
基于PPARγ/AMPK/NF-κB信号通路研究对羟基苯甲醛对TNBS诱导的小鼠克罗恩病的治疗作用
2022-07-07 09:25许晓飞罗爱林徐笑天王宇晖段小群
天然产物研究与开发 2022年6期
许晓飞,罗爱林,徐笑天,卢 曦,王宇晖,段小群
桂林医学院药学院,桂林 541199
克罗恩病(Crohn’s disease,CD)是一种慢性复发性胃肠道疾病,是炎症性肠病(inflammatory bowel disease,IBD)的主要疾病之一,其发病通常以腹泻、腹痛、直肠出血、发烧、体重减轻和疲劳等常见症状为特征[1]。在CD中,炎症通过肠壁从黏膜延伸到浆膜,这种疾病持续复发和缓解,随着多次复发,CD可以从最初的轻度至中度炎症发展为严重的穿透性(瘘管形成)或狭窄性疾病,更严重者会导致癌变,严重影响患者的生活质量[2]。另外,CD患者还表现出许多肠外炎症,如眼睛、肝脏、皮肤和关节中等炎症反应,反映了这种疾病的全身性[3]。CD的治疗包括一线治疗和其他治疗,其中一线治疗包括类固醇、抗肿瘤坏死因子α(TNF-α),可快速缓解症状。其他治疗可能包括针对IL-12/23或整联蛋白α4β7的单克隆抗体、免疫调节剂、联合疗法或手术[4]。然而,这些药物的统一机制是免疫抑制,长期使用会导致患者感染风险增加,除此之外还伴随其他严重的副作用,包括骨髓抑制、骨质疏松症和肝损伤[5]。因此,迫切需要开发来自自然资源和功能性食品的替代药物来治疗CD。
雷公菌(Nostoccommune)是一种蓝藻生物,由于其营养全面丰富,经常被用于预防和治疗疾病。近年来,其提取物已被证明对肠道疾病有效[6]。我们在前期研究中发现,雷公菌的主要活性成分对羟基苯甲醛(p-hydroxybenzaldehyde,HD)能通过抑制肠道炎症反应改善溃疡性结肠炎(ulcerative colitis,UC)疾病症状[7]。基于UC与CD的诸多相似性,我们推测HD也许会有抗CD作用,为了验证此假设,我们使用2,4,6-三硝基苯磺酸(TNBS)诱导建立小鼠CD模型和TNF-α诱导的Caco-2细胞炎症模型,探究HD对CD的保护作用及机制,为富含HD的食物作为防治IBD的补充替代治疗提供科学依据。
1 材料与方法
1.1 仪器
2-16KL低温高速离心机(Sigma-aldrich公司);80-2C室温低速离心机(常州市凯航仪器有限公司);DSZ5000X荧光倒置显微镜(重庆澳浦光电技术有限公司);EPS 300蛋白电泳、转膜系统(北京原平皓生物技术有限公司);iMark酶标仪、CFX Opus 96 PCR仪(Bio Rad公司);ES-J120分析天平(天津市德安特传感技术有限公司);Tanon 6600全自动化学发光图像分析检测系统(上海天能科技有限公司);NanoDrop 2 000分光光度计(赛默飞世尔科技公司)。
1.2 药物与试剂
HD(C7H6O2,分子量为122.12,纯度≥98%,上海源叶生物科技有限公司,Lot:123-08-0);柳氮磺胺吡啶(SASP,上海源叶生物科技有限公司,Lot:1346606-50-5);TNBS(C6H3N3O9S,分子量为293.17,纯度 ≥5.0%(w/v)水溶液,西亚化学科技有限公司,Lot:23407-1);TNF-α、白介素6(IL-6)、白介素1β(IL-1β)酶联免疫吸附试验(ELISA)试剂盒(武汉新启迪生物科技有限公司,Lot:06460、05884、05874);髓过氧化物酶(MPO)活性测定试剂盒(南京建成生物工程研究所,Lot:044-1-1);Caco-2细胞(美国典型培养物保藏中心);胎牛血清(北京同立海源生物科技有限公司,Lot:07133)、DMEM(Gibco公司,8118251);重组刺激因子TNF-α(R&D Systems公司,Lot:10291);anti-PPARγ、anti-p-AMPK、anti-AMPK、anti-NF-κB p65、anti-p-NF-κB p65和anti-β-actin(Bioworld公司,Lot:4444、4457、1009、1256、4137、6007);RIPA裂解液、5× SDS-PAGE蛋白上样缓冲液和BCA蛋白浓度测定试剂盒(碧云天生物技术研究所,Lot:0013、0015、0012);Super ECL Plus超敏发光液、Hi-script RT Super Mix和Ace Q-PCR SYBR Green Master Mix(南京诺唯赞生物科技股份有限公司,Lot:412-01、123-01、131-02)。
1.3 动物
