
准噶尔山楂种子内源抑制物质活性研究
2022-06-27 03:52:50如马南木尼合买提聂必林热依汉古丽夏迪巫利梅吕海英
种子 2022年5期
如马南木·尼合买提, 聂必林, 热依汉古丽·夏迪, 巫利梅, 吕海英
(1.旱区植物逆境生物学实验室, 新疆 乌鲁木齐 830054;2.新疆特殊环境物种保护与调控生物学实验室, 乌鲁木齐 830054;3.新疆师范大学生命科学学院, 乌鲁木齐 830054)
种子休眠是植物具有的一种复杂的机制,是种子萌发和植物生长发育的重要保障[1-3]。种子休眠不仅在高等植物个体发育中起重要作用,也和种群的生存及发展息息相关[4-5]。对于植物而言,种子休眠是一种有利的生态适应特征,它会推迟种子的发芽,直到季节和生长条件对幼苗的生长有利时才会打破休眠[6-8]。但是对种子生产、人工扩散和栽培带来许多不便,影响该植物种群的更新速度[9-11]。内源抑制物质是种子休眠的主要原因之一[12],种子的不同部位可能存在不同的内源抑制物质,这些抑制物通过不同途径阻碍种子生理活动或某个代谢环节,从而降低种子的发芽率[13-15]。种子通过分解这些化学物质来解除抑制作用进而萌发[16-17]。随着国内外对内源抑制物研究的深入,各式各样的植物内源抑制物不断被发现。朱铭玮等[18]对油用牡丹‘凤丹’种子内源抑制物研究发现,‘凤丹’种皮和种胚中含有抑制作用较强的内源抑制物,不同部位不同提取相对白菜种子发芽的影响不同。邹雨婷等[19]对绿山楂内源抑制物质成分的分析表明,绿山楂种子浸提液中的乙醚相和甲醇相对白菜种子发芽的抑制作用最强,其内源抑制物以酚类、酯类、有机酸类等为主。由此可见,不同种类植物,抑制物质在种子内的存在部位以及所含抑制物类型不同,其解除方法也不同[20-21]。因此,通过研究植物种子中内源抑制物质的类型、功能、活性和作用机理,打破种子休眠、促进种子萌发,不仅是解决植物人工繁殖和扩大资源问题的关键技术[22],而且在农业和林业生产中也具有重要意义[23]。
准噶尔山楂(Crataegussongarica)为蔷薇科山楂属光核组唯一一种植物,也是分布于新疆伊犁部分山区的野山楂资源之一[24],具有很强的抗旱和抗寒特性,可作为观赏树种以及栽培山楂的砧木及育种材料,利用价值高[25]。同时准噶尔山楂也是珍稀的天然花色苷类色素资源之一[26],而且作为天山野果林的组成树种之一,是天山水源涵养林的一部分[27],对防止水土流失、改善生态环境、维持其原生性具有重要现实意义[28]。野外调查发现,在自然环境中准噶尔山楂幼苗数量稀少,海拔在1 100~1 600 m范围内,实生苗仅为0.001 9株/m2。张侠等[29]研究表明,准噶尔山楂种子具有深度休眠特性,导致出苗率低。此外,近年来由于环境变化和人为因素的影响,已导致准噶尔山楂种群的栖息地自然资源萎缩、病害严重、群落退化和严重破坏[30]。因此,在前人研究的基础上,本研究以准噶尔山楂种子为材料,通过系统溶剂法分析准噶尔山楂种子不同部位的浸提液对白菜种子的化感作用,明确其内源抑制物的活性及分布,旨在为准噶尔山楂种子休眠破除技术及休眠机理的深入研究提供参考,为该类群植物的保护和利用提供科学依据。
1 材料与方法
1.1 材 料
供试的准噶尔山楂采自新疆霍城县大西沟,剥去果肉,洗净,阴干后-80 ℃冰箱保存备用。实验所用白菜品种为四季小白菜,种子纯度95%,净度98%,发芽率85%,含水量8%。
1.2 研究方法
1.2.1千粒重测定
随机抽取100粒山楂籽,用千分之一电子天平称量并记录,重复8次,计算其标准偏差和千粒重。
1.2.2准噶尔山楂种子不同部位甲醇浸提液的制备
