
青藏高原东缘3种次生林优势种的种群结构与数量动态
2022-07-12 14:04:42曹秀文刘锦乾陈学龙白青蒙王若鉴
西北农林科技大学学报(自然科学版) 2022年7期
王 飞,曹秀文,刘锦乾,齐 瑞,赵 阳,张 涛,陈学龙,李 波,杨 静,白青蒙,王若鉴,要 静
(1 甘肃省白龙江林业科学研究所,甘肃 兰州 730070;2 甘肃白龙江森林生态系统国家定位观测研究站,甘肃 舟曲746300)
种群是物种进化的基本单位[1],是指同一物种个体在同一地区、同一时期的集合[2]。种群结构是种群最基本的特征,包括年龄结构、径级结构、高度结构等不同类型,与种群动态同为种群生态学研究的核心问题之一[3]。生命表是种群统计学的核心,其反映种群对当前环境的适应状态[4]。利用静态生命表和生存分析展现种群现状及其发展规律,并结合基于种群数量动态变化的时间序列分析,对种群内不同大小个体数量的分布状况和未来时空变化规律进行预测[5-6],有利于揭示种群生物学特征与生境之间的关系[7],对物种多样性保护和管理具有重要意义[8]。次生林是因人为破坏或异常自然干扰使原始林的林分结构、建群种组成或基本功能发生显著变化,随后经过天然更新或人工诱导天然更新恢复形成的林分[9]。甘肃南部林区主要以白龙江和洮河林区为主,20世纪六七十年代大规模采伐以后,形成了以辽东栎(QuercuswutaishanseaMary)、红桦(BetulaalbosinensisBurk)和糙皮桦(BetulautilisD.Don)等为主的次生林,在涵养水源、保持水土、调节气候以及碳循环中发挥着重要作用,也对农业生产起着屏障保护作用,其森林生态服务功能价值占甘肃省的20%以上[10]。相关学者对次生林种群结构研究较多,如张婕等[3]对灵空山辽东栎的研究表明,其生存曲线为Deevey-Ⅱ型,且表现出前期衰败、中期增长、后期稳定的特征;王进等[11]研究发现,鄂西南亮叶桦幼龄苗数量不足,种群生存能力较低,趋于衰退,亮叶桦作为群落演替先锋树种逐渐被优势种代替;郭垚鑫等[12]认为,小陇山林区红桦次生林占明显优势地位,并在未来仍具有生长优势,但从径级结构看,其幼苗匮乏,天然更新不良;田慧霞等[13]通过分析得出,山西太岳山白桦种群为衰退型。相较而言,近年来对甘肃南部白龙江、洮河林区的研究多针对人工林和亚高山灌丛[14-16],而对次生林群落结构和动态变化等的研究较少。
甘肃南部白龙江林区和洮河是甘肃重要的水源涵养区,也是长江、黄河上游重要的水源补给区和生态屏障,对生态安全起到重要作用。本研究以白龙江、洮河林区辽东栎、红桦和糙皮桦这3种次生林为对象,通过实地调查,利用种群数量变化动态指数和生存分析方法,分析3种次生林的种群结构和生存现状,揭示种群动态发展趋势,以期为甘肃南部群落资源和次生林的健康发展提供科学依据。
