
深黄伞形霉与黑曲霉混合固态发酵麸皮积累油脂
2022-06-27 02:14王子硕周化岚SarmanOktovianusGultom张建国
工业微生物 2022年2期
郑 潇, 冯 豪, 于 洁, 王子硕, 周化岚,Sarman Oktovianus Gultom, 张建国*
1.上海理工大学健康科学与工程学院,食品科学与工程研究所,上海200093;
2.Department of Agricultural and Biosystems Engineering,Papua University Manokwari,Papua Barat,Indonesia
油脂作为人类生活和生产活动中不可缺少的物质[1],不仅是构成和维持细胞生命的重要大分子物质,还是人类重要的能量物质来源。我国是一个人口大国,油脂供应短缺,根据现有资源无法满足社会需要。据统计,2014年我国食用油消费量达3 167.4万吨,约60%油脂的缺口需要通过进口[2]。因此,人们正在积极开发新的油源途径。而深黄伞形霉(Umbelopsis isabellina,U.isabellina.曾用名深黄被孢霉和Mortierella isabellina,M.isabellina)可以利用多种糖生长,有广泛的底物适应性和油脂积累潜力[3-5]。在日本,深黄伞形霉被用于生产单细胞油脂,这种单细胞油脂被用作饮料和糖果中的食品添加剂,并且以片剂的形式作为一种功能性食品[6,7]。这就消除了大规模使用深黄伞形霉的顾虑。并且,深黄伞形霉所需的营养简单,能利用碳水化合物进行发酵合成自身胞内油脂,油脂含量可达86%,尤其富含油酸,亚油酸和γ-亚麻油酸[8]。
黑曲霉不仅是一种广泛用于固体发酵的丝状真菌,而且是FDA批准的食品安全性好的工业微生物菌株。目前已广泛用于柠檬酸、葡萄糖酸等[9]食品组分的工业生产。此外,黑曲霉可产出纤维素酶、纤维二糖酶、木聚糖酶、淀粉酶、蛋白酶、糖化酶、果胶酶、柠檬酸合成酶[10-15]等多种酶,并可用来水解多种木质纤维素类物质的废弃物,例如土豆、菠萝、豆渣、豆角梗、玉米芯、木薯、甘蔗渣、咖啡壳[16-18],甚至可以水解人工的食品聚合物(聚氨酯)[19]。
固态发酵是微生物将生物质转化为油脂的一种简单方法,固态发酵是指没有或有少量自由水存在下一种或多种微生物在固态基质上生长的生物反应过程。固态发酵已经广泛用于工农业生产:例如微生物代谢物、工业用酶、中国传统食品、以及油脂。固态发酵的优点为操作方式简单、成本低,是一种容易推广的处理方式。固态发酵条件下,丝状真菌发酵示意图如下(见图1):
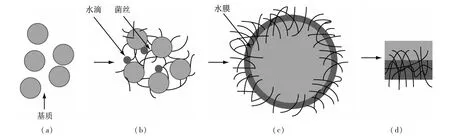
图1 丝状真菌在固态发酵时生长示意图
研究也发现,制约固态发酵深黄伞形霉进一步积累油脂的因素是产酶能力不足。添加外源纤维素酶后油脂产量明显提高。ZHANG[20]将玉米芯经过液氨预处理后利用酶水解释放单糖时发现酶的需要量是50 FPU/g葡聚糖的纤维素酶,30 CBU/g葡聚糖的纤维二糖酶。所以,纤维素酶是富油微生物转化食品废弃物为油脂的必要条件。依据以上的分析,提出利用深黄伞形霉和黑曲霉在麸皮[21]上进行固态发酵积累油脂的思路,因为混合培养固态发酵不仅有利于丝状真菌的生长和酶的分泌,也有利于丝状真菌在基质上的附着和对基质的渗透。结合黑曲霉高效产多种酶和深黄伞形霉易积累油脂的优点将食品废弃物转化为油脂,并且黑曲霉和深黄伞形霉两株菌混合培养有利于食品废弃物中糖的释放、转化过程中抑制物的去除以及油脂产量的提高。可以为食品中废弃物资源的转化以及可再生能源的研究提供参考。