对试验的山楂种子进行解剖,并将其分为三部分(种壳、种皮和种胚)。每一部分用破碎机破碎,并在33~35 ℃下干燥至恒重。取准噶尔山楂不同部位粉末40 g,分别置于250 mL锥形瓶中,加入5倍体积的80%甲醇,混匀,置于4 ℃冰箱中恒温提取。期间多次取出摇匀,充分提取,24 h后过滤,重复提取3次,将所得提取液混合,减压浓缩后定容至250 mL。
1.2.3准噶尔山楂种子不同部位内源抑制物质的初步分离
根据图1所的过程,通过系统溶剂法[31]分离种子各部分的提取物。分离出的石油醚相、乙醚相、乙酸乙酯相、甲醇相和水相提取物用旋转蒸发器蒸发和浓缩。当定容达到250 mL后,将其置于4 ℃冰箱中保存备用。
1.2.4准噶尔山楂种子不同提取相的活性测定
以白菜种子为材料,测定上述提取相的活性:白菜种子在45 ℃水浴中提前浸泡10 min,并在此期间持续搅拌。在12 cm培养皿中放置一层滤纸,分别加入各相提取液3 mL,取50粒白菜种子均匀放置于滤纸中,湿度保持在85%以上,在(23±2)℃恒温暗培养,并加入与对照组(ck)相同体积的蒸馏水,每组重复4次。48 h后,计算白菜种子的发芽率(发芽标准为幼根伸长长度大于种子长度)。72 h后,测定幼苗高度和根长、干重和鲜重,并检验差异显著性。根据实验数据,进一步计算种子的发芽率(GP)、发芽指数(GI)、活力指数(VI)、平均发芽时间(AGT)和相对发芽率(RGP)。各指标的计算公式如下:
GP(%)=(Ni/N)×100%,Ni为第i天发芽数;N为种子总数;
RGP(%)=(处理发芽率/对照发芽率)×100%;
抑制活性=(1-处理组/对照组)×100%;
GI=∑(Gt/Dt),式中Gt为第t日发芽种子数,Dt为对应Gt的发芽天数;
VI=S×GI,式中S为实验结束时胚根平均鲜重(g);
AGT=∑(dn)/∑n,式中d表从播种之日起到种子萌发所用天数,n表对应各日新增发芽种子数。
1.3 数据分析
所得数据用Excel 2017软件进行处理和绘图,并用SPSS 17.0软件进行差异显著性分析和多重比较。实验结果以平均值±标准误差表。

图1 准噶尔山楂种子内源物质的初步分离流程Fig.1 Preliminary separation process of endogenous substances in C.songarica seeds
2 结果与分析
2.1 种子千粒重的测定结果与分析
种子的千粒重为确定种子的大小和饱满度提供重要的参考值,种子的质量也可以通过千粒重来测试[32]。本试验中,准噶尔山楂种子的千粒重为112.769 g,标准偏差为0.001 85,变异系数为14.25%。变异系数较高,这可能是准噶尔山楂种子采集于不同海拔高度所致。
2.2 甲醇浸提液各提取相对白菜种子发芽率的影响
由图2可见,准噶尔山楂种子不同部位的浸提液对白菜种子发芽率的抑制效果不同。种壳的乙酸乙酯相对白菜种子发芽率具有明显的抑制作用,并与ck以及其他提取相均差异极显著(p<0.01),处理后的白菜种子几乎不能发芽,发芽率与ck相比降低了95%,抑制活性达到了96.94%,且显著高于种皮和种胚的同相(p<0.01)。种壳的其他提取相以及种皮、种胚的各提取相的发芽率与ck差异均不显著(p>0.05)。由此可以看出,影响白菜种子发芽的抑制物质主要存在于种壳的乙酸乙酯相中。

注:不同大写字母表同一部位提取不同相间差异显著(p<0.01);不同小写字母表同一部位不同提取相间差异显著(p<0.05)。图2 甲醇浸提液各提取相对白菜种子发芽率的影响Fig.2 Effects of methanol extracts on germination rate of cabbage seeds
2.3 甲醇浸提液各提取相对白菜种子GI和VI的影响