1 研究区概况与研究方法
1.1 研究区概况
研究区设在甘肃南部白龙江和洮河林区,位于东经102°46′~104°52′,北纬33°04′~35°09′,海拔2 000~3 500 m,地处青藏高原东缘与黄土高原、秦巴山地和四川盆地的交界处,其地理位置独特,也是长江和黄河上游交汇区,汇集了森林资源、珍稀野生动植物资源、高原湿地、高原草甸等多种自然资源[17],属于典型的高山地形,温度、降水具有明显的垂直分布和水平差异[18]。林区成土母质为坡积母质,土层厚度在1 m以内,表层石砾含量少;全年平均气温1.3 ℃,年降水量1 048 mm,多集中在5-10月,年平均空气相对湿度80%,无霜期80~103 d[15]。经过上世纪的采伐后,林区形成了以红桦、辽东栎和糙皮桦为主的天然次生林。
1.2 研究方法
1.2.1 样地设置 2019年7-9月,依据甘肃南部3种次生林的基本分布情况和实地踏查结果,采用典型抽样法,选择具有代表性的辽东栎、红桦和糙皮桦种群,各设置8块40 m×40 m的标准样地(样地信息见表1),对样地内胸径(DBH)≥3 cm的所有树木进行每木检尺,测量胸径、冠幅、树高、枝下高等指标。采用相邻格子法,以5 m×5 m为基本单元,对DBH<3 cm的树木逐一测量地径、株高、冠幅。样地采用GPS定位,记录经、纬度和海拔,同时记录调查时间、坡向、坡度、郁闭度等因子。

表1 甘南3种次生林优势种样地信息
1.2.2 龄级划分 种群的径级和龄级有所不同,但同一环境、同一种群的龄级和径级对环境的反应规律具有一致性[19]。由于在不破坏树木情况下很难获取种群具体的树龄,故多数学者采用径级结构代替龄级结构[20-21],本研究也用径级结构代替龄级结构分析种群数量动态。因3种次生林均属于同一地区且形成原因一致,故采用相同的径级划分方法,具体为:按胸径大小每4 cm为一级,DBH<3 cm为Ⅰ龄级,3 cm≤DBH<7 cm为Ⅱ龄级,7 cm≤DBH<11 cm为Ⅲ龄级,…,依此类推。据此将辽东栎划分为11个龄级,红桦划分为12个龄级,糙皮桦划分为9个龄级。按上述划分标准,统计3个种群每个龄级的个体总数,建立种群静态生命表。
由林木中各个径级株数所占比例能够推测出林分的动态变化和群落的发展趋势[22]。利用Me-yer[23]的负指数分布和q值理论可得出异龄林径级与株数的关系,其公式为:
N=ke-ad;q=keah。
