
不同诱导系对不同基因型玉米材料的诱导率评价
2022-10-13 03:38梁建斌张旭辉闫治斌李世风支海林梁歌恒李丽仙夏瑞英柴君臣
中国种业 2022年9期
梁建斌 张旭辉 闫治斌 李世风 支海林 梁歌恒 李丽仙 李 悦 王 倩 夏瑞英 柴君臣
(甘肃省敦煌种业集团股份有限公司研究院,酒泉 735000)
玉米作为中国第一大粮食作物,在国民经济及国家粮食安全中发挥着重要的作用。随着生产需求的不断增长,玉米育种者不断探索和改进现代育种技术。玉米双单倍体(DH,Doubled haploid)技术是生物技术与常规育种方法结合产生的一条新的育种途径,作为快速获得纯系的手段,可以显著缩短育种年限,加快育种进程[1]。种质狭窄是影响玉米产量提高的主要原因,也是导致我国育种进程、育种水平与发达国家差距较大的主要原因。外来种质是玉米育种重要的种质基础,单倍体育种对外来种质进行遗传改良具有重要意义。单倍体育种是目前各大种业公司广泛应用的现代育种技术,已成为现代玉米育种的关键核心技术之一[2]。
诱导系诱导获得单倍体,是指利用诱导率较高的诱导系作父本、改良基础群体材料作母本,诱导产生单倍体,是目前应用最多的实用技术[3]。其中选择诱导系与基础材料杂交,单倍体诱导系是一个关键因素,1959 年Coe[4]发现了第一个单倍体诱导系Stock6,诱导率仅为2%左右,诱导率低严重制约了该项技术的应用。前人研究表明,不同的诱导系对不同的基础材料诱导时诱导率差异较大[5],选择诱导率高且稳定的诱导系及更适应诱导的具有优良性状的基础材料至关重要,我国玉米单倍体育种总体上还处于引进熟化创新阶段,与发达国家比较还有相当大的差距。