种子的GI、VI可以用来衡量该种植物在一定条件下的发芽能力和活力,能够很好地反映环境对种子发芽的影响程度[33]。由图3可知,除了种胚的水相和种皮的石油醚相外,准噶尔山楂种子各部位不同提取相对白菜种子发芽指数都有显著的抑制作用(p<0.05),但准噶尔山楂种子部位不同、不同提取相,其对白菜种子发芽指数的抑制程度也有所不同。显著性分析结果显,种壳各提取相对白菜种子发芽指数的影响显著高于种皮和种胚(p<0.05);除了水相外,种皮和种胚各相的发芽指数差异不明显(p>0.05)。其中,准噶尔山楂种壳中的乙酸乙酯相对白菜种子发芽指数的影响最大,与ck以及其他各相的差异极显著(p<0.01)。

图3 甲醇浸提液各提取相对白菜种子GI的影响Fig.3 Effects of methanol extracts on germination index (GI) of cabbage seeds
由图4可知:除了种壳的水相外,准噶尔山楂种子各部位不同提取相对白菜种子活力指数都有显著的抑制作用(p<0.05),且有机相抑制作用均显著大于水相(p<0.05)。其中,准噶尔山楂种壳乙酸乙酯相的抑制作用最大(p<0.01),活力指数从0.74降至0.03。说明影响白菜种子发芽指数和活力指数的抑制物存在于准噶尔山楂种子各部位。

图4 甲醇浸提液各提取相对白菜种子VI的影响Fig.4 Effects of methanol extracts on vigor index (VI) of cabbage seeds
2.4 甲醇浸提液各提取相对白菜种子平均发芽时间的影响
种子的发芽速度对育苗具有重要的参考价值,其值越小说明种子发芽能力越强,是种子萌发过程中极其重要的指标[34]。从表1可见,准噶尔山楂种皮和种胚各提取相对白菜种子平均发芽时间的影响较小,而种壳反之,其中种壳乙酸乙酯相的影响最大,与ck及同组其他各相的差异极显著(p<0.01),与ck比平均发芽时间从1.15 d延长至2.86 d。除种壳的乙酸乙酯相外,白菜种子平均发芽时间集中在1.10~1.49 d之间。以上结果表明,影响白菜种子平均发芽时间的内源性抑制物主要集中在种壳浸提液中。

1 甲醇浸提液各提取相对白菜种子平均发芽时间的影响Table 1 Effects of methanol extracts of different phases on average germination time of cabbage seeds
2.5 甲醇浸提液各提取相对白菜幼苗生长的影响
从图5可知,准噶尔山楂种壳的乙酸乙酯相和水相以及种皮的水相对白菜幼苗根长的抑制作用不明显,种胚的水相和石油醚相有促进作用,其他相都有不同程度的抑制作用。其中,种壳的乙醚相、甲醇相和种皮的石油醚相、乙酸乙酯相、甲醇相以及种胚的乙醚相、甲醇相对白菜幼苗根长的抑制作用极显著(p<0.01)。
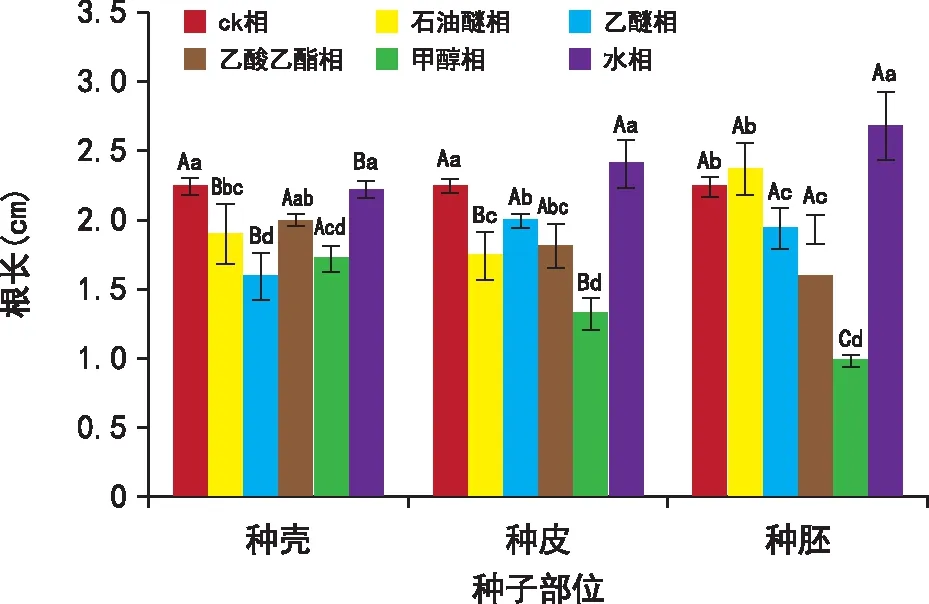
图5 甲醇浸提液各提取相对白菜幼苗根长的影响Fig.5 Effects of methanol extracts on root length of cabbage
由图6可知,在种壳中,除了水相和乙酸乙酯相外,乙醚相、石油醚相和甲醇相对白菜苗高均有显著的抑制作用(p<0.01),抑制作用表现为乙醚相>石油醚相>甲醇相。种皮和种胚提取相中除了水相外,其余各相对白菜苗高有极显著的抑制作用(p<0.01),抑制作用表现为:乙醚相>石油醚相>乙酸乙酯相>甲醇相。以上结果表明,准噶尔山楂种壳、种皮和种胚浸提液都对白菜苗高有不同程度的抑制作用。

图6 甲醇浸提液各提取相对白菜苗高的影响Fig.6 Effects of methanol extracts on seedling height of cabbage
3 讨论与结论