式中:N为株数,d为胸径,k和a为负指数分布的结构常数,q为相邻径阶株数之比,h是径阶距。
1.2.3 种群静态生命表编制与生存分析 静态生命表利用同一时期种群所有个体的径级编制而成,不是对某一种群全部生活史的追踪[17],实际上反映了种群在某一特定时刻的数量存活情况,也反映了多个世代重叠的年龄动态历程中一个特定时间内种群数量动态的变化规律。对于自然种群,在调查中可能存在系统误差,死亡率可能会成为负值,为了避免这一情况[1],采用匀滑处理[24]后的数据制作静态生命表。
静态生命表主要包括以下参数:ax是种群在x龄级的实际存活数,Ax是ax匀滑处理后的结果,lx是种群在x龄级的标准化存活数,dx是种群从x到x+1龄级的标准化死亡数,qx是种群从x到x+1龄级的标准化死亡率,Lx是种群从x到x+1龄级的标准化存活个体数,Tx是种群从x龄级到超过x龄级的个体数,ex是进入x龄级个体的平均期望寿命,Kx为消失率(损失度),Sx为存活率,其计算公式如下[25-29]:
lx=(Ax/a1)×100;
dx=lx-lx+1;
qx=dx/lx;
Lx=(lx+lx+1)/2;
Tx=∑Lx;
ex=Tx/lx;
Kx=lnlx-lnlx+1;
Sx=lx+1/lx。
存活曲线借助存活个体数量来描述特定年龄的死亡率,反映种群在各个龄级的生存现状[30]。本研究以标准化存活数(lx)为纵坐标、以龄级为横坐标绘制存活曲线。存活曲线分3种类型,Deevery-Ⅰ型为凸线型,符合该线型曲线的种群前期死亡率低,到一定生理年龄时,种群出现大量死亡,死亡率增大;Deevery-Ⅱ型是直线型,种群在各个龄级死亡率基本相同;Deevery-Ⅲ是凹线型,种群前期死亡率高,高龄级死亡率相对较低[31]。 采用2种数学模型对存活曲线进行拟合检验,即用指数函数方程式lx=ae-bx描述Deevey-Ⅱ存活曲线,用幂函数指数方程式lx=ax-b描述Deevey-Ⅲ存活曲线,以检验各种群存活状况更符合哪种曲线。式中lx为标准化存活数,x为龄级,a和b为常数。
1.2.4 种群动态分析和时间序列预测 种群动态量化分析方法参考陈晓德[32]的种群与群落结构动态量化分析方法。忽略外部干扰时,用Vpi表示整个种群结构的数量变化动态指数,用Vn表示n到n+1龄级的种群数量变化,Sn和Sn+1分别表示第n和n+1龄级的种群个体数,k为种群龄级,P表示种群对外界干扰所承担的风险概率。当考虑未来的外部干扰时,Vpi还与种群龄级(k)以及各龄级内的个体数(Sn)相关,可进一步用修正后的V′pi表示。其计算公式分别如下:
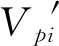