到目前为止单倍体诱导系的研究在国内外学者的共同努力下已经有了重大进步,已经选育出非常有价值的高油性诱导系列CAUHOI、农大高诱号系列、吉高诱系3 号和单倍体诱导系H01~H05、WS14、Zarodyshevy Marker Saratovsky、Korichnevv Marker Saratovsky、Moldovian Haploid Inducer 等[6-13]。目前国内诱导系的遗传背景普遍较为单一,所以不仅要组配更具有杂种优势的杂交诱导系,还需要拓宽国内种质资源的遗传背景。前人已有广泛的研究来提高单倍体诱导率和单倍体诱导系的农艺性状,除了提高诱导系的诱导率外,研究诱导材料对单倍体诱导系的影响也很重要[14]。本研究用不同的诱导系诱导不同的基础群体材料,分析诱导率和杂株率差异,探讨诱导系的选用策略,筛选标记明显、诱导率稳定且高、杂株率少的诱导系和基础群体材料。本研究结果可充实单倍体的规模化生产和提高育种效率相关的研究。
1 材料与方法
1.1 试验材料供试的4 个单倍体诱导系材料(表1)均具有籽粒Navajo 标记,均引自中国农业大学国家玉米改良中心。供试的12 个基础材料类型见表2。
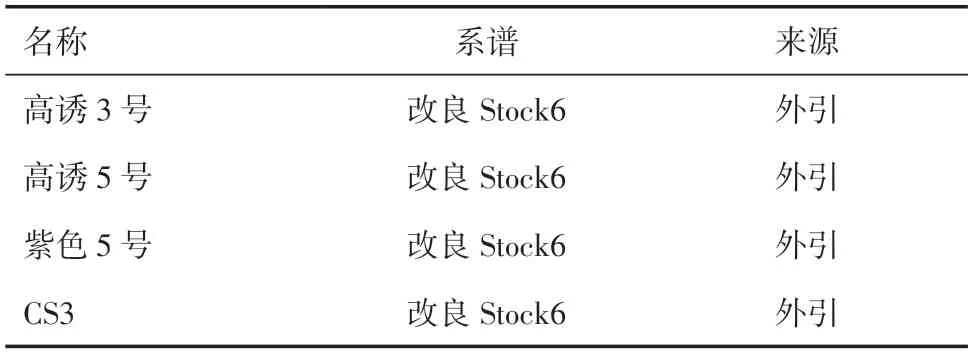
表1 供试单倍体诱导系材料

表2 供试基础材料类型
通过杂交诱导所得籽粒主要有以下3 种类型(图1)。拟单倍体籽粒:籽粒胚乳上有紫色,胚部无色;杂合二倍体籽粒:籽粒胚部和胚乳上均有紫色;杂合二倍体黄粒:籽粒胚乳黄色,胚部无色。
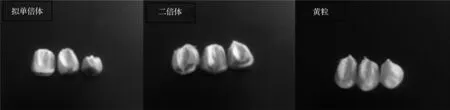