大多数休眠性种子的不同部位都存在内源抑制物[35],种类繁多,影响种子正常萌发和幼苗生长[36]。通常提取其成分并用非休眠的白菜种子做生物测定,以确定这些物质是否存在[37]。本研究从内源抑制物质的角度分析准噶尔山楂种子休眠的原因,结果表明,准噶尔山楂种壳、种皮及种胚不同提取相对白菜种子萌发、苗高和根长均有抑制作用,而且不同部位浸提液不同有机相的抑制作用差异很明显,有时还呈现促进生长作用。
杨晓玲等[37]通过比较4种山楂种子抑制物质对白菜种子萌发的影响,发现4种山楂种子各部位的乙醚相、乙酸乙酯相及甲醇相对白菜种子萌发均有较强的抑制作用,其中内果皮的抑制活性最强,其次是种皮和种胚;廖源林等[11]通过研究野鸦椿种壳和种胚的内源抑制物,发现种壳的抑制作用小于种胚,白菜种子发芽率随着种壳和种胚浸提液浓度的增加而持续降低,但幼苗的根长和苗高表现为低促高抑;史锋厚等[38]发现,南京紫椴种皮的抑制活性大于种皮和胚乳,种皮中含有的抑制物质主要存在于乙醚相和甲醇相,而种子中的抑制物质主要存在于甲醇相。本研究结果表明,准噶尔山楂种子不同部位均含有抑制剂,不同部位内源抑制剂的抑制程度不同:准噶尔山楂种子中活性较强的内源抑制剂主要分布在乙酸乙酯相、乙醚相和甲醇相,可能与它们的生理休眠密切相关,这与上述结果相似。而准噶尔山楂种子同一部位的同一提取物对白菜种子萌发和幼苗生长的抑制作用不同。种皮对白菜种子发芽率、发芽指数、活力指数和平均发芽时间的抑制作用大于种皮和种胚、种皮和种胚对白菜幼苗生长有较强的抑制作用。结果表明,准噶尔山楂种子中不仅存在一种抑制剂,而且这些抑制剂对种子萌发和幼苗生长有不同的抑制作用。种壳对白菜种子发芽率、发芽指数、活力指数及平均发芽时间的抑制作用大于种皮和种胚,而种皮和种胚对白菜苗生长的抑制作用较强。说明准噶尔山楂种子中所含抑制物并非只有一种,且这些抑制物对种子萌发和幼苗生长的抑制作用也不同。
另外,本研究发现准噶尔山楂种子不同部位的浸提液有机相对白菜种子和幼苗的抑制作用较大,而水相的抑制作用较小,种壳和种皮的水相以及种胚的水相和石油醚相等提取相有时对白菜种子和幼苗的生长具有促进作用。原因可能是准噶尔山楂种子甲醇浸提液的内源性抑制物主要存在于有机相,而且不同部位所含有的内源抑制物的种类或浓度不同,有些物质在适宜的浓度时可能会出现促进生长的作用。这与蒋玉婷等[10]、刘艳梅等[39]分别通过研究珍稀濒危种斧形沙芥和桔梗种子内源抑制物得到的结果是一致的。
内源抑制物能不同程度地对种子萌发起到抑制作用[40-41],不同内源抑制物抑制作用及其解除方法也不同[42-43]。本研究以及以往的研究结果表明,准噶尔山楂种子之所以在自然状态下萌发率低,不仅是因为外界环境等因素,而且种子本身也含有内源性抑制物。准噶尔山楂种子不同部位的浸提液中可能含有不同的抑制性化学成分,这些成分能抑制种子萌发及幼苗生长,然而不同部位抑制物的抑制强度不同。因此,要确定准噶尔山楂种子内源抑制物质的成分、种类及含量,还需要进一步深入研究。
猜你喜欢
粮油与饲料科技(2022年2期)2022-11-24 13:12:07
卫拉特研究(2021年0期)2022-01-19 02:32:16
卫拉特研究(2021年0期)2022-01-19 02:32:00
蒙古学问题与争论(2019年0期)2019-03-29 05:46:38
福建中医药(2017年6期)2018-01-09 06:49:54
卫拉特研究(2017年0期)2017-12-07 00:35:44
上海农业学报(2016年2期)2016-10-27 00:49:48
西南农业学报(2016年4期)2016-05-17 05:42:14
华东理工大学学报(自然科学版)(2015年4期)2015-12-01 04:00:23
动物营养学报(2015年10期)2015-12-01 02:26:21