1 材料与方法
1.1 材料和仪器
深黄伞形霉(Umbelopsis isabellina,Mortierella isabellina NBRC 7884)购自日本技术与评价研究所的生物资源中心(NBRC)、黑曲霉(Aspergillus niger ATCC 1015)购自美国典型生物资源保藏中心(ATCC)、小麦麸皮(39.28%C、2.47%N)来自山东菏泽牡丹区,化学试剂购买自国药化学试剂有限公司。
光学显微镜CX40(奥林巴斯公司);气质联用仪7890A(Agilent Technologies公司);分析天平ML104/02(梅特勒公司);高压灭菌锅HVE-50(Hirayama);全温振荡器TQZ-312(上海精宏实验设备有限公司)。
1.2 实验培养基
PDA培养基(g/L):称取3.7 g PDA固体溶于100.0 mL去离子水,121℃、15 min灭菌后备用。
察氏培养基(g/L):25.0无水葡萄糖,30.0 NaNO3,10.0 K2HPO4·3H2O,5.0 KCl,5.0 MgSO4·H2O,0.1 FeSO4,调节至pH 5.6,121℃、15 min灭菌后备用。
1.3 实验方法
1.3.1 深黄伞形霉孢子液的制备
将-20℃下贮藏的深黄伞形霉孢子液接种到PDA培养基上,在28℃下倒置培养,分别在7 d、9 d和11 d不同成熟度时收集孢子,用去离子水洗下,再以20%的甘油保藏,用血球板计数后备用。
1.3.2 黑曲霉孢子液的制备
将-20℃下贮藏的黑曲霉孢子液接种到察氏培养基平板上,在28℃下倒置培养,分别在7 d、9 d和11 d不同成熟度时收集孢子,用去离子水洗下,再以20%的甘油保藏,血球板计数备用。
1.3.3 初始含水量对双菌培养积累油脂的影响
小麦麸皮为发酵原料,称取30 g于250 mL三角瓶中,麸皮含水量分别50%、67%、75%和80%,121℃高压灭菌20 min,然后按照深黄伞形霉∶黑曲霉(1∶1)将两种菌混匀,孢子数为106CFU/g麸皮,28℃培养6周,每周定时取样,取样烘干保留,测定油脂含量。
1.3.4 孢子数对双菌积培养累油脂的影响
小麦麸皮为发酵原料,称取30 g于250 mL三角瓶中,麸皮含水量65%,121℃高压灭菌20 min,然后按照深黄伞形霉∶黑曲霉(1∶1)将两种孢子混匀,孢子浓度分别按104CFU/g、105CFU/g、106CFU/g和107CFU/g接入,28℃培养6周,每周定时取样,取样烘干保留,测定油脂含量。
1.3.5 孢子比例对双菌培养油脂积累的影响
小麦麸皮为发酵原料,称取30 g于250 mL三角瓶中,麸皮含水量分65%,121℃高压灭菌20 min,然后按照深黄伞形霉:黑曲霉孢子比例2∶1、1∶1、1∶2将两种孢子液混合,接入孢子浓度为106CFU/g麸皮,28℃培养6周,每周定时取样,取样烘干保留,测定油脂含量。
