虾肝肠胞虫,说明虾肝肠胞虫具有抵御低温的能力,这为生产上虾肝肠胞虫的预防和治疗增加了困难。虾类的肝胰腺为虾肝肠胞虫提供了最适宜的寄生条件,因此虾肝肠胞虫主要感染虾类的肝胰腺[31],而虾肝肠胞虫本身没有合成腺苷三磷酸的能力,完全依赖于截取宿主的腺苷三磷酸来获取营养,从而影响肝胰腺和肠道的正常消化吸收功能,阻碍宿主的正常生长和肌肉蛋白的积累[5,32-33]。程东远[5]研究发现,宿主腺苷三磷酸的含量与虾肝肠胞虫的载量显著负相关,刘珍等[1]通过实时荧光定量PCR检测发现,当对虾肝胰腺中EHP SSU rDNA的相对拷贝数在103拷贝/ng数量级或虾肝肠胞虫载量指数在3以上时,虾肝肠胞虫已处于较高的风险水平,且虾类的体长、体质量等生长指标与虾肝肠胞虫的载量显著负相关。本试验中,亚硝态氮6.000 mg/L组的虾肝肠胞虫载量最低,为4.4×103拷贝/mg,推测是因为亚硝态氮通过呼吸作用进入虾体内,引起组织缺氧,摄食减少,损伤肝胰腺及肠道,导致机体合成腺苷三磷酸的能力下降,虾肝肠胞虫不能获取足够的能量,生长发育受到抑制,因此虾类携带虾肝肠胞虫的量减少[34]。本试验结果表明,亚硝态氮6.000 mg/L组抑制虾肝肠胞虫增殖的效果是最好的。
4 结 论
综上所述,在一定质量浓度内(<6.000 mg/L),亚硝态氮可抑制虾肝肠胞虫的生长繁殖,从而促进感染虾肝肠胞虫的脊尾白虾的生长,而较高质量浓度(6.000~12.000 mg/L)的亚硝态氮对感染虾肝肠胞虫的脊尾白虾的生长不利,高质量浓度(≥12.000 mg/L)的亚硝态氮会引起感染虾肝肠胞的脊尾白虾死亡。
猜你喜欢
教育周报·教研版(2021年47期)2021-12-19
健康之家(2021年6期)2021-09-08
作文周刊·小学二年级版(2021年20期)2021-08-10
人人健康(2020年4期)2020-05-25
恋爱婚姻家庭·养生版(2016年11期)2016-11-03
中学生数理化·高三版(2016年9期)2016-05-14
海外英语(2013年1期)2013-08-27
中国民族民间医药·下半月(2011年10期)2011-12-27
中学生数理化·高一版(2009年6期)2009-08-31
