
冬小包脚菇的鉴定及其菌丝生长最佳营养条件研究
2022-06-27 03:52张贵合陶文广刘忠玄
种子 2022年5期
李 鹏, 熊 雪, 张贵合, 陶文广, 向 准, 王 晶, 刘忠玄, 黄 静
(1.贵州省生物研究所, 贵阳 550009; 2.贵州农业职业学院, 贵阳 551400)
冬小包脚菇(VolvariellabrumalisHe sp. nav.)是一种贵州特有野生可食用大型真菌,最早由贵州省生物研究所何绍昌先生于1987年正式命名并发表[1],俗称谷桩菌、低温草菇。该菌隶属于担子菌门(Basidiomycota),蘑菇纲(Agaricomycetes),蘑菇目(Agaricales),光柄菇科(Pluteaceae),包脚菇属(Volvariella)[1-2]。冬小包脚菇营养成分丰富,口感上佳,富含多种氨基酸及无机元素[3],其口感及外观和同属著名品种草菇(Volariellavolvacea)较为相似,但该菇子实体形成温度为4~14 ℃,与喜高温高湿环境的草菇有明显区别。草菇是包脚菇属最著名的食用菌,广受人们喜爱[4],其味道鲜美,富含粗纤维、氨基酸、维生素和特异性蛋白等[5-6],素有“中国蘑菇”之称,在食用菌市场上占有较为优势的地位。但草菇喜高温且子实体不耐低温储存的特性严重限制了其产业的发展[7-11]。而冬小包脚菇的发现填补了该属可食用种类均为中高温型的缺陷,对其种群理论、生物遗传学理论研究等方面具有重要的科学意义,且对弥补草菇冬季市场具有重要意义。但分离和培养难度大等原因,虽有许多学者对其开展研究,但目前冬小包脚菇除营养成分[3]、野外生境[12-13]、形态特征[14]等方面外,在其他基础研究、品种选育等方面仍处于空白状态。本实验按照基础生物学的方法对分离得到的冬小包脚菇菌株开展营养条件研究,旨在摸索菌丝生长规律,为冬小包脚菇优质母种的产业化、标准化生产提供理论依据,助推贵州特色冬小包脚菇产业规模化发展,填补我国草菇的冬季市场。

图1 野生冬小包脚菇子实体Fig.1 Wild Volvariella brumalis fruiting bodies
1 材料与方法
1.1 材 料
2019年12月于贵州省龙里县[1]采集而得的冬小包脚菇野生子实体,经组织分离培养后得到供试菌株,现保存于贵州省生物研究所。
活化培养基:马铃薯(去皮)200 g,稻草40 g(剪成2~3 cm段,沸水煮30 min),葡萄糖20 g,磷酸二氢钾0.5 g,磷酸氢二钾0.5 g,MgSO4·7 H2O 0.5 g,蛋白胨2 g,维生素B110 mg,琼脂粉20 g,去离子水1 000 mL,pH自然。
基础培养基:葡萄糖20 g,蛋白胨2 g,磷酸二氢钾0.5 g,磷酸氢二钾0.5 g,MgSO4·7 H2O 0.5 g,琼脂20 g,蒸馏水1 000 mL,pH自然。
1.2 试验方法
1.2.1菌种活化
制备冬小包脚菇活化培养基平板20个,无菌环境下将试管母种接种到冬小包脚菇加富PDA培养基上(接种至平板的中心位置),15 ℃恒温培养箱中培养20 d(待菌落生长至平板直径的1/2~2/3之间)。
1.2.2ITS序列分析