本实验的所有程序均经过桂林医学院动物伦理委员会的严格审核和批准,动物协议批准文号为GLMC201904008。36只SPF级健康雄性Balb/c小鼠,7周龄,体重20±2 g,由湖南斯莱克景达实验动物有限公司提供,许可证号:SCXK(湘)2016-0002。所有小鼠均在标准条件下(温度:22~25 ℃;湿度:45%~50%;12~12 h光/暗循环),自由采食标准饲料和水。
1.4 方法
1.4.1 动物造模、分组及处理
根据我们前期研究发现剂量为40 mg/kg的HD可以显著改善小鼠UC[7],基于CD与UC具有一定的相似度,我们在TNBS诱导的小鼠CD模型上使用40 mg/kg的HD进行预实验,发现HD可以显著改善小鼠CD,因此选择40 mg/kg作为高剂量组,并将剂量以倍数减少的方式分别选择20 mg/kg和10 mg/kg作为中剂量和低剂量。适应性喂养一周后,将小鼠随机分组正常组(N)、模型组(M)、HD高剂量组(HD-H,40 mg/kg)、HD中剂量组(HD-M,20 mg/kg)、HD低剂量组(HD-L,10 mg/kg)和柳氮磺胺吡啶组(SASP,300 mg/kg),每组6只。如前所述[8],使用TNBS(250 mg/kg)诱导CD小鼠模型。除正常组外,其余各组小鼠实验前均禁食不禁水24 h,实验当天灌肠前挤出小鼠粪便,进一步排空,随后用戊巴比妥钠麻醉小鼠。将灌肠管连接在注射器上,用棉签蘸取花生油,涂抹在灌肠管上;将灌肠管从小鼠肛门插入3.5 cm,之后缓缓注入100 μL含有2.5% TNBS的50%乙醇溶液,同时向外拔出灌肠管,灌肠后将小鼠尾巴用胶带纸粘在桌边,保持倒吊状态4 min,确保肛门无滴液后放回鼠笼。小鼠给予TNBS 1次后,SASP组与HD组小鼠在模型组的基础上每天灌胃给予相应的治疗药物(0.1 mL/10 g),共给药7天。正常组和模型组小鼠给予水作为对照。第7天,给药4 h后,用戊巴比妥钠麻醉小鼠后,收集小鼠距离肛门边缘3 cm处的长度大约为1 cm的结肠组织。
1.4.2 疾病活动性指数(DAI)评分
每天记录小鼠体重变化、大便稠度及粪便隐血情况。DAI评分为(体重减轻+大便稠度+大便隐血)/3[9]。评分见表1。
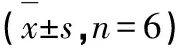
表1 DAI 评分表
1.4.3 结肠组织病理观察
第7天处死所有小鼠,取出结肠,量取长度,纵向打开,用PBS冲洗干净。取部分结肠用4%多聚甲醛固定24 h,经过脱水、石蜡包埋、切片(5 μm)、苏木精-伊红染色(H&E),使用显微镜观察组织病变情况并拍照。并根据杯状细胞耗竭(存在=1,不存在=0)、隐窝脓肿(存在=1,不存在=0)、黏膜结构破坏(正常=1,中度=2,广泛=3),肌肉增厚(正常=1,中度=2,广泛=3)和细胞浸润(正常=1,中度=2,透壁=3)[10]情况来量化结肠组织的损伤程度。
1.4.4 ELISA检测结肠组织和细胞中IL-1β、IL-6、TNF-α蛋白表达
将结肠组织剪碎后,以PBS作为匀浆液将结肠组织置于研磨管中充分研磨,制备结肠组织匀浆,在4 ℃、12 000 r/min条件下离心10 min,分离上清,按照ELISA试剂盒说明书检测小鼠结肠组织匀浆上清中TNF-α、IL-6和IL-1β的蛋白表达水平。另外,采用10 ng/mL的TNF-α诱导Caco-2细胞模型进一步研究HD的抗克罗恩病作用。实验分为对照组(C,无HD和TNF-α)、模型组(M,添加10 ng/mL的TNF-α)、HD低浓度组(HD-L,添加10 ng/mL的TNF-α和3 μmol/L的HD)、HD中浓度组(HD-M,添加10 ng/mL的TNF-α和10 μmol/L的HD)、HD高浓度组(HD-H,添加10 ng/mL的TNF-α和30 μmol/L的HD)和柳氮磺胺吡啶组(SASP,添加10 ng/mL的TNF-α和30 μmol/L的SASP)。化合物处理细胞24 h后,收集各组培养基并根据ELISA试剂盒说明书检测细胞中TNF-α、IL-6和IL-1β的蛋白表达水平。
1.4.5 结肠组织MPO活性检测
收集结肠组织,称重、剪碎匀浆后,按照试剂盒说明书操作,检测结肠组织中的MPO活性,该试剂盒以每克湿组织的重量为单位。
1.4.6 小鼠结肠和Caco-2细胞中TNF-α、IL-6和IL-1β的mRNA表达
采用Trizol试剂从结肠组织和Caco-2细胞中提取总RNA。使用Nano Drop 2000分光光度计量化总RNA。随后,用Hi-script RT Super Mix将RNA转录为cDNA。以cDNA为模板,在95 ℃,10 s和60 ℃,30 s的条件下,用Ace Q-PCR SYBR Green Master Mix和MyiQ2检测系统检测TNF-α、IL-6、IL-1β的mRNA表达,共40个循环。目的基因的表达以GAPDH作为内参,数据分析使用2-ΔΔCt方式计算,平行三次实验。引物序列及长度见表2。