另外,采用时间序列分析中的一次移动平均法[32],对3种次生林种群的龄级结构进行预测,其公式为:
式中:Mt表示未来n年时t龄级的种群存活数;n表示需要预测的未来时间年限;t为龄级;Xk为k龄级内的个体数,k=t-n+1。本研究对未来经过2,4,6,8个龄级后各种群的数量进行了时间序列预测[33]。
2 结果与分析
2.1 3种次生林优势种种群的龄级结构
种群龄级结构是对种群现状的客观反映,也可以通过其预测未来种群的变化趋势。如图1所示,辽东栎、糙皮桦和红桦均以Ⅱ龄级个体最多,分别占该种群总数的43.62%,43.30%和30.07%;辽东栎和糙皮桦种群结构均呈单峰变化,Ⅰ龄级个体数小于Ⅱ龄级,其后随着龄级的增大种群个体数逐渐减少;红桦种群结构呈双峰变化,第1个峰值出现在Ⅱ龄级,第2个峰值出现在Ⅸ龄级。辽东栎Ⅰ龄级个体数占总数的26.42%,Ⅲ~Ⅺ龄级个体数之和只占总数的29.96%,Ⅱ龄级以后个体逐步减少且种群结构趋于稳定,故研究区辽东栎主要以Ⅰ和Ⅱ龄级个体为主。辽东栎Ⅰ龄级个体数量大于红桦和糙皮桦,3种次生林种群Ⅰ~Ⅳ龄级个体数之和分别占个体总数的91.23%,80.98%和91.44%,说明3种次生林种群主要以幼树为主。

图1 甘南3种次生林优势种种群的龄级结构
利用负指数分布和q值理论对3种次生林种群径级分布进行拟合后的参数见表2,其中a值表示林木株数相对于径级的递减速率,其值越大,说明林木株数随径级的增加迅速下降,相反说明随着径级的增加而平稳下降[34]。由表2可知,3种次生林种群a值均相对较小,说明种群株数随着径级的增大平稳下降;但R2值较低,表明拟合效果不佳。异龄混交林的径级结构q值以1.2~1.7为标准,当q值小于1.2,表明林木径级较小;q值大于1.7,则表明林木径级较大[32]。3种次生林的q值在1.012~1.024,均小于1.2,说明林木径级较小,龄级结构呈“钟型”。上述结果表明,虽然3种次生林群落为稳定型,但是各龄级株数分布不合理。
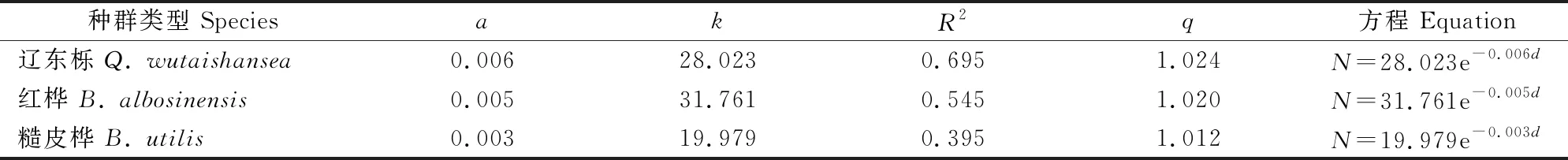
表2 甘南3种次生林优势种种群径级分布的拟合参数
2.2 3种次生林优势种种群的静态生命表和生存分析
静态生命表是种群动态分析的一种强有力的手段,反映种群在某一时间段内的生存现状。3种次生林优势种种群的静态生命表如表3所示。
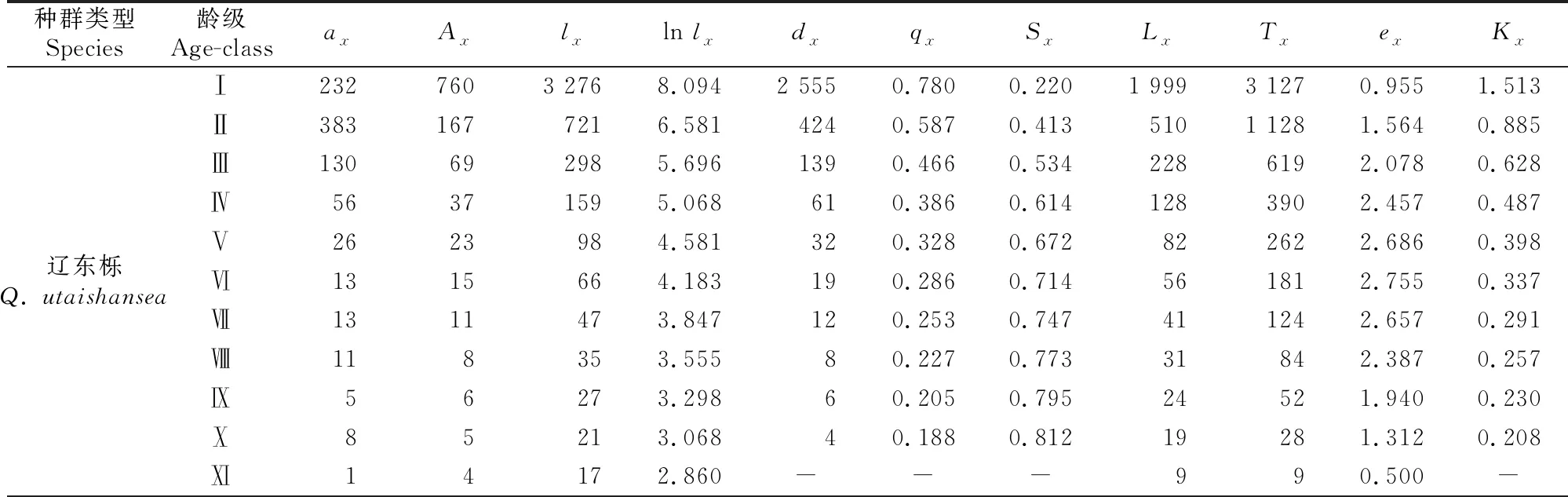
表3 甘南3种次生林优势种种群的静态生命表

表3(续) Continued table 3