采取购买的生工生物工程(上海)股份有限公司Ezup柱式真菌基因组DNA抽提试剂盒提取分离纯化真菌基因组DNA,提取之后的DNA选用真菌通用引物(ITS 1:TCCGTAGGTGAACCTGCGG;ITS 4:TCCTCCGCTTATTGATATGC)进行PCR扩增。采用25 μL的PCR反应体系:Template(基因组20~50 ng/μL)0.5 μL;10×Buffer(和Mg2+);dNTP(各2.5 mmol/L)1 μL;Taq0.2 μL;F(10 μmol/L)0.5 μL;R(10 μmol/L)0.5 μL;加双蒸水至25 μL。反应程序为94 ℃预变性4 min,35个循环(94 ℃变性15 s、58 ℃退火15 s、72 ℃延伸1 min),72 ℃延伸5 min,4 ℃终止反应保存。扩增得到的PCR产物经1%琼脂糖电泳检测扩增结果后,送生工生物工程(上海)股份有限公司测序,测序并完成序列拼接后登陆GenBank上传所测序列,得到GenBank接收号,并通过BLAST对测序结果进行比对分析,下载最相近菌株的ITS rDNA序列,并构建系统发育树。
1.2.3温度筛选试验
将菌丝长至平板2/3的活化平板,用5 mm的打孔器将菌落打成圆片,然后接种于活化培养基上,将接种好的平板分别放置在10 ℃、15 ℃、18 ℃、20 ℃、22 ℃、25 ℃的恒温培养箱中培养,每处理重复5次,观察菌丝生长情况,同时记录菌丝萌发时间。菌丝生长速度测定采用“十”字交叉法,从菌丝萌发开始,每3天测量一次。菌丝铺满平板时,观察记录菌落的菌丝形态、色泽、菌落形状、菌丝长势和边缘特征等,用“+”“-”表菌丝的长势,“+”越多表菌丝生长得越好、越健壮、均匀,“-”表菌丝不生长。
1.2.4酸碱度筛选试验
以1 mol/L HCl和1 mol/L的NaOH,按不同比例配制基础培养基,分别设pH值为3、4、5、6、7、8、9、10、11、12共10个处理。将菌丝长至平板2/3的活化平板,用5 mm的打孔器将菌落打成圆片,然后接种于不同pH值平板的中心位置,每处理重复5次,置15 ℃恒温箱中培养,观察菌丝生长情况。选择菌落最大的条件作为最适pH值。
1.2.5碳源优化试验
以基础培养基为对照(ck),分别以等量蔗糖、麦芽糖、乳糖、果糖、甘露醇、可溶性淀粉、糊精及麦芽提取物代替基础培养基中的葡萄糖,作为不同碳源培养基,将菌丝长至平板2/3的活化平板,用5 mm的打孔器将菌落打成圆片,然后接种于不同碳源培养基上,每处理重复5次,置15 ℃恒温箱中培养,观察菌丝生长情况。
1.2.6氮源优化试验
以基础培养基为对照(ck),分别以等量的硝酸钾、硫酸铵、尿素、牛肉膏、豆粕粉、硝酸铵、酵母浸粉、酵母膏和麦麸分别代替基础培养基中的氮源蛋白胨,后续操作同1.2.5。
1.2.7不同碳氮比试验
在固定温度15 ℃和pH自然的培养基中调整碳氮比进行试验,基础培养基中碳源浓度为2%,添加不同重量蛋白胨(蛋白胨含碳量忽略不计),配成碳氮比为10∶1、20∶1、30∶1、40∶1、50∶1、60∶1、70∶1、80∶1、90∶1和100∶1的培养基,观察不同碳氮比对冬小包脚菇菌丝生长的影响。每个处理5次重复,置于15 ℃恒温培养箱中培养。
1.2.8最佳碳氮源浓度组合
选择最优的培养温度和pH值,选取的2个适宜碳源、氮源作为试验的四因素,采用 L9(34) 正交表进行四因素三水平正交试验,以不含碳源、氮源的基础培养基为基础,组合成供试培养基(表1)。每组5个重复,后续操作同1.2.5。
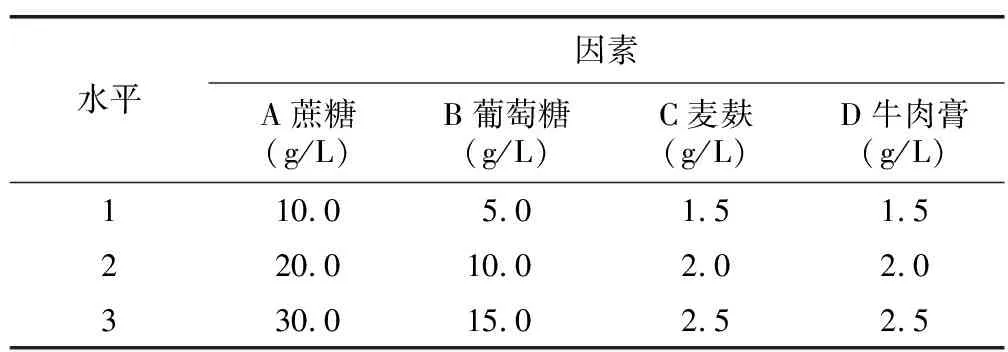