表2 引物序列
1.4.7 小鼠结肠中PPARγ/AMPK/NF-κB信号通路相关蛋白表达
结肠组织用PBS清洗后,用滤纸吸干水分,准确称量40 mg样品,使用含有PMSF的RIPA裂解液提取组织和Caco-2细胞总蛋白。根据制造商的方案,使用BCA蛋白检测试剂盒检测蛋白总浓度。制备10%的电泳凝胶,将等量蛋白样品加到凝胶中,然后通过80 V,30 min和120 V,50 min的电泳条件进行蛋白分离。根据目标蛋白的分子量,将所需蛋白转移到PVDF膜上。然后在添加5%脱脂奶粉的TBST缓冲液中室温封闭膜2 h。随后,PVDF膜与一抗在4 ℃条件下孵育过夜。次日,用TBST缓冲液冲洗后,室温下用二抗避光孵育1.5 h。通过ECL系统检测免疫反应性蛋白质,用全自动化学发光图像分析检测系统下收集图片。β-actin用作蛋白标准化对照。最后,通过Quantity One软件量化条带中的蛋白质强度。
1.4.8 统计分析

2 结果
2.1 HD对TNBS诱导的CD小鼠体重、腹泻、隐血和结肠长度的影响
为了确定HD是否具有改善TNBS诱导的小鼠克罗恩病疾病症状的作用,我们对克罗恩病小鼠灌胃HD治疗7天,并以SASP为阳性药物。结果如图1所示,与正常组相比,使用TNBS可以导致小鼠体重出现明显减轻、严重的腹泻和血便,说明建模成功。然而给予HD干预后,大大阻止小鼠体重降低(见图1A)、DAI评分升高(见图1B)以及结肠缩短(见图1C)。同时研究发现,HD(40 mg/kg)的作用优于SASP(100 mg/kg),说明HD具有改善CD小鼠结肠疾病症状的作用。

图1 HD对TNBS诱导的CD小鼠体重、腹泻、潜血和结肠长度的影响
2.2 各组小鼠结肠组织形态学变化
为了研究HD对各组小鼠结肠组织形态学变化的影响,显微镜下观察各组小鼠结肠组织形态学,结果如图2所示,正常组小鼠结肠表现为黏膜和杯状细胞形态正常,未见炎性细胞浸润与溃疡。模型组小鼠结肠组织黏膜上皮出现缺失,大量杯状细胞被破坏,黏膜下层明显增厚水肿,血管增生,黏膜层及黏膜下层可见较多的炎症细胞浸润,另外淋巴组织增生,黏膜层有裂隙状溃疡形成,并且部分溃疡可深达肠壁肌层。然而,HD组发现小鼠的黏膜上皮较为完整,少见炎性细胞浸润,杯状细胞多数表现为完整,腺体排列较为规则,黏膜下层增厚减轻,组织水肿减少。以上说明HD具有改善小鼠结肠病理情况。

