
施肥对黄栀子幼苗生长与光合特性的影响1)
2022-06-24 08:13:44尹梦雅李志辉杨艳李昌珠汤玉喜唐洁秦平书吴兴华
东北林业大学学报 2022年5期
尹梦雅 李志辉 杨艳 李昌珠 汤玉喜 唐洁 秦平书 吴兴华
(中南林业科技大学,长沙,410004)(湖南省林业科学院)(南华国家森林管理处)(凤凰县林业局)
黄栀子(Gardeniajasminoides)别名山栀子、红果树、林兰,是茜草科(Rubiaceae)栀子属(Gardenia)常绿灌木,喜温暖湿润气候,适生性强,在贫瘠的条件下也可顽强生长,是退化边际土地生态修复的理想树种[1]。黄栀子花香四溢,具有较高的营养价值,是理想的蜜源、菜肴及精油提取原料。黄栀子的果实中含有栀子黄和栀子兰等天然可食用色素以及栀子苷、角鲨烯等生理活性成分,具有泻火除烦、清热利尿、凉血解毒、散瘀血、去黄利肝等功效,是很多中成药的重要原料[2-3]。黄栀子花朵皎洁美丽,果实为卵形橙红色,观赏性强,也可做观花、观果盆栽。综合以上,黄栀子在食品、制药、化工行业、国土绿化及生态修复等方面,具有广阔的开发应用前景,是理想的药食赏兼具型生态经济树种[4]。
关于黄栀子的研究,较多集中于扦插繁殖技术[5-6]、丰产栽培技术[7-8]、无性快繁技术[9-10]、栀子黄色素的提取[11-12]、花粉利用[13]、活性有效成分分析评价[14-15]等方面,而关于黄栀子幼苗施肥方面的研究较少。孙刚等[16]对栀子进行田间磷、钾配施发现,可提高产量;黄树军等[17]开展了有机肥对栀子生长影响的研究,初步得出施肥能够促进苗木生长;以上研究尽管揭示了施肥对黄栀子生长的影响,但关于施肥对黄栀子光合特性影响的研究较少。为此,本研究以1年生黄栀子实生苗进行土壤盆栽试验;试验肥料分别为氮肥、磷肥、钾肥,以不施肥处理作为对照,施肥设计3种组合,共4个处理,每个处理3次重复,每个重复10株,共120株;处理30 d后,进行各评价指标的测定。以黄栀子幼苗株高、地径、叶面积、叶片叶绿素质量分数、光合日变化以及光饱和点、光补偿点、暗呼吸速率为评价指标,分析氮磷钾平衡施肥对黄栀子光合特性的影响,优化黄栀子幼苗氮磷钾的施肥设计。旨在为黄栀子良种壮苗生产及高效栽培管理提供参考。
1 材料与方法
试验采用1年生黄栀子(Gardeniajasminoides)实生苗,选择生长情况基本一致的苗木于2020年5月份移植到塑料盆里(盆体规格:上口径18 cm、下口径15 cm、高20 cm),育苗基质为珍珠岩和黄土混合,比例为V(珍珠岩)∶V(黄土)=1∶4,基质表面距离盆钵边缘1 cm左右,1盆1株。