图1 杂交诱导籽粒类型
1.2 试验设计试验于2020 年冬天在甘肃省敦煌种业海南玉米育种基地进行。为了保证诱导系与基础群体材料的花期能够相遇,先播种全部基础群体材料与第1 期诱导系,第2 期、第3 期诱导系分别间隔5d、10d 播种。花期采用人工去雄并套袋授粉的方式,分别取4 个诱导系(高诱3 号、高诱5 号、紫色5 号和CS3)的花粉授予12 个基础群体材料。收获后依据籽粒的Navajo 标记进行籽粒挑选,胚乳上有诱导标记、胚部无诱导标记的籽粒被认定为拟单倍体籽粒,并统计拟单倍体籽粒、有诱导标记的二倍体籽粒数量。2021 年在敦煌种业西北育种基地(酒泉)对拟单倍体籽粒进行催芽、加倍和田间单株移栽。待植株散粉前对植株进一步鉴定,植株高大粗壮或带紫色标记的为二倍体植株,植株矮小,叶片短小、皱缩、上冲的为单倍体植株,并统计单倍体植株数。
1.3 项目统计统计诱导总粒数、拟单倍体籽粒数和田间鉴定单倍体株数,按以下公式计算诱导率和杂株率,杂株包含非单倍体和发育不正常的籽粒。
诱导率(%)=田间鉴定单倍体株数/总粒数×100
杂株率(%)=(拟单倍体籽粒数-田间鉴定单倍体株数)/拟单倍体籽粒数×100
2 结果与分析
2.1 不同诱导系诱导率和杂株率统计如表3 所示,本研究中杂交诱导后籽粒的数量较大,总共有845354 粒,各基础材料都在5269~52060 粒之间,平均17612 粒,统计所得诱导率和杂株率具有代表性。
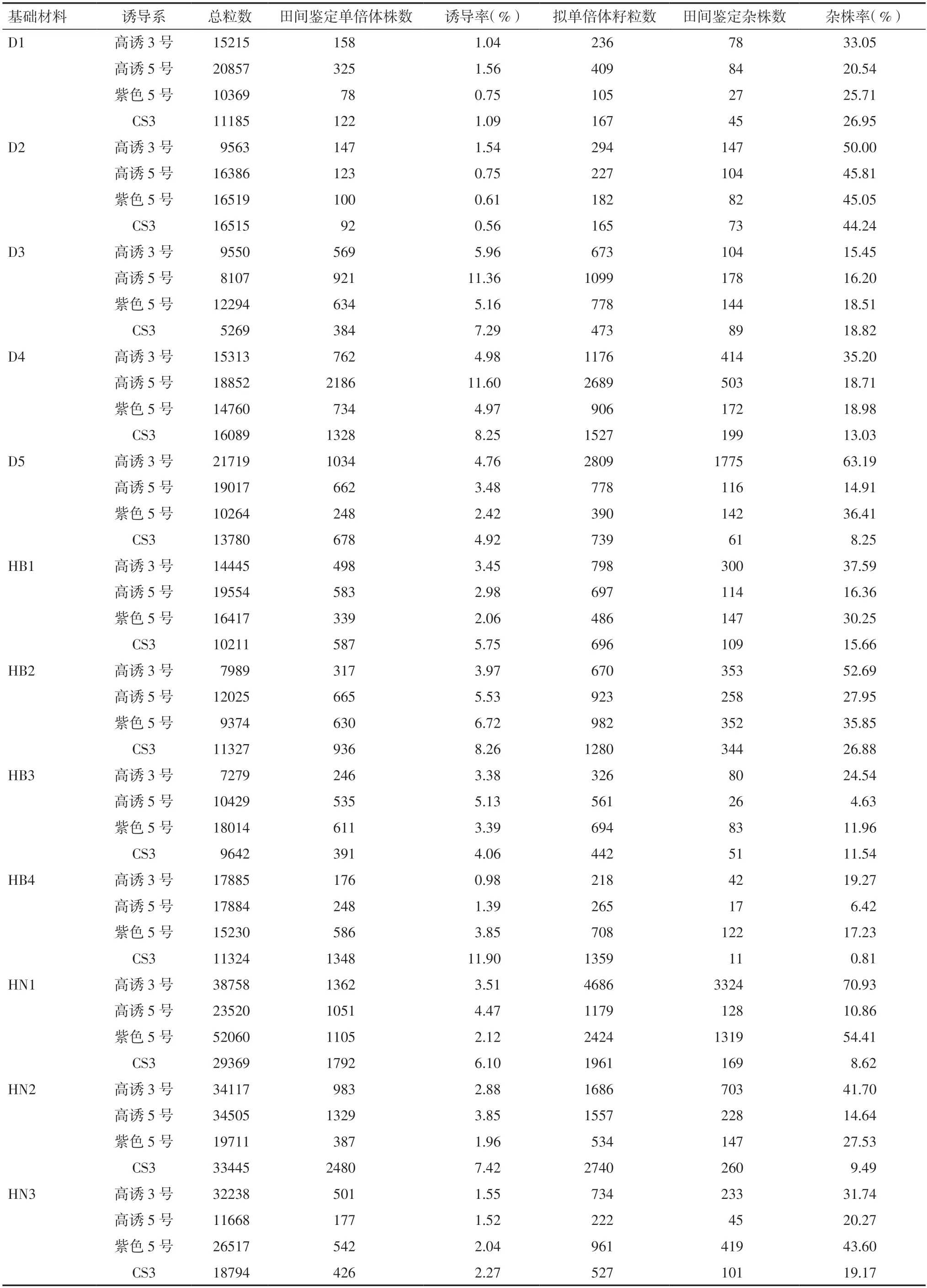
表3 不同诱导系诱导率和杂株率统计