图2 HD对各组小鼠结肠组织形态学变化的影响(×200)
2.3 HD对结肠炎症反应的影响
为了证明促炎因子在克罗恩病的炎症反应中发挥重要作用,我们研究了促炎因子在TNBS诱导的小鼠CD中的表达。如表3和4所示,相对于对照组,TNBS增加了结肠中TNF-α、IL-6和IL-1β蛋白质和mRNA的表达。HD干预后,显著抑制TNF-α和IL-6蛋白质和mRNA的表达,且呈剂量依赖性,然而不影响IL-1β的表达。研究还发现,HD(40 mg/kg)几乎可以使TNF-α和IL-6的表达恢复到正常水平。另外,体外实验也进一步证明了HD的抗结肠炎炎症作用。以上表明,HD的抗CD作用与其抑制促炎因子的表达密切相关。

表3 HD对TNBS诱导的CD小鼠结肠组织中促炎因子mRNA表达的影响

表4 HD对TNBS诱导的CD小鼠结肠组织中促炎因子蛋白质表达的影响
表5 HD对TNF-α诱导的Caco-2细胞炎症模型中促炎因子mRNA表达的影响
表6 HD对TNF-α诱导的Caco-2细胞炎症模型中促炎因子(TNF-α、IL-6和IL-1β)蛋白质表达的影响
2.4 HD对TNBS诱导的CD小鼠结肠MPO活性的影响
MPO是由中性粒细胞嗜酸性颗粒表达的一种血液蛋白,通常被用作中性粒细胞的标记物。为了探究HD对CD小鼠结肠MPO含量表达的影响,通过试剂盒检测小鼠结肠中MPO的表达水平。结果发现,与正常组相比,TNBS处理导致小鼠结肠MPO活性升高(P<0.01)。而HD干预后,可以显著抑制MPO的活力,且呈剂量依赖性。HD(40 mg/kg)抑制MPO作用优于SASP(见图3)。以上说明HD可以抑制中性粒细胞在CD小鼠结肠的浸润。

图3 HD对TNBS诱导的CD小鼠结肠MPO活性的影响
2.5 HD对PPARγ/AMPK/NF-κB信号通路相关蛋白表达的影响
为了检测HD是否通过PPARγ/AMPK/NF-κB信号通路改善小鼠克罗恩病,通过蛋白免疫印迹法检测小鼠结肠中PPARγ/AMPK/NF-κB信号通路相关蛋白表达。结果发现(见图4),与正常组相比,模型组小鼠结肠组织中PPARγ、p-AMPK蛋白表达水平显著降低(P<0.01),p-NF-κB p65蛋白表达水平显著升高(P<0.01)。与模型组相比,SASP和HD均可以使结肠组织中PPARγ、p-AMPK蛋白表达水平显著升高(P<0.01),p-NF-κB p65蛋白表达水平显著降低(P<0.01),其中对HD(40 mg/kg)效果最佳。另外,在TNF-α诱导的Caco-2细胞炎症模型中发现,HD对PPARγ/AMPK/NF-κB信号通路相关蛋白表达的影响与动物实验结果相一致(见图5)。以上结果表明,HD通过激活PPARγ促进AMPK磷酸化抑制NF-κB活化,进而改善小鼠CD症状。