试验设计:借鉴已有研究成果,参考相关经济林果1~2年生苗木[18]及黄栀子生长所需的施肥种类与用量,制定黄栀子施肥试验设计。试验肥料分别为氮肥(尿素,N质量分数为46%)、磷肥(过磷酸钙,P2O5质量分数为14%)、钾肥(硫酸钾,K质量分数为60%),以不施肥处理作为对照(CK),共4个处理(见表1),每个处理3次重复,每个重复10株,共120株。缓苗1个月,开始处理,采用浇灌施肥方法,于2020年6月份施肥1次。处理30 d后,测定各指标。
生长量与生物量的测定:应用卷尺测定株高、游标卡尺测量地径、LI-3000C叶面积仪测定叶面积,每个处理选用9株标准株进行测量,取其平均值。生物量的测定,每个处理选取3个标准株,从基质中挖出,洗净根部,置于干燥箱中105 ℃烘干至恒质量,测量其生物量。

表1 黄栀子幼苗施肥试验设计
叶绿素质量分数测定:参照张志良等[19]的方法测定叶绿素质量分数。从每个处理中选择9株标准株,分别采集成熟新鲜叶片,各称取约0.2 g,用体积分数为95%的乙醇提取叶绿素,提取液用紫外可见分光光度计在649、665 nm比色,并计算总叶绿素质量分数(w)、叶绿素a质量分数(wa)、叶绿素b质量分数(wb)、wa∶wb,求平均值。
光合日变化及光响应曲线测定:应用LI-6400便携式光合仪测定,选择晴天测定光合日变化,从07:00—17:00每2 h测定1次,每个处理选取3株标准株,测定3片生长健康、长势一致的叶片;光响应曲线测定于08:00—11:30,测定黄栀子叶片的净光合速率(Pn)对光强的响应曲线。每处理随机选取3株生长良好的黄栀子苗木作为测定样本,选择植株中间成熟叶片测量,每株测量3枚叶片。测定时光强由强到弱,依次设定光合有效辐射(RPA)为2 000、1 600、1 200、800、600、400、200、150、100、80、60、40、20、0 μmol·m-2·s-1,测定各处理叶片的净光合速率(Pn)、气孔导度(Gs)、胞间CO2摩尔分数(Ci)、蒸腾速率(Tr)等光合参数。
光响应曲线拟合,采用直角双曲线修正模型[20]:Pn=[αRPA(1-βRPA)]/(1+γRPA)-Rd。α为光响应曲线的初始斜率,β、γ为系数,RPA为光合有效辐射,Rd为暗呼吸速率。根据模型拟合,求出表观量子效率(YAQ)、暗呼吸速率(Rd)、最大净光合速率(Pn,max)、光饱和点(PLS)、光补偿点(PLC)。