表1 正交试验因素水平Table 1 Table of factor level by orthogonal test
1.2.9最佳碳氮源浓度组合验证实验
经碳氮源正交试验得到该水平最优组合培养基,若组合不在实验处理中,则进行验证培养,操作同1.2.5。
2 结果与分析
2.1 DNA的提取及ITS序列分析
经PCR扩增和序列测定,结果显,供试冬小包脚菇菌株rDNA ITS区段长度为640 bp,上传GenBank得到登录号为OM 108495,经Blast搜索,击中结果均为Volvopluteusgloiocephalus和Volvariellagloiocephala,下载相似序列20条,并结合Volvopluteus、Volvariella及光柄菇科下光柄菇属(Pluteus)常见种序列,共同构建冬小包脚菇系统发育树(图2)。由图2可知,该系统发育关系若分为两大类群,则Volvopluteus、Volvariella在同一类群,Pluteus单独为一个类群。若分为三大类群,则Volvopluteus、Volvariella和Pluteus各为一个类群,且冬小包脚菇属于Volvopluteus类群,同Volvopluteusgloiocephalu亲缘性近;同著名草菇栽培种(Volvariellavolvacea)亲缘性较远,存在本源差异。

图2 基于ITS序列构建的冬小包脚菇系统发育树 Fig.2 Phylogenetic tree of Volvariella brumalis constructed based on ITS sequence.
2.2 不同温度对冬小包脚菇菌丝生长的影响
如表2所,在所选温度范围内,冬小包脚菇菌丝萌发、生长速度和菌落长势均随温度变化差异明显。在低温(5 ℃)、高温(30 ℃)条件下冬小包脚菇菌丝均不能正常萌发生长,10 ℃培养下菌丝萌发需耗时16 d,其余温度条件下仅需2 d,说明温度过高或过低均会对冬小包脚菇菌丝萌发产生明显抑制作用。在10~15 ℃条件下,冬小包脚菇菌落均呈绒毛状白色圆形菌落,有气生菌丝,发达程度不一,随温度升高菌落长势整体呈现“弱—旺—弱”的趋势,其中15 ℃条件下培养菌落长势最好,18 ℃和20 ℃培养下次之。菌丝日均生长速度也随温度升高先增后减,18 ℃培养条件下菌丝日均生长速度最快,达1.03 mm/d,15 ℃和20 ℃培养条件下次之,分别为0.95 mm/d和0.90 mm/d。综上说明冬小包脚菇菌丝生长适宜温度条件范围在15~20 ℃之间,最佳适宜温度为15 ℃。

表2 不同温度对冬小包脚菇菌丝生长的影响Table 2 Effects of different temperatures on Volvariella brumalis mycelium
2.3 不同酸碱度对冬小包脚菇菌丝生长的影响
如表3所,冬小包脚菇菌丝萌发和生长受培养基酸碱度影响明显,该菌在过酸和过碱条件下均不能正常萌发和生长。pH<5或pH>10时,冬小包脚菇菌丝均不能萌发生长,其菌丝萌发适宜生长酸碱度范围在6~9之间,且菌丝萌发后随着酸碱度的增加,菌丝日均生长速度呈先增后减的抛物线型变化。在pH=8条件下菌丝生长最快,日均生长速度达3.75 mm/d,其次为pH=9和pH=7条件,日均生长速度分别为2.74 mm/d和2.26 mm/d,其菌落长势表现同生长速度趋势一致。综上可知,冬小包脚菇菌丝对酸碱度变化反应敏感,过酸过碱均会抑制其萌发和生长,其菌丝生长喜弱碱性环境,适宜酸碱度范围为7~9,其中8为最佳。