图4 HD对结肠中PPARγ/AMPK/NF-κB信号通路相关蛋白表达的影响

图5 HD对细胞中PPARγ/AMPK/NF-κB信号通路相关蛋白表达的影响
3 讨论与结论
CD是一种病因不明的特发性慢性炎症性疾病,易复发,累及口腔到肛门的整个消化道。在欧洲和北美人群的发病率最高,但近年来亚洲人群的发病率急剧上升[11]。研究发现,CD的发病机制与肠黏膜免疫反应失调引起的肠道炎症密切相关[12]。多项研究表明,改善肠道炎症反应可以显著缓解CD。例如,沙利度胺通过恢复TH17/Treg细胞的失衡和改变促炎和抗炎细胞因子的平衡减轻炎症反应来改善小鼠CD[13]。马基多酚提取物通过下调COX-2和iNOS的表达抑制肠道炎症改善小鼠CD[14]。以上说明抑制肠道炎症反应是治疗CD的一个主要手段。一致地,我们发现HD可以通过抑制炎症反应来改善TNBS诱导的CD。
炎症是IBD发病的主要诱因,引起IBD肠组织损伤。作为重要的炎症生物标志物,相关细胞因子包括TNF-α、IL-1β、IL-6和IL-17,它们在炎症反应中起重要作用,抑制其过表达可以改善CD的肠道炎症反应[15]。例如LB可以抑制TNBS诱导的小鼠肠道IL-6、IL-1β、TNF-α的过表达改善克罗恩病[16]。丁酸钠在TNBS诱导的炎症性肠病小鼠模型中抑制炎症并维持上皮屏障完整性[10]。以上研究均表明抑制促炎细胞因子的产生对CD具有治疗作用。在这项研究中,TNBS会诱发小鼠出现肠道严重炎症反应。然而,HD治疗后减轻了结肠炎症细胞浸润,并且在动物和细胞水平上均显著下调了这些炎性细胞因子水平,说明HD可以抑制炎症反应改善小鼠CD。
NF-κB是IBD炎症细胞因子转录的重要信号转导途径,负责调节促炎细胞因子(TNF-α和IL-6)的转录[17]。作为NF-κB家族成员之一,p65一旦被激活可以转位到细胞核中上调促炎细胞因子的转录表达[18]。研究发现,通过抑制NF-κB信号通路可以减轻TNBS模型中的炎症[15]。在本研究中,TNBS的诱导使CD小鼠结肠中NF-κB p65的磷酸化蛋白表达增多,而HD的干预导致NF-κB p65的磷酸化蛋白表达降低。另外,HD还可以抑制TNF-α诱导的Caco-2细胞中NF-κB p65的磷酸化蛋白表达的升高。说明HD通过抑制NF-κB信号通路改善结肠炎症反应。
AMPK是一种丝氨酸/苏氨酸激酶,在葡萄糖和脂质代谢、细胞生长和凋亡、自噬和炎症中发挥重要作用[19]。研究发现,AMPK通过抑制NF-κB的激活来实现抗炎作用[20]。Xu等[21]发现AMPK的激活可以抑制NF-κB通路进而抑制缺氧和复氧过程中的炎症反应。Chen等[22]发现,使用化合物C(AMPK抑制剂)可以阻止红景天苷抑制NF-κB信号通路改善AGEs诱导的内皮炎症和氧化应激。同样,本研究发现HD具有促进AMPK磷酸化的作用,且呈剂量依赖性。以上说明HD通过激活AMPK抑制NF-κB通路从而改善TNBS诱导的炎症反应。
PPARγ是一种转录因子,在抗炎、抗氧化和吞噬细胞介导的清除过程中起重要作用,在结肠上皮中高度表达[23]。许多研究表明,PPARγ是AMPK的激活因子,而PPARγ的抗炎作用是通过激活AMPK来实现。例如,PPARγ激动剂(噻唑烷二酮药物)可以通过AMPKα的磷酸化激活AMPK抑制肠道病理性炎症[24]。而GW9662、T0070907和PPARγsiRNA可阻止羟基积雪草酸激活AMPK恢复Th17/Treg平衡来改善结肠炎[25]。本研究发现HD具有PPARγ激动剂作用,说明HD通过激活PPARγ抑制CD的炎症反应。
综上所述,HD可有效减缓TNBS诱导的小鼠CD炎症反应,其机制可能与PPARγ/AMPK/NF-κB信号通路介导的炎症反应有关。我们的研究表明,在日常生活中补充雷公菌不仅可以防治UC,对CD也有一定的防治作用,在IBD发病率逐年增高以及当前治疗药物诸多局限的大背景下,我们的研究为富含HD的食物作为防治IBD的补充替代治疗提供科学依据。
猜你喜欢
中国交通信息化(2022年8期)2022-10-28
中国种业(2022年9期)2022-10-13
福建医科大学学报(2022年3期)2022-09-10
中华实用诊断与治疗杂志(2022年1期)2022-08-31
基层中医药(2022年1期)2022-07-22
九江学院学报(自然科学版)(2022年2期)2022-07-02
医学食疗与健康(2022年2期)2022-04-23
现代临床医学(2022年2期)2022-04-19
天津医科大学学报(2021年4期)2021-08-21
现代临床医学(2021年1期)2021-01-26