数据处理:采用Excel2019进行数据处理,采用SPSS19.0软件进行光响应曲线拟合和显著性检验(最小显著差异法)。
2 结果与分析
2.1 不同施肥配比对黄栀子幼苗生长的影响
本研究比较分析了不同施肥处理的黄栀子幼苗生长量及生物量(见表2)。由表2可见:T1、T2、T3施肥处理,幼苗的株高、地径、叶面积、生物量与CK相比均显著增加,其中株高分别增加30.48%、23.90%、25.49%,地径分别提高23.77%、9.84%、22.34%,叶面积分别增加68.95%、61.51%、63.95%,生物量分别增加58.05%、51.72%、78.13%。3个施肥处理间,除株高和叶面积差异不显著外,T3与T1、T2处理的地径和生物量均存在显著差异。
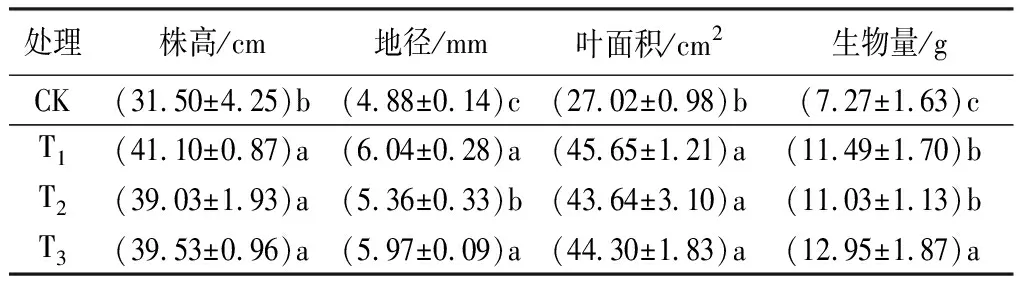
表2 不同施肥配比下黄栀子幼苗生长指标
2.2 不同施肥配比对黄栀子幼苗叶绿素质量分数的影响
由表3可见:3个施肥处理的黄栀子幼苗叶片叶绿素各指标,与CK相比均呈现出明显增加的趋势。与CK相比,T1、T2、T3处理的叶绿素a质量分数,分别显著增加84.85%、84.85%、101.01%;叶绿素b质量分数,分别显著提高了92.86%、85.71%、100.00%;叶绿素(a+b)质量分数,分别显著提升了85.92%、83.80%、99.30%。其中T3处理的叶绿素质量分数增加最大,叶绿素(a+b)质量分数达到2.83 mg·g-1,接近CK叶绿素(a+b)质量分数的2倍。
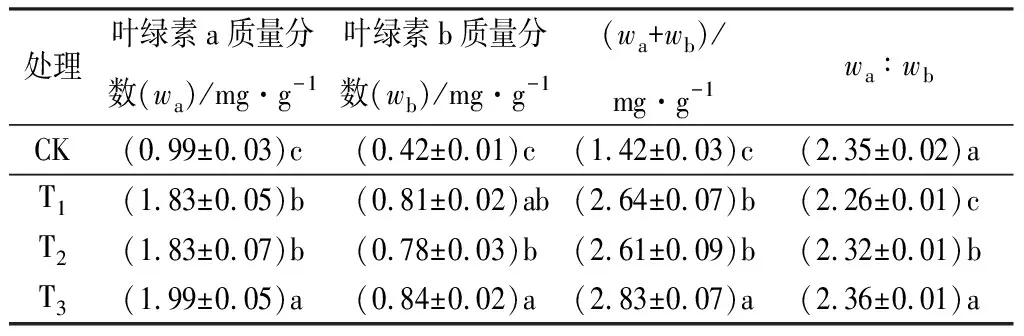
表3 不同施肥配比时黄栀子幼苗的叶绿素质量分数
2.3 不同施肥配比对黄栀子光合日变化的影响
2.3.1不同施肥配比时黄栀子幼苗净光合速率的日变化
由图1可见:不同施肥配比时,黄栀子幼苗净光合速率的日变化趋势大致相同,均呈“双峰”曲线。从07:00开始净光合速率开始增加,随着时间的后延,各施肥处理的净光合速率逐渐增大,均在09:00时达到第一个峰值;之后净光合速率逐渐下降,在11:00左右开始出现“光合午休”现象,随后又逐渐增加,并于13:00—15:00之间出现第二个峰值。各施肥处理的首个峰值差异较大,净光合速率由大到小依次为T3(9.95 μmol·m-2·s-1)、T2(9.76 μmol·m-2·s-1)、T1(7.30 μmol·m-2·s-1)、CK(6.88 μmol·m-2·s-1);第二峰值没有显著性差异,但均高于CK。