辽东栎、红桦、糙皮桦种群死亡率(qx)和消失率(Kx)均在Ⅰ龄级最大,死亡率分别是78.0%,72.0%和76.2%,消失率依次是151.3%,127.3%和143.5%;随着龄级的增加,3种次生林的死亡率和消失率均依次减小,说明由于环境的筛选和种内、种间的竞争,导致幼苗死亡率高,而中龄木和成熟木对环境的适应能力更强。种群期望寿命(ex)反映个体在x龄级的生存能力,3种种群期望寿命值均随着龄级的增加先增大后减小,辽东栎和红桦均在Ⅵ龄级最大,糙皮桦在Ⅴ龄级最大,都表现为在中龄级最大,说明中龄级次生林种群的生存能力最强。
从图2可知,3种次生林种群存活曲线均在低龄级时曲线斜率较大,植株死亡较多,在Ⅴ龄级以后均表现出死亡株数减少,种群呈相对平稳的发展趋势。总体来看,3种次生林种群存活曲线介于Deevery-Ⅱ和Deevery-Ⅲ之间。
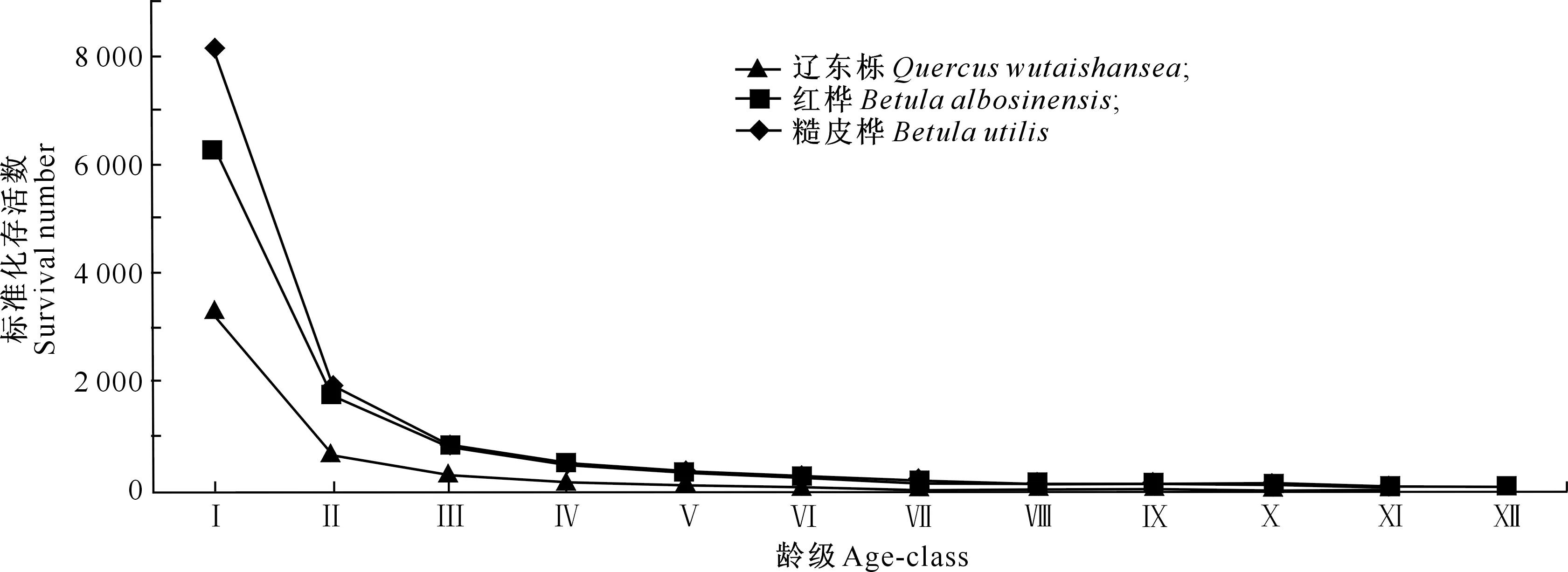
图2 甘南3种次生林优势种种群的存活曲线
采用2种数学模型分别对不同次生林种群的存活曲线进行拟合检验,结果如表4所示。由表4可知,3种次生林种群的存活曲线用幂函数拟合的R2均大于用指数函数拟合,因此3种次生林种群存活曲线更趋近于Deevey-Ⅲ型。
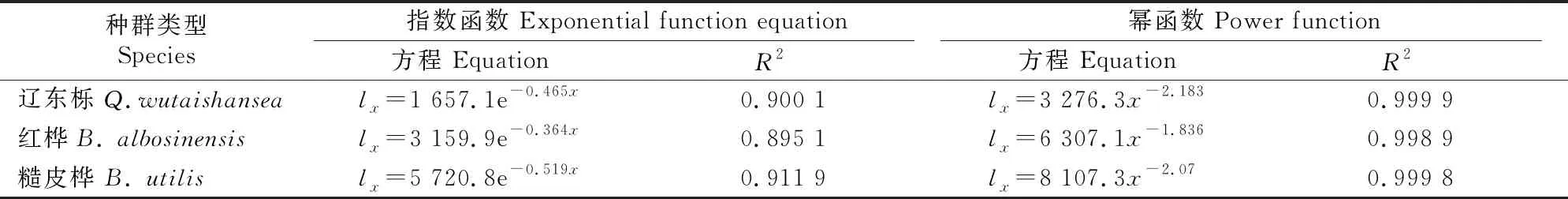
表4 甘南3种次生林优势种种群存活曲线的拟合方程
2.3 3种次生林优势种种群的动态分析和时间序列预测
由表5可知,不同次生林种群相邻龄级的Vn随龄级的增加呈现出不同的变化规律。3种次生林V1均小于0,表明Ⅰ龄级种群个体数少于Ⅱ龄级,这是因为环境承载能力有限以及种群竞争所致。辽东栎在Ⅸ-Ⅹ龄级出现了一次衰退(V9<0),红桦在Ⅷ-Ⅸ和Ⅹ-Ⅺ龄级出现了2次衰退(V8<0,V10<0),这种衰退是由于较小龄级个体数不足造成的;糙皮桦除V1外均大于0,说明种群处于良好的增长状态。3种次生林种群的动态指数Vpi均大于0,以糙皮桦最大,红桦最小,表明种群均为增长型,其中糙皮桦增长潜力最大。3种种群的V′pi也均大于0,且辽东栎最大,红桦最小,表明受外界干扰时各种群也表现为增长型。种群对外界干扰所承担的风险概率极大值Pmax以辽东栎最大,这与各种群的V′pi表现一致,说明辽东栎增长潜力最大但受干扰的概率也最大;红桦和糙皮桦Pmax较小,表明其受外界干扰的概率较小,种群更加稳定。

表5 甘南3种次生林优势种种群的动态变化指数
以各种群不同龄级实际存活数为原始数据,运用一次移动平均法预测未来经过2,4,6,8个龄级后种群各龄级存活个体数,结果如表6所示。由表6可知,3种种群在未来Ⅱ龄级种群个体数呈减小的趋势;随着时间的推移,辽东栎和红桦种群数量偶有衰退,但很快便能得到补充,衰退现象消失。总体来看,3种次生林种群都有较好的发展,种群数量前期锐减但后期稳定,总体更新能力较强,种群呈现出稳定的增长趋势。