表3 不同酸碱度对冬小包脚菇菌丝生长的影响Table 3 Effects of different pH conditions on Volvariella brumalis mycelium
2.4 不同碳源培养基对冬小包脚菇菌丝生长的影响
如表4所,在9种供试碳源培养基条件下,冬小包脚菇菌丝萌发时间均为2 d,且均呈现出绒毛状灰白色圆形菌落,有气生菌丝着生,但表现差异较大,菌落长势差异明显。其中在蔗糖培养基中长势最佳,乳糖、糊精和麦芽提取物培养下菌落长势最差。各种培养基条件下的冬小包脚菇菌丝日均生长速度在p<0.05水平上有差异。其中碳源为甘露醇培养条件下菌丝显著快于其他培养基,日均生长速度为2.59 mm/d。其次为蔗糖和葡萄糖,日均生长速度分别为2.27 mm/d和2.06 mm/d。结合菌落长势可知,冬小包脚菇最适碳源为蔗糖,其次为甘露醇和葡萄糖。

表4 不同碳源培养基对冬小包脚菇菌丝生长的影响Table 4 Effects of different carbon source media on the growth of Volvariella brumalis mycelium
2.5 不同氮源培养基对冬小包脚菇菌丝生长的影响
由表5可知,冬小包脚菇在10种供试氮源培养基上均能萌发生长,且表现为着生有一定量气生菌丝的绒毛状灰白色圆形菌落,但各处理下菌落长势和日均生长速度差异明显。其中氮源为尿素、牛肉膏和麦麸培养下长势较佳,酵母膏和酵母浸粉次之,氮源为硫酸铵和硝酸铵培养时,菌丝长势最差,说明有机氮较无机氮更有利于冬小包脚菇菌丝生长。冬小包脚菇在麦麸培养基上菌丝生长速度显著快于其他培养基(p<0.05),日均生长速度达2.65 mm/d。其次为牛肉膏、硝酸铵和尿素,其日均生长速度分别为2.24 mm/d、2.20 mm/d和2.01 mm/d。因此,结合菌落长势可知,冬小包脚菇菌丝生长最佳氮源为麦麸,其次为牛肉膏和尿素。

表5 不同氮源培养基对冬小包脚菇菌丝生长的影响Table 5 Effects of different nitrogen source media on the growth of Volvariella brumalis mycelium
2.6 不同碳氮比培养基对冬小包脚菇菌丝生长的影响
如表6可知,在不同碳氮比实验中,冬小包脚菇菌丝萌发情况不受碳氮比的影响,且菌落长势差距不大,均表现为绒毛状白色圆形菌落,并着生有一定量气生菌丝,但在碳氮比为20∶1、30∶1和40∶1的条件下时,菌落气生菌丝明显较其他碳氮比多且发达,整体长势较佳。结合图3可知,冬小包脚菇菌丝日均生长速度整体差距不大,其中碳氮比为70∶1时,生长速度最快,达1.84 mm/d,碳氮比为50∶1时,生长速度最慢,为1.35 mm/d。综合菌落长势可知,冬小包脚菇日均生长速度适宜碳氮比在20∶1~40∶1之间,最佳碳氮比为30∶1。
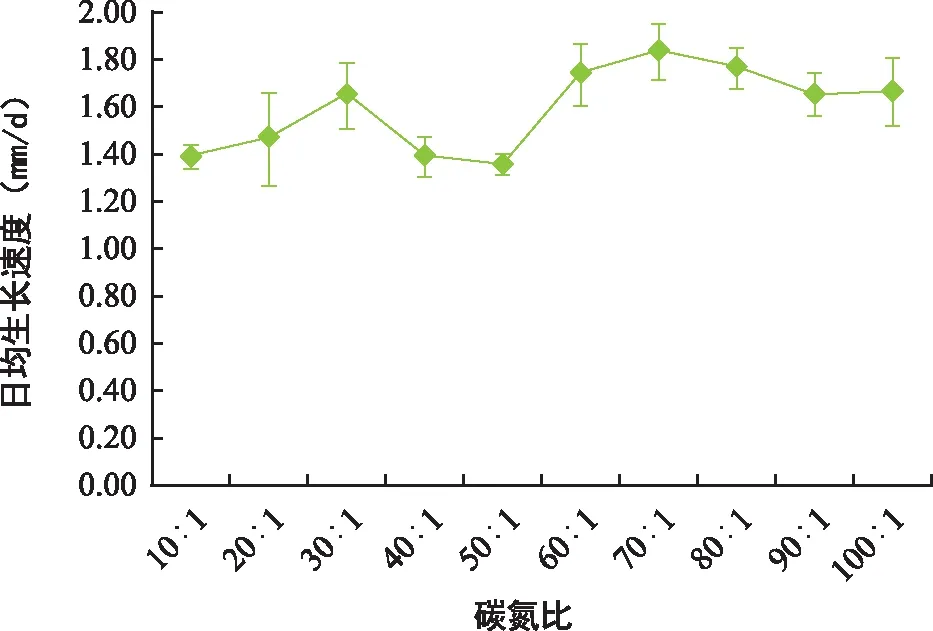
图3 不同碳氮比培养基对冬小包脚菇菌丝生长速度的影响Fig.3 The effect of different C/N ratio media on the growth of Volvariella brumalis mycelium