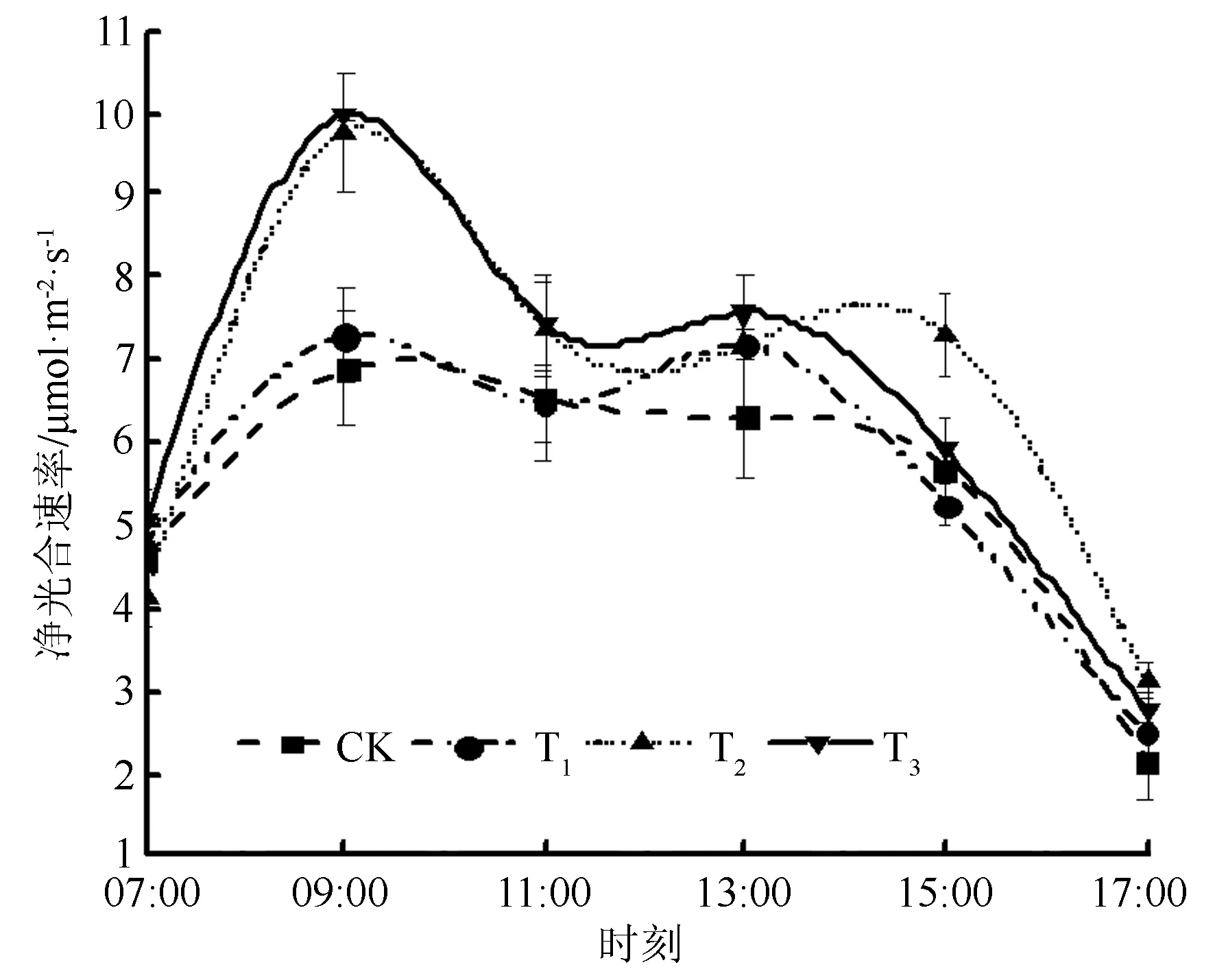
图1 不同施肥配比时黄栀子幼苗净光合速率日变化
2.3.2不同施肥配比时黄栀子幼苗气体交换参数的日变化
由图2可见:CK、T1、T2、T3各施肥处理,气孔导度日变化均呈“双峰型”。从07:00开始各施肥处理的气孔导度随着时间的增加而逐渐增大,直到09:00出现第一次峰值,分别为0.106 7(CK)、0.128 1(T1)、0.130 7(T2)、0.131 0 μmol·m-2·s-1(T3);之后逐渐减小,直到11:00左右开始出现“光合午休”现象,后又逐渐增加,13:00出现第二次峰值,分别为0.101 4(CK)、0.104 8(T1)、0.105 8(T2)、0.110 8 μmol·m-2·s-1(T3),最后随着时间的推移,逐渐减小。不同施肥处理的气孔导度峰值中,T3处理均为最大,两次峰值分别高出了CK的22.77%、9.27%。
各施肥处理的蒸腾速率(Tr)日变化呈现“单峰型”。从早上07:00开始逐渐增加,随着时间的延长逐渐增大,直到13:00,不同施肥处理的Tr达到最大值,随后逐渐减小。不同施肥处理的Tr峰值,由大到小依次为T3(5.824 4 μmol·m-2·s-1)、CK(5.820 7 μmol·m-2·s-1)、T1(5.461 0 μmol·m-2·s-1)、T2(5.221 9 μmol·m-2·s-1)。
不同施肥处理胞间CO2摩尔分数(Ci)的日变化,呈现出相似的“L型”变化曲线。随着时间的后延,胞间CO2摩尔分数从07:00的最高值快速降低,到09:00左右,各施肥处理的胞间CO2摩尔分数逐渐呈现出缓慢减小的趋势,17:00左右达到最小值。胞间CO2摩尔分数在整个日变化过程中,CK均高于其他施肥处理,这与CK光合速率较低,消耗CO2量少有一定的关系。
2.4 黄栀子幼苗光合和气体交换参数间的相关性
由表4可见:不同处理间黄栀子幼苗气体交换参数间存在一定的相关关系。黄栀子幼苗的净光合速率,与气孔导度、蒸腾速率呈正相关关系,与胞间CO2摩尔分数呈负相关关系;蒸腾速率,与气孔导度呈极显著正相关关系,与胞间CO2摩尔分数呈正相关关系;气孔导度与胞间CO2摩尔分数呈正相关关系。本研究结果进一步说明了不同施肥处理的胞间CO2摩尔分数(Ci)日变化与光合速率日变化大小相悖的现象。