2.2 不同诱导系诱导不同基础群体材料的诱导率由表4 可知,4 个诱导系作父本与12 个基础材料杂交诱导后,不同基础群体材料之间的诱导率存在差异,基础材料D3 和D4 平均诱导率较高,分别为7.44%和7.45%,其中高诱5 号对D3 和D4 基础群体的诱导率分别达11.36%和11.60%。基础群体D1、D2 和HN3 的平均诱导率较低,分别为1.11%、0.87%和1.85%。4 种诱导系对HB4 基础群体的诱导率差异较大,变异系数达112.03%,其中CS3 对HB4 基础群体的诱导率达11.90%。
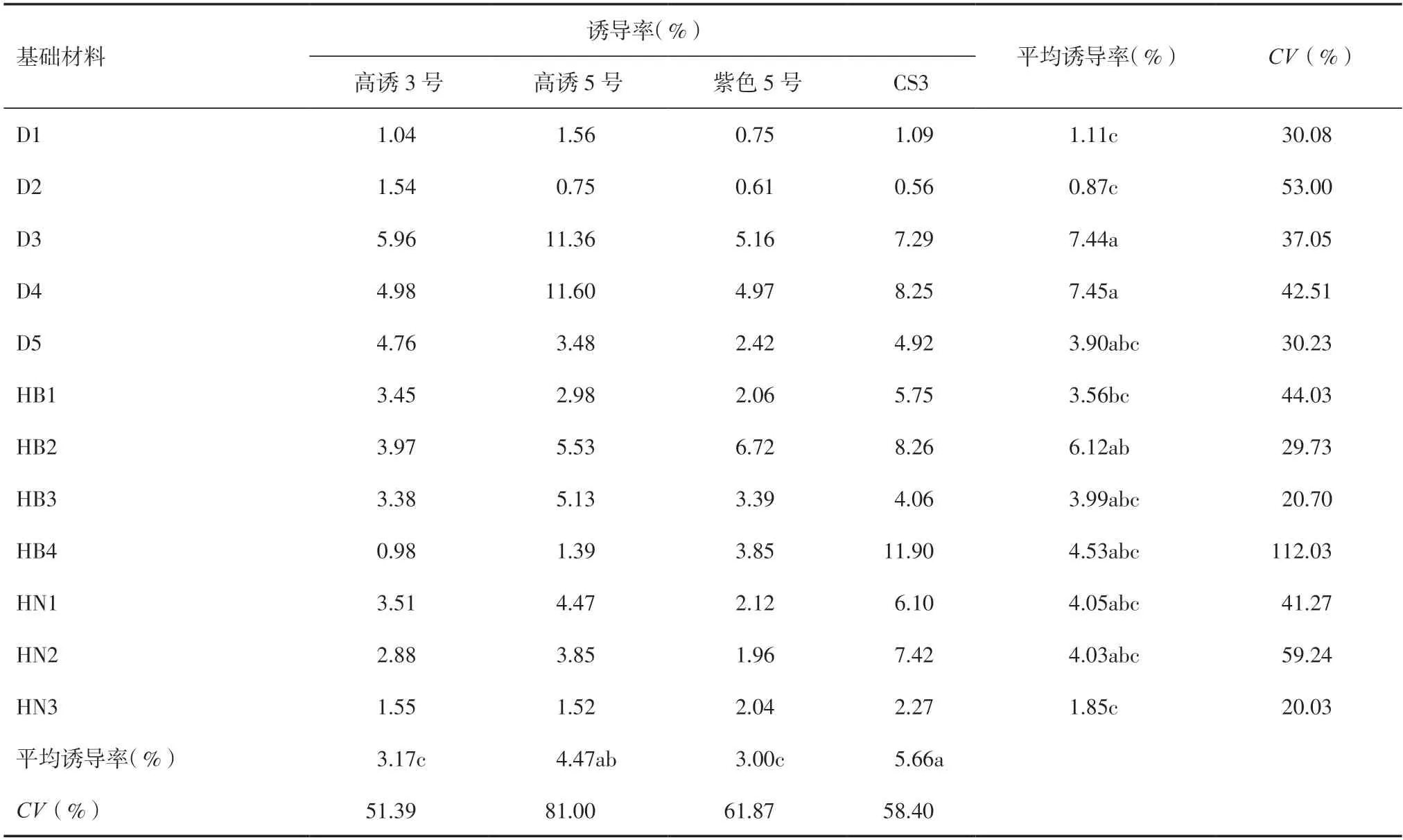
表4 不同诱导系诱导不同基础群体诱导率分析
同一诱导系诱导不同的基础材料后平均诱导率大小依次为CS3>高诱5号>高诱3号>紫色5号,分别为5.66%、4.47%、3.17%和3.00%;高诱3 号和CS3 分别对12 个基础材料的平均诱导率差异相对较小,高诱5 号对12 个基础材料的平均诱导率差异较大,变异系数达81.00%。
2.3 不同诱导系诱导不同基础群体材料的杂株率
如表5 所示,不同诱导系和不同基础材料杂交诱导的杂株率差异较大。不同基础群体材料之间,D2基础群体平均杂株率最大,达46.28%;HB4 基础群体最小,为10.93%;HN1 的变异系数最大,说明1种诱导系诱导HN1 时杂株率差异较大,其中CS3 诱导HN1 基础群体时的杂株率为8.62%,而高诱3 号诱导HN1 基础群体时的杂株率为70.93%;D2 的变异系数最小,说明4 种诱导系诱导D2 基础群体时杂株率差异较小。同一诱导系对不同基础材料诱导时平均杂株率大小依次为高诱3 号>紫色5 号>高诱5 号>CS3,其杂株率分别为39.61%、30.46%、18.11%和16.96%,其中CS3 诱导HB4 时杂株率仅为0.81%。