表6 不同碳氮比培养基对冬小包脚菇菌丝生长的影响Table 6 Effects of different C/N ratio media on the growth of Volvariella brumalis mycelium
2.7 最佳碳氮源浓度组合
以2.4、2.5中所得冬小包脚菇适宜碳源、氮源为依据,选择适宜碳源蔗糖、葡萄糖和适宜氮源麦麸、牛肉膏建立 L9(34) 正交表进行四因素三水平正交试验(表7),各处理培养下冬小包脚菇菌株萌发时间无差异,且均为绒毛状整齐灰白色圆形菌落,并着生大量气生菌丝,菌丝健壮、发达且浓密度高。其中V 2处理下冬小包脚菇菌丝显著高于其他处理,日均生长速度为4.28 mm/d,其次为V 1和V 3,日均生长速度分别为3.52 mm/d和2.96 mm/d。综上可知,在所有处理中,冬小包脚菇菌丝生长最佳碳氮源组合为V 2(A1B2C2D2),日均生长速度达4.28 mm/d。

表7 不同浓度碳氮源对冬小包脚菇菌丝体生长的影响Table 7 Effects of different concentrations of carbon and nitrogen sources on the growth of Volvariella brumalis mycelium
由表8可知,4个因素R值从大到小为RA、RC、RD、RB,表明各因素对冬小包脚菇菌丝生长速度的影响为A(蔗糖)>C(麦麸)>D(牛肉膏)>B(葡萄糖)。此外,在4种影响因素中,因素A对菌丝生长速度有极显著影响,因素C对菌丝生长速度影响显著(表9),因素B、D影响不显著,但因素D影响高于因素B(pD

表8 碳氮源对冬小包脚菇菌丝体生长的正交分析Table 8 Orthogonal analysis of carbon and nitrogen sources on the growth of Volvariella brumalis mycelium
说明碳源蔗糖和氮源麦麸是冬小包脚菇菌丝培养阶段的关键因素,其中主要因素为碳源蔗糖,次要因素为氮源麦麸。
按照各因素的优水平,得到A1B1C2D2水平组合为本试验的最优水平组合,结合表1可知,该组合为蔗糖10.0 g,葡萄糖5.0 g,麦麸2.0 g,牛肉膏2.0 g。但该组合并不在试验处理中,通过后续试验验证得到冬小包脚菇日均生长速度为4.35 mm/d,比上述正交试验处理所得最佳组合V 2(A1B2C2D2)高0.07 mm/d。

表9 冬小包脚菇正交试验方差分析Table 9 Variance analysis of Volvariella brumalis by orthogonal test
3 讨 论
冬小包脚菇是一种独具特色的珍稀食用菌,在外形、口感上同著名栽培种草菇较为相似,但草菇出菇嗜高温高湿,子实体贮存湿度一般不低于15 ℃,且储存时间非常短,而冬小包脚菇偏好低温出菇,子实体可耐低温保存,这样的先天条件决定了冬小包脚菇重要的研究及市场价值。本研究对分离得到的冬小包脚菇菌丝体进行了分子鉴定,初步研究了其系统发育关系,发现冬小包脚菇属于Volvopluteus类群,且同Volvopluteusgloiocephalus、Volvariellagloiocephala亲缘性最近;同著名草菇栽培种(Volvariellavolvacea)亲缘性较远,存在本源差异。