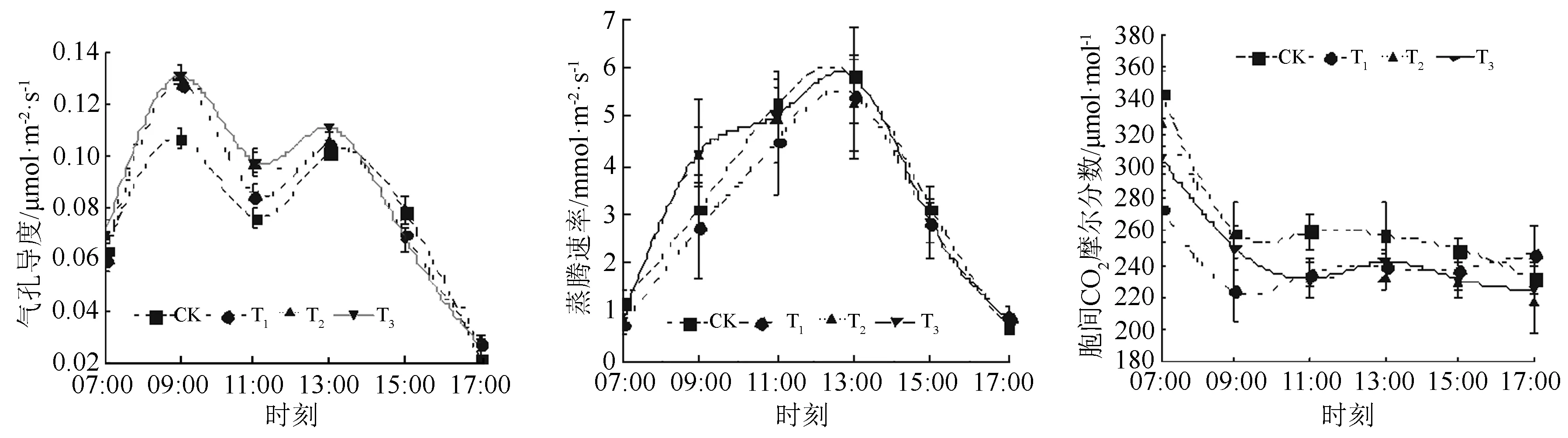
图2 不同施肥配比时黄栀子幼苗气孔导度、蒸腾速率、胞间CO2摩尔分数日变化

表4 黄栀子幼苗气体交换参数间的相关系数
2.5 不同施肥配比对黄栀子幼苗叶片光合参数的影响
根据拟合方程可得出不同施肥配比的黄栀子叶片的最大净光合速率、暗呼吸速率、光饱和点、光补偿点等相关光合参数及表观量子效率。由表5可见:各施肥处理间,表观量子效率差异不显著;与CK相比,T1、T3处理的最大净光合速率分别显著增加了33.33%、51.55%,T2处理的最大净光合速率仅提高10.97%,差异不显著;各施肥处理的光饱和点与CK相比均有所增加,其中T2、T3处理分别显著增加18.57%、12.29%(P<0.05)。与CK相比,T1、T2施肥处理的光补偿点,分别下降了15.42%、2.37%,T3则显著降低了32.22%;各施肥处理的暗呼吸速率与CK相比均有减少,其中T1处理减少了10.97%,差异不显著,T2、T3处理则显著下降了36.37%、33.89%。

表5 不同施肥配比时黄栀子幼苗的光合参数
3 讨论
光合作用是植物生长的物质和能量来源,通过施肥可影响植物光合产物的合成与积累,进而影响植物的生长。叶绿素可以捕获光能并传递到光反应中心,影响植物光合作用。氮是叶绿素的主要成分,影响植物体内光合酶类的合成与活性,进而提高净光合速率;磷供应促进1,5-二磷酸核酮糖的再生,促进光合产物从叶片中输出;钾可使叶绿体基粒数增多,提高光合电子传递链活性剂光合磷酸化活力,提高净光合速率。张卫强等[21]研究表明,合理施用氮肥有利于银叶树的生长,提高叶绿素质量分数和净光合速率。吴家胜等[22]研究表明,钾元素可显著提高植物叶片中叶绿素质量分数,以此显著增强植物的光合作用。本研究结果表明,3个施肥处理均提高了叶绿素质量分数,这主要是氮磷钾肥营养元素为叶绿素的合成提供了物质基础。在本研究施肥范围内,并未达到施肥阈值,因此,氮、钾不断增施达到最大值,即T3处理对于叶片光合作用效果最为显著。