表6 甘南3种次生林优势种种群数量动态的时间序列预测
3 讨 论
植物种群结构是种群动态变化规律的体现,也是植物对立地环境条件适应性的反映[33]。3种次生林种群结构变化基本一致,都是Ⅱ龄级幼苗个体最多,Ⅰ龄级少于Ⅱ龄级,从Ⅰ龄级到Ⅱ龄级幼苗个体明显减少,这与郭垚鑫等[12]、陈国鹏等[35]、赵阳等[17]的研究结果一致。对于红桦和糙皮桦次生林而言,产生这种现象的原因,一方面可能是由于红桦和糙皮桦林内郁闭度较高,林下光照不足,种群竞争激烈,环境承载有限,从而导致Ⅰ龄级幼苗大量死亡[36];另一方面,可能是因为红桦和糙皮桦幼苗的更新、生长强烈依赖于林窗斑块[37],而研究区种群生长良好,成熟林死亡个体较少,但幼苗耐阴性差,致使部分Ⅰ龄级幼苗死亡。而对于辽东栎次生林,可能是因为其种群分布于半阳坡,林内光照充足,水分蒸发较快,林下比较干燥,Ⅰ龄级幼苗较少可能是受水分和种群竞争的影响,这与于世川等[37]对黄龙山辽东栎种群结构的研究结果相似。通过负指数分布和q值理论对3种次生林径级分布进行拟合可知,3种群落株数随着径级的增大平稳下降,q值较小,种群总体径级较小,且Ⅰ龄级幼苗较少,后期如果得不到及时补充,种群可能逐渐走向衰退,向老龄化演替[38-39]。种群龄级结构总体呈“钟型”,虽然群落为稳定型,但是各龄级株数分布不合理,需调整径级结构培养稳定的林分状态。
静态生命表可以判断植物种群发展趋势,反映种群现状,展现种群与环境的动态关系,对植物保护和利用具有重要意义[1]。3种次生林种群静态生命表显示,种群期望寿命值随着龄级的增加先增大后减小,呈正态分布。随着种间、种内对环境和营养物质的竞争,致使3种次生林种群在Ⅰ、Ⅱ龄级植株死亡较多,但在竞争中存活下来的植株生存能力更强。各种群死亡率均以Ⅰ、Ⅱ龄级较高,Ⅶ龄级后较低,种群存活曲线均趋向于Deevey-Ⅲ型,这与王道亮等[40]对黄龙山辽东栎种群结构的研究结果一致,但与赵阳等[17]认为甘肃南部辽东栎种群存活曲线为Deevey-Ⅱ型不同,也与冯云等[41]对北京东灵山辽东栎存活曲线呈Deevey-Ⅰ的研究结果不同,可能是立地条件和方法的选择不同所致。赵阳等[17]选择腊子口一个地方,区域具有局限性,且辽东栎种群均以成熟林为主,平均胸径为15.65 cm,而本研究样地设置面较广,种群主要是采伐以后形成的次生林;冯云等[41]选择的是连续山体组成的样带,这也可能导致与本研究辽东栎存活曲线不同。因环境承载能力有限以及种群之间的竞争,种群受外界干扰的风险概率指数也不相同,总体而言,辽东栎种群增长潜力虽然最大,但由于其分布海拔较低,分布区人类活动较多,加之辽东栎是很好的用材树种,所以受人类活动的影响较大,受干扰程度也最大;红桦和糙皮桦受干扰程度相对较小,因此种群更加稳定。