1.3.6 不同孢龄对双菌积累油脂的影响
小麦麸皮为发酵原料,称取30 g于250 mL三角瓶中,麸皮含水量65%,121℃高压灭菌20 min,然后分别将孢龄为7 d、9 d和11 d孢子,按照深黄伞形霉:黑曲霉接菌1∶1比例将两种菌液混合,接入孢子浓度为106CFU/g麸皮,28℃培养6周,每周定时取样,取样烘干保留,测定油脂含量。
1.3.7 油脂含量的测定
取烘干的深黄伞形霉0.1 g,加入10 mL配制好的氯仿∶甲醇=2∶1,放入摇床180 r/min震荡16 h。加入2.5 mL去离子水并摇匀1 min,再4 500 r/min离心10 min,取下层溶液经过0.45μm滤膜过滤,70℃加热去除氯仿,得到油脂。油脂称重后计算菌体中油脂的含量。
1.3.8 脂肪酸组分的测定
取油脂0.10 g与10 mL氯仿∶甲醇∶浓硫酸(5∶4.25∶0.75)混合、摇匀,在90℃水浴90 min,加入2.5 mL去离子水,振荡1 min,4 500 r/m离心15 min。然后将上层与3 mL氯仿混合,振荡1 min,4 500 r/min离心15 min。将两次所得氯仿层混匀过滤用于GC-MS分析。色谱柱为Agilent 19091S-433HP-5MS毛细管色谱柱(30 m×250μm×0.25 μm)。升温程序为初始温度60℃,以15℃/min的速度升温到180℃,保持1 min,再以10℃/min升温到270℃,保持5 min。载气为氦气;流速为1 mL/min;压力56.75 kPa;进样量为1μL,分流比10∶1。质谱部分电子轰击离子源的电子能量70 eV;离子源温度230℃;四级杆温度150℃。图谱采集方式为全扫描方式,质量扫描范围m/z为20~550。
1.4 数据统计
本研究所有实验均进行3次平行实验,利用Microsoft Office 2016进行数据分析,Origin 9.1软件绘制所有数据图,利用IBM SPSS Statistics 25软件对数据进行统计学分析,并采用Duncan法进行多重比较。
2 结果
2.1 初始含水量对油脂产量的影响
在不同的初始水分含量培养条件下,不同初始水分含量的培养基油脂产量有着相对一致的趋势。如图2所示,随着培养时间的延长,油脂产量均呈现前4周下降,第4周到第6周略微上升的趋势。而在培养初期,不同的初始水分含量对固态发酵初期的影响较大,如初始水分含量67%和75%的油脂产量明显优于50%和80%,这是因为固态发酵初期,含水量过低、过高均不利于菌体的培养。当水分含量过高时,培养基基质颗粒之间结合较为紧密,通气量不足,黑曲霉无法获得足够的氧气进行生长。而当水分含量过低时,培养基基质过于干燥,并不利于菌丝的附着,因此不利于菌体生长。且随着培养时间的延长,水分含量不断降低,黑曲霉与深黄伞形霉的生长受到抑制,因此油脂产量逐渐下降。而在固态发酵末期此时黑曲霉利用自身的复合酶体系,将麸皮转化为深黄伞形霉可利用的碳水化合物,油脂含量逐渐上升,因此最适初始水分含量为75%。