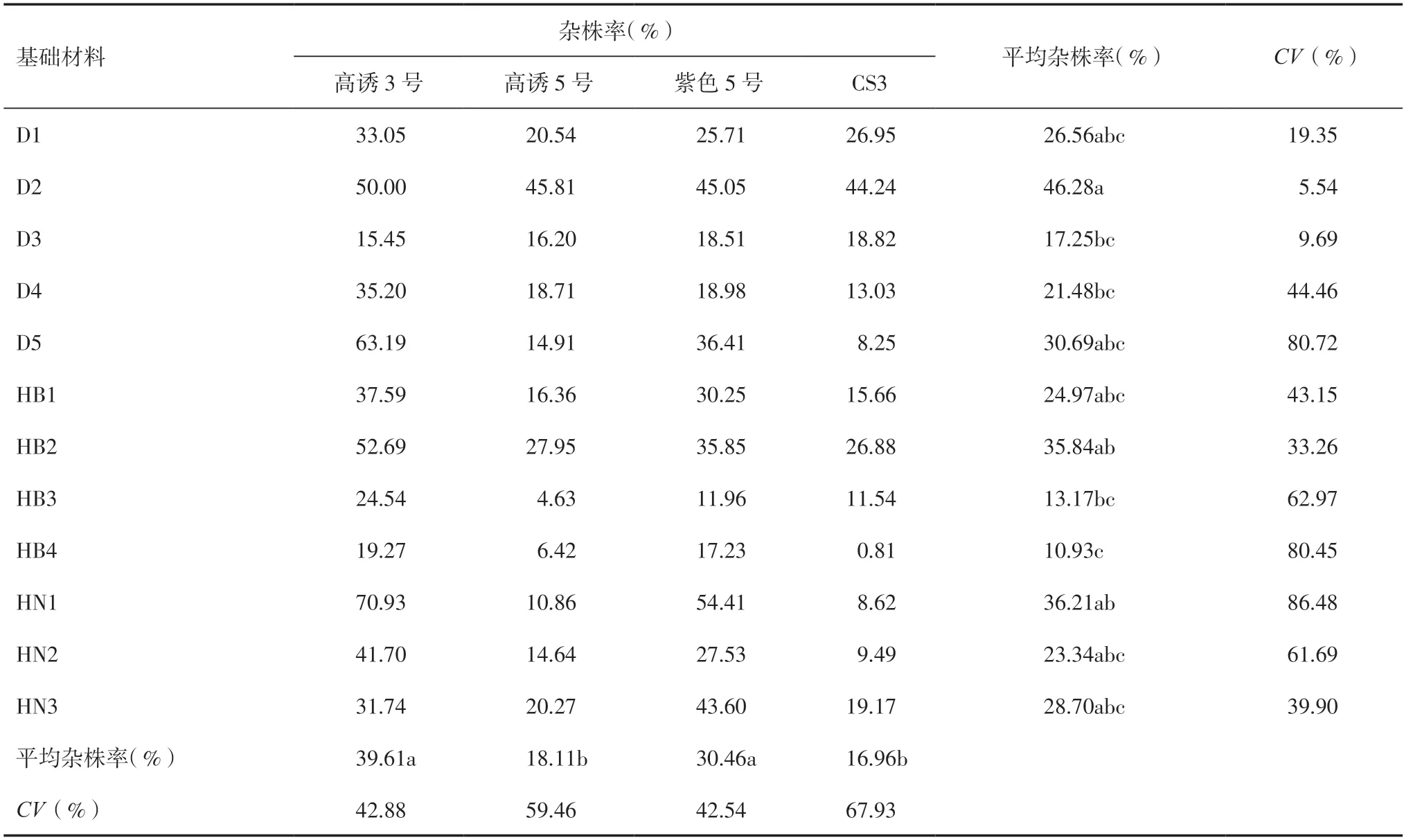
表5 不同诱导系诱导不同基础群体杂株率分析
3 讨论与结论
诱导系的选择首先注重于提高诱导率,从而产生大量单倍体,提高配子体选择效率,筛选聚集更多有利基因的目标植株。目前,基于Navajo 标记的单倍体籽粒筛选主要依靠人工目测,遗传标记在不同种质资源中的表达水平差异很大,影响诱导系的利用效率和育种成本。因此,选择和诱导在注重提高诱导率的同时,还需要加大筛选标记表达的力度。
不同群体材料之间的诱导率存在差异,研究诱导率高的诱导材料对提高诱导效率具有重要意义[15]。本研究发现,同一基础材料与不同的诱导系进行杂交诱导后所得到的单倍体诱导率存在显著差异,12 份基础材料诱导后,D3 和D4 的平均诱导率显著高于其他基础材料,杂株率也较低;D3 和D4材料与诱导系高诱5 号进行组合后其诱导率分别达到11.36%和11.60%,这一最佳组合显著提高了其诱导率,其可能原因是D3 和D4 的遗传基础更为复杂,优良的血缘使得其适应性更强、育性恢复能力强,使得诱导率偏高,但具体原因还需进一步研究,以期将此材料广泛应用于单倍体诱导的基础材料来满足商业育种的需求。不同诱导系对同一基础材料进行杂交诱导所得单倍体诱导率存在显著的差异,本研究发现4 个诱导系单倍体平均诱导率在3.00%~5.66%之间,这一研究结果与李高科等[16]研究结果相一致。