时间序列分析是对种群未来龄级结构动态和发展趋势的预测[21]。本研究对3类种群未来经过2,4,6,8个龄级后的数量动态进行时间序列预测,表明前期各种群数量锐减,但是很快能得到补充,后期稳定,总体种群更新能力强,结构合理,呈稳定的增长趋势。本研究中3个树种均是喜光树种,幼苗耐阴性差,个体死亡率高,幼苗转化成幼树的比例低,这可能导致种群在整个生活史中形成自身生理生长与环境不协同的瓶颈。3种次生林种群数量变化动态存在波动性,除Ⅰ-Ⅱ龄级种群数量变化出现负值外,辽东栎种群还在Ⅸ-Ⅹ龄级出现负值,红桦种群则在Ⅷ-Ⅸ和Ⅹ-Ⅺ龄级2次出现负值,说明辽东栎和红桦种群在后期生长中的数量补充并不是连续和稳定的,或者可能是自身生理障碍的原因,但类似报道尚未见到;糙皮桦种群后期数量变化动态均为稳定连续增长,相比较糙皮桦未来种群更加稳定,结构更加合理。
4 结 论
青藏高原东缘甘肃南部3种次生林优势种种群Ⅰ~Ⅵ龄级个体数之和均占种群个体总数的80%以上,且Ⅱ龄级个体数量最多,林木径级较小,需调整径级结构培养稳定的林分状态; 3种次生林种群结构变化基本一致,存活曲线均为Deevey-Ⅲ型,Ⅰ龄级幼苗个体数量少于Ⅱ龄级,Ⅱ龄级以后总体随着龄级的增大种群个体数减少,但是各龄级成活个体分布不合理,株数相差较大;Ⅰ龄级幼苗死亡率最高,后期降低,种群期望寿命值随着龄级的增加先增大后减小;预测未来3种次生林优势种种群数量变化动态和发展呈稳定增长趋势,但存在波动性,后期生长中个体数量补充并不连续。
猜你喜欢
湖南生态科学学报(2022年4期)2022-11-11 03:44:34
河南农业大学学报(2020年2期)2020-05-22 02:56:30
山西林业科技(2019年1期)2019-05-27 12:24:52
现代农业研究(2017年11期)2018-01-12 21:03:03
辽宁林业科技(2017年4期)2017-06-22 10:23:36
辽宁林业科技(2017年4期)2017-06-22 10:23:36
绿色科技(2016年23期)2017-03-15 15:41:56
现代园艺(2017年20期)2017-02-03 18:49:29
科教导刊·电子版(2016年25期)2016-11-16 10:24:35
江苏农业科学(2016年5期)2016-07-23 15:59:31