这同何绍昌等[1]将冬小包脚菇命名在Volvariella类群下并不冲突,因为Volvopluteus是2011年由Justo等从传统Volvariella类群中分出来的新类群,而冬小包脚菇早在1987年已被命名。而同一分支下Volvopluteusgloiocephalus、Volvariellagloiocephala也被Justo等[15]归属在Volvopluteus类群,并统一名称为Volvopluteusgloiocephalus[17]。冬小包脚菇同Volvopluteusgloiocephalus虽具有较近的亲缘关系,但具体的种间关系需要进一步的研究。
冬小包脚菇菌丝体基础生物学特性研究发现,冬小包脚菇菌丝生长适宜温度范围在15~20 ℃之间,最佳生长温度为15 ℃,且当环境温度低于5 ℃或高于25 ℃时无法萌发,但菌丝体不会死亡。这同草菇适宜35 ℃环境,且在低于20 ℃或高于40 ℃环境下不生长或生长不良的习性具有较大差异[18]。此外冬小包脚菇菌丝体对环境酸碱度反应敏感,且偏好弱碱性环境,在pH<5和pH>10的环境下,菌丝体均无法正常萌发生长,适宜酸碱度范围在7~9之间,最适为8。这同草菇对酸碱度的反应表现较为一致,但草菇对酸碱度的适应能力更强于冬小包脚菇。历年来对草菇的持续研究中,均发现草菇对酸碱度的适宜范围较广,在pH=4~11范围内均能生长,更偏爱碱性环境[18-21]。而在适宜碳氮源选择上,冬小包脚菇最佳碳源为蔗糖,其次为甘露醇和葡萄糖。最佳氮源为麦麸,其次为牛肉膏和尿素,且对冬小包脚菇进行四因素三水平正交试验中发现,影响冬小包脚菇菌丝生长占主要因素的是碳源蔗糖。在荆蓉和郭成金[19]对银丝草菇培养条件研究中也发现,银丝草菇对碳源蔗糖和葡萄糖有较好的吸收能力,而对氮源牛肉膏和麦麸的吸收力度则没有酵母粉和蛋白胨高。伍国明和李梅[20]对草菇不同菌株研究中发现,草菇对碳源蔗糖的吸收更强,且氮源表现不一,但整体而言同银丝草菇一样,草菇对酵母粉、蛋白胨等复杂氮源均能较好地吸收[20-22]。这说明尽管冬小包脚菇和草菇菌丝适宜生长温度范围差异较大,但相近的酸碱度和碳源选择,也是冬小包脚菇野生资源开发的一个突破口,这对冬小包脚菇栽培基质配方的摸索具有指导意义。试验最终得到冬小包脚菇菌丝生长最优培养条件为蔗糖10.0 g,葡萄糖5.0 g,麦麸2.0 g,牛肉膏2.0 g,KH2PO41 g,MgSO4·7 H2O 0.5 g,琼脂20 g,蒸馏水1 000 mL,酸碱度调至8,接种后置于15 ℃条件下培养。但尽管在最适宜的条件下,冬小包脚菇在平板上的平均生长速度仅保持在4.35 mm/d,相对草菇培养生长速度(5.10 cm/d)慢了10倍以上[19]。母种生长缓慢是冬小包脚菇培养及产业化发展的难关,这将是下一步研究的重点之一。
猜你喜欢
今日农业(2022年3期)2022-11-16
油气田地面工程(2022年9期)2022-10-26
中国农业科技导报(2022年7期)2022-09-27
江苏农业科学(2022年6期)2022-04-15
铀矿冶(2022年1期)2022-02-17
饮食与健康·下旬刊(2019年3期)2019-03-08
饮食与健康·下旬刊(2018年3期)2018-04-11
农家顾问(2016年12期)2017-01-06
世界热带农业信息(2016年1期)2016-04-13
分析化学(2014年10期)2014-10-24