图2 不同初始含水量对油脂产率影响
2.2 孢子数对油脂产量的影响
当黑曲霉与深黄伞形霉混合发酵时,虽然不同的孢子数对油脂产量具有不同的影响,但不同接种孢子量的培养基油脂产率有着相对一致的趋势,如图3所示,油脂产量随培养时间的延长呈现前4周整体下降,4~6周,油脂产量上升趋势。在混合固态发酵初期,第一周,此时接种的黑曲霉的孢子萌发,代谢旺盛,产出的酶较多,因此接菌量为106CFU/g和107CFU/g麸皮时,油脂产率较高,而随着培养时间的延长,低接菌量在固态发酵后期优势明显,发酵末期接菌量为104CFU/g麸皮时,油脂产率较其他接菌量第二、四、六周都要高,这是因为过多的接种量会导致培养后期所需的营养不足以支持其生长,因此最适的孢子接种量为107CFU/g麸皮。

图3 不同孢子数对油脂产率的影响
2.3 孢子比例对油脂产量的影响
不同的孢子比例对油脂产量具有不同的影响,当黑曲霉占比较高,深黄伞形霉占比较低时,此时黑曲霉水解生成的碳水化合物较多,但缺乏足够的深黄伞形霉将其转化为油脂,因此油脂产量较低;而当深黄伞形霉占比较高,黑曲霉占比较低时,此时水解生成的碳水化合物较少,深黄伞形霉可利用的碳水化合物较少,转化生成的油脂较少,因此油脂产量较低。如图4所示,黑曲霉和深黄伞形霉接入不同孢子比例时,油脂产量随时间的变化趋势基本一致。当黑曲霉和深黄伞形霉接菌比例为1∶1或者1∶2时,油脂产量随时间变化趋势更为相近,但随着培养时间的延长油脂产量逐渐下降,这是因为随着营养的消耗菌种竞争剩余营养,因此油脂产量逐渐下降。而当黑曲霉和深黄伞形霉接菌比例为2∶1时,0~2周油脂含量逐渐上升,在第二周达到峰值,随后2~6周油脂含量逐渐下降,且当黑曲霉和深黄伞形霉接菌比例为1∶2时,自第一周到第六周其油脂产量均优于其他两种接菌比例。因此,可知深黄伞形霉过量时的油脂产量比黑曲霉过量时更高,混合发酵过程中深黄伞形霉的比例对油脂产量的影响更为显著,因此最适的孢子比例为黑曲霉∶深黄伞形霉=1∶1。
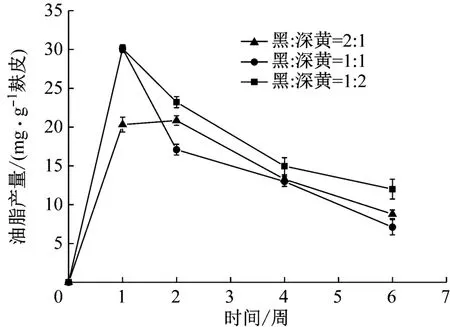
图4 不同孢子比例对油脂产率的影响
2.4 孢龄对油脂产量的影响
不同阶段的孢子对深黄伞形霉积累油脂具有不同的影响,不同孢龄的孢子由于其生长情况与自身的代谢程度不同,所以产出的酶量不同,因此可供深黄伞形霉利用的碳水化合物含量不同,油脂产量不同。如图5所示,不同的孢龄的孢子,油脂产量随时间变化趋势基本一致,但油脂产量各不相同。孢龄7 d的孢子,发酵第一周的油脂产量高于9 d和11 d的,为最大值,这是因为孢龄越小,其代谢愈加旺盛,产出的酶量也更多,所以油脂产量最高。而随着培养时间的延长,不同的孢龄的孢子油脂产量逐渐降低,其中孢龄11 d的油脂产量下降最快,这是因为其自身代谢较为迟缓,生长较慢,产出的酶量较少,油脂产量最低。因此选择孢龄7 d的孢子进行混合固态发酵最为合适。

图5 不同孢龄对油脂产率的影响
2.5 最适条件下的油脂产量