其中CS3 的诱导能力最强,变异系数也小于高诱5 号和紫色5 号,说明其诱导系的稳定性较高,平均杂株率也低于其他诱导系,表明在玉米单倍体诱导育种中,可继续加大对CS3 的利用。本研究结果表明,基础材料和诱导系都会对玉米单倍体诱导率和杂株率产生一定影响,因此诱导系的诱导能力不仅受自身遗传背景的影响,也受其诱导的基础材料遗传背景的影响,但由于本研究所用的基础材料有限,还需进一步针对玉米单倍体进行专用型诱导系的选育以及基础材料的筛选,这是本课题组下一阶段的研究重点。
目前,虽然单倍体技术已广泛应用于商业化育种,但仍有以下4 个方面有待改进。首先,应选择具有优良农艺性状的高频单倍体诱导系。其次,在新的诱导系中整合多个有效标记,提高单倍体鉴定的有效性和准确性。第三,结合倍率相关的QTL 定位和基因编辑技术,建立高效的倍增体系。最后,利用组织培养和选择标记缩短DH 系生产所需的时间。今后,解决上述方面的技术难题,将促进单倍体诱导技术在现代玉米育种中得到更广泛的应用。
猜你喜欢
中国农学通报(2022年13期)2022-05-31
华人时刊(2022年3期)2022-04-26
中学生数理化·高一版(2021年12期)2021-09-05
作文·小学低年级(2021年10期)2021-01-25
大众健康(2020年6期)2020-07-06
新班主任(2020年2期)2020-05-03
群众(2018年18期)2018-10-26
现代农业科技(2018年13期)2018-10-20
科学种养(2017年6期)2017-06-13
农村百事通(2016年8期)2016-05-14