不同的影响因素对于固态发酵积累油脂具有不同的影响,如图6所示。实验结果表明,油脂含量在0 d~7 d不断升高,第7 d达到最高值,且随着培养时间的延长,油脂含量不断下降,因此固态发酵收获油脂的最佳时间为7 d。并且通过对初始水分含量、接种孢子数量、两种微生物孢子数比例和孢子的培养时间等因素进行优化可得,在最适培养条件下的油脂产量可达59.04 mg/(g麸皮),因此通过对固态发酵培养条件优化可以获得较高的油脂产量。
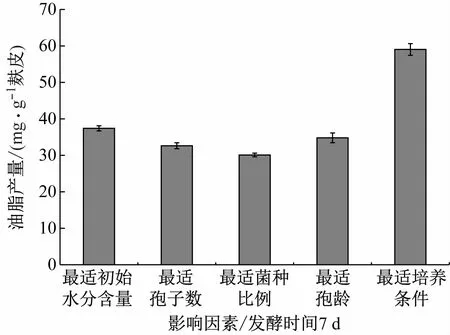
图6 不同影响因素对油脂产量的影响
2.6 脂肪酸成分分析
气相质谱联用仪能够对固态发酵得到的油脂成分进行定性和定量分析,按照实验方法对接菌量为107CFU/g麸皮,初始水分含量为75%,孢龄为7 d,两种菌比例1∶1的固态发酵所产油脂进行测定分析,结果见表1、表2和表3。
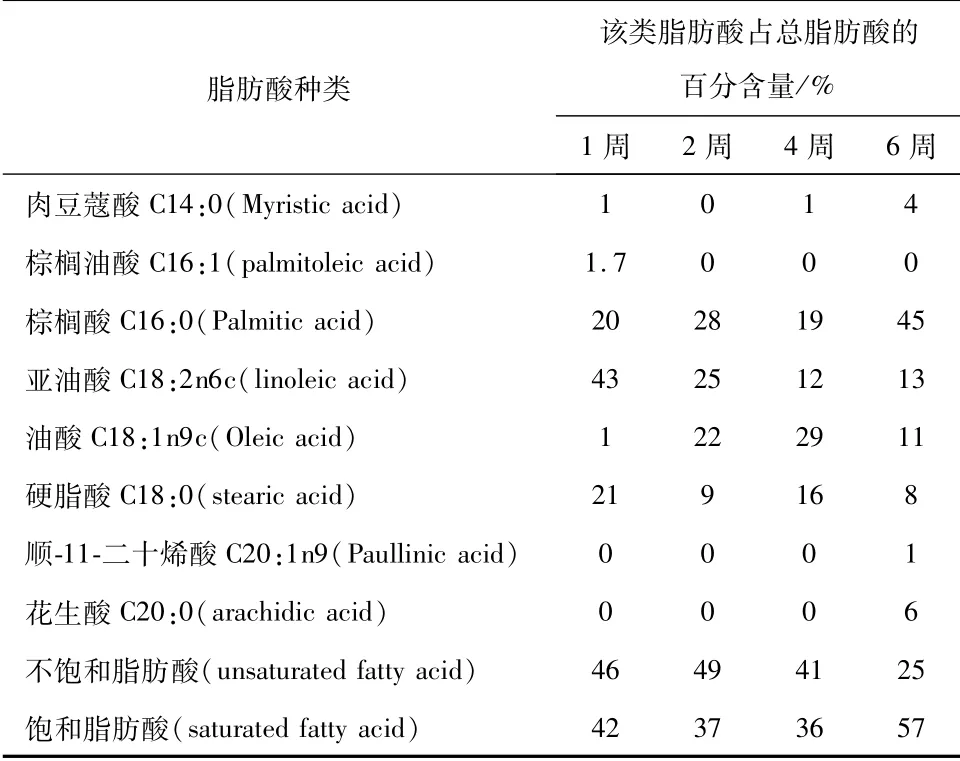
表2 微生物脂肪酸组成和含量实验分析结果
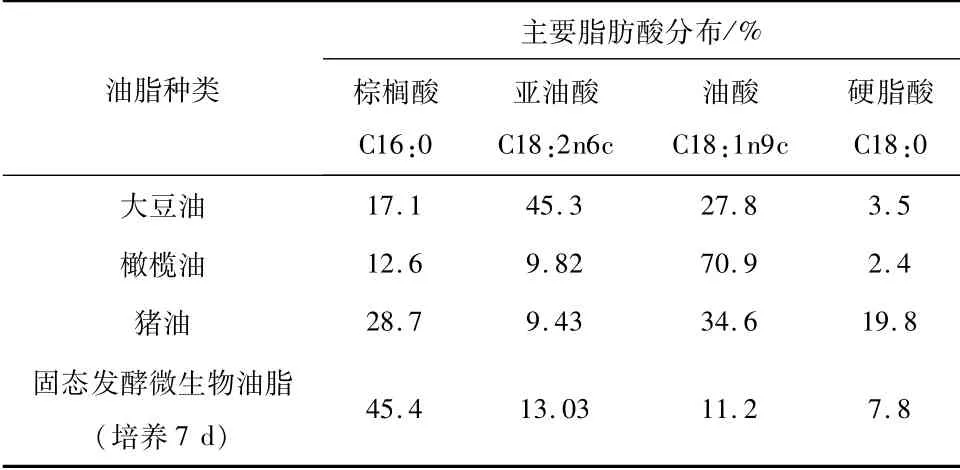
表3 四种油脂的主要脂肪酸分布表
表1中列出了通过气相质谱联用仪所测得的固态发酵油脂的主要脂肪酸成分,通过对深黄色孢霉积累的油脂成分分析可知,其脂肪酸主要成分与肉豆蔻酸、棕榈油酸、棕榈酸、亚油酸、油酸、硬脂酸、顺-11-二十烯酸、花生酸等皆有良好的相关度。其中棕榈油酸、亚油酸、油酸、顺-11-二十烯酸为不饱和脂肪酸,具有降低血脂、软化血管、降低血压降低心脏疾病、中风等风险的作用,并且在人和动物的新陈代谢过程中也起着重要作用。此外,不饱和脂肪酸还可用来制造肥皂、乳化剂、静电剂以及润滑柔软剂。而肉豆蔻酸、棕榈酸、硬脂酸、花生酸等为饱和脂肪酸,可用作硫化活性剂、润滑剂、脱模剂、稳定剂、表面活性剂以及塑料耐寒增塑剂,生产硬脂酸盐、橡胶工业、塑料、纺织等工业制品,因此通过深黄伞形霉与黑曲霉进行混合固态发酵积累油脂的途径是可行的。

表1 深黄伞形霉油脂成分分析
表2为微生物脂肪酸组成和含量随着培养时间的变化结果。由表可知,双菌固态发酵所产的微生物油脂的主要脂肪酸为棕榈酸、亚油酸、油酸和硬脂酸,且饱和脂肪酸含量随着培养时间的延长呈现先降低后升高的趋势;而不饱和脂肪酸则相反,呈现先升高后降低的趋势。在发酵初期,不饱和脂肪酸的含量较高,饱和脂肪酸的含量较低,随着培养时间的延长,不饱和脂肪酸的含量在0~2周逐渐升高,第2周达到最高值49%,随后2~6周逐渐下降,降至25%,而饱和脂肪酸的含量在0~4周先升高后逐渐降低,第4周达到最低值36%,随后4~6周逐渐升高达到最高值57%。其中不饱和脂肪酸的各个主要组分的变化也尽不相同,如棕榈油酸只在第1周存在,随后逐渐降至为0;顺-11-二十烯酸只在第6周存在,0~4周并不存在;亚油酸含量则在0~4周逐渐降低,4~6周短暂升高;油酸含量在0~4周逐渐升高,4~6周逐渐降低。而饱和脂肪酸中各个组分的变化也不同,如花生酸只在第1周存在;肉豆蔻酸含量为0~1周升高,1~2周降低,2~6周升高;棕榈酸含量为1~2周升高,2~4周降低,4~6周升高,硬脂酸含量则表现为1~2周降低,2~4周升高,4~6周降低,因此可知在固态发酵过程中,脂肪酸的成分并不是一成不变,随着培养时间的延长,不同的脂肪酸含量亦会发生不同的变化。由表2可知,表中的脂肪酸所占总脂肪酸的比例随着培养时间的延长呈现先下降后上升的变化。虽然培养后期表中脂肪酸所占比例增加至82%,但仍低于第1周的88%,且由图1~图4可知第1周的油脂含量最高,因此收获油脂的最佳时间为1周。
通过将深黄伞形霉与黑曲霉固态发酵积累的油脂与不同来源的油脂成分分析对比。由表2可知,双菌固态发酵所产的微生物油脂的主要脂肪酸为棕榈酸、亚油酸、油酸和硬脂酸。如表3所示,四种脂肪酸占总脂肪酸的77.43%,不饱和脂肪酸含量只占24%左右,低于动物油(猪油)和植物油(大豆油和橄榄油)。其中棕榈酸含量关系为固态发酵所得油脂>猪油>大豆油>橄榄油;亚油酸含量关系为大豆油>固态发酵所得油脂>橄榄油>猪油;油酸含量关系为橄榄油>猪油>大豆油>固态发酵所得油脂;硬脂酸含量关系为猪油>固态发酵所得油脂>大豆油>橄榄油;较其他三种油脂相比,固态发酵所得油脂油酸(11.2%)最低,硬脂酸(7.8%)含量低于猪油(19.8%)。与不同来源的油脂成分分析对比,双菌固态发酵所产的微生物油脂的主要脂肪酸为棕榈酸、亚油酸和油酸,这三种脂肪酸成分含量接近于大豆油。因此,通过深黄伞形霉与黑曲霉固态发酵所产的微生物油脂与植物油较为相近,与动物油相比存在较大差别。
3 结论与讨论
目前,大部分研究者致力于对深黄伞形霉的诱导突变以提高油脂的产量,诱导突变的常用方法有紫外照射、低能离子束、化学试剂诱导以及抗性筛选等方法。如刘胜男[22]通过氮离子反复诱变筛选获得一株遗传稳定的高产突变株F312,其γ-亚麻酸产量为1.2 g/L,比原始菌株(480 mg/L)提高了157.5%;而郝冉[23]通过紫外照射与抗性筛选,最终得到了一株具良好遗传稳定性的突变株,其GLA产量为889.8 mg/L,比原始菌株提高了60.3%。虽然通过诱导突变可以获得高产量的突变菌株,但此方法具有很强的不确定性,因此并不能用于提高油脂产量的实际生产中。此外,部分研究者还致力于对深黄伞形霉培养基的配方和培养条件优化以提高油脂产量。如张敏[24]首先对培养基中的葡萄糖、酵母膏和MgSO4进行单因素优化,并在此基础上进行响应面优化培养基的不同组分,得到了深黄伞形霉产培养基的最佳配方,在此条件下油脂产量为12.0 g/L,比优化前(6.2 g/L)提高了93.1%;SOMACAL[25]通过对酵母膏、蛋白胨和蔗糖三种不同培养基成分和三种发酵参数(搅拌速度、温度和pH)的优化分析,确定了250 mL摇瓶发酵的最佳生产条件,在此培养方式下,产脂率为31.77%,其中多不饱和脂肪酸含量为16.7%,而杨勇[26]通过响应面分析法优化深黄伞形霉突变菌株发酵制备花生四烯酸工艺,确定最优发酵工艺条件为:接种量15.7%,发酵温度28.29℃,发酵时间6.58 d,发酵pH 6.0,在此条件下花生四烯酸的产量为3.1 g/L。虽然优化培养基配方以及培养条件可以一定程度提高油脂产量,但效果并不明显,且此方法会增加成本,会造成资源的浪费,因此并不能应用于大规模的工业生产。深黄伞形霉虽然可以利用多种有机废弃物原料转化生成油脂,但是部分有机物在有机废弃物转化为油脂的过程中还会产生部分抑制物,如ŠANTEK[27]通过研究木质纤维素生物质水解过程中会产生的不同毒性抑制剂,毒性最大的是酚类,其次是呋喃甲醛和脂肪酸。酚类化合物对深黄伞形霉生长和脂质积累的抑制作用最强。而脂肪酸在低浓度时促进细胞生长和脂质积累,高浓度则强烈抑制这两个过程。因此当使用黑曲霉与深黄伞形霉进行混合固态发酵时可以很好地解决这些问题,因为黑曲霉可以分泌出较多的复合酶将食品废弃物水解成碳水化合物,不会生成抑制剂,不会对黑曲霉和深黄伞形霉生长造成影响。
由于双菌结合固态发酵的影响因素较多,如初始含水量、接菌量、接菌比例和接菌的孢龄等因素。因此通过对固态发酵的多个条件进行优化可得,固态发酵的最适初始水分含量为75%、最适接菌量为107CFU/g麸皮、最适孢龄为7 d、最适菌种比例为黑曲霉∶深黄伞形霉=1∶1,在此条件下可获得较高的油脂产量,为59.04 mg/(g麸皮)。其中所得油脂中的脂肪酸以棕榈酸(45.4%)、亚油酸(13.0%)、油酸(11.2%)和硬脂酸(7.8%)为主。虽然不同的发酵条件下油脂产量不同,但油脂产量几乎均呈现相同的降低趋势,随着培养时间的延长油脂含量不断下降,这可能是因为培养时间的延长,固态发酵过程面临菌种退化、营养原料不足以及代谢产物的抑制问题,因此为了提高油脂产量必须对这些问题予以解决。虽然采用基因工程手段提高深黄伞形霉纤维素酶类的产量也是一个技术上可行的措施,但由于废弃物的成分复杂需要很多种酶来协同作用,需要深黄伞形霉表达多种酶。这样就增加了技术难度,重组后深黄伞形霉的安全性也有待于重新评估。因此未来可以通过对黑曲霉菌株进行改造以及营养原料的改变以提高油脂产量,因为黑曲霉是食品安全性良好的工业菌株,已经广泛应用于淀粉加工、饮料、柠檬酸等食品物质的生产。通过对黑曲霉控制多种酶的基因进行改造,可以提高复合酶的产量,进而提高黑曲霉的水解麸皮的能力,提高黑曲霉与深黄伞形霉固态发酵的油脂产量。并且混合培养固态发酵不仅有利于丝状真菌的生长和酶的分泌,也有利于丝状真菌在基质上的附着和对基质的渗透,因此通过此方法提高油脂产量是可行的。而且还可通过添加不同的食品废弃原料以及改变不同的食品废弃物原料配比提高油脂产量,因为改变食品废弃物原料配比筛选出最适的食品废弃物原料配比体系,促进食品废弃物中糖的释放,进而提高食品废弃物的利用率,为深黄伞形霉积累油脂提高足够的原料。此外,转化过程中由于菌株代谢会产生部分抑制物抑制菌株的生长,可通过对转化过程中的抑制物去除以提升菌株的活力进而提高固态发酵过程中的油脂产量。因此可通过这些途径进一步提高油脂产量,为食品废弃物资源的转化以及可再生能源的研究提供参考。
猜你喜欢
农业工程学报(2022年1期)2022-03-25
河南畜牧兽医(2021年9期)2021-12-10
上海理工大学学报(2021年3期)2021-07-20
核化学与放射化学(2021年2期)2021-04-20
陶瓷学报(2021年1期)2021-04-13
陶瓷学报(2021年1期)2021-04-13
农村百事通(2018年13期)2018-09-13
作文周刊·小学四年级版(2017年21期)2017-07-10
中央民族大学学报(自然科学版)(2017年1期)2017-06-11
江苏农业科学(2016年1期)2017-05-17