“单峰”型[23]、“双峰”型[24]、“三峰型”[25]曲线,是目前研究光合日变化中常见的三种表现形式。本研究中,不同处理的黄栀子幼苗的净光合速率,分别在09:00左右和13:00左右出现峰值,日变化呈现“双峰型”,说明黄栀子幼苗存在“光合午休”现象。导致“光合午休”现象的主要有气孔因素和非气孔因素,判断是否为气孔因素主要看气孔导度和胞间CO2摩尔分数受外部环境影响是否一致,若一致,则为气孔因素,反之为非气孔因素[26]。本研究不同施肥处理的黄栀子气孔导度均为双峰变化趋势,胞间CO2摩尔分数变化均为不标准的“L”型,两者均在09:00—11:00逐渐降低,变化趋势具有一定的相似性;同时又根据黄栀子幼苗气孔导度与胞间CO2摩尔分数呈正相关关系分析结果,所以可判断黄栀子幼苗“光合午休”现象主要受气孔因素影响。
相关研究表明,不同的施肥处理能够刺激植物对环境的适应性,进而影响植物的光合速率,提升植物对光能的利用效率,改善植株品质[27]。本研究根据不同施肥处理的净光合速率比较分析,CK在各个时间点的净光合速率都比较低,在磷肥不变的前提下,随着氮肥和钾肥的增施,净光合速率逐渐增大,处理T3的净光合速率最大。一方面,由于施肥增大了叶片的气孔导度,从而增加了CO2的供应,净光合速率变大;另一方面,是施肥提高了土壤养分含量,有利于根系对养分的吸收与利用,使叶面积增加,改善了光合作用的条件,提高了光合利用效率。这与张青青等[28]对柚木的研究结果相近。
光响应曲线是研究植物光合生理生态的重要途径之一,对于了解光能利用效率有很大的意义[29]。相关研究表明,具有高光饱和点和低光补偿点的植物,对光的适应能力越强[30]。本研究中,与CK相比,各施肥处理的光饱和点均有所增加,光补偿点和暗呼吸速率均有所降低,其中T3最为显著,表明施肥能够增强黄栀子对光的适应性,提高植株对光能的利用率。而产生这一现象的原因,是由于氮、磷施肥量增加,提高黄栀子叶片中核酮糖-1,5-二磷酸羧化酶活性,使其适应更广的光照利用范围,增长光合反应的时间,提高了对强光和弱光的利用率,积累更多光合产物。这与杨腾等[31]研究施氮可以提高文冠果幼苗的最大净光合速率、表观量子效率、光饱和点,增强光能利用效率,提高光合能力的研究结果相近。刘红云等[32]、吴焦焦等[33]的研究结果也表明,合理施肥可以促进丹参和黄栌的净光合速率、最大净光合速率、光饱和点,降低光补偿点,增强光合能力。可见在本试验设置中,随着氮肥和钾肥质量浓度的增加,黄栀子幼苗光合潜力更强,更有利于植株的生长。
4 结论
供试的施肥配比均能有效促进苗木生长,且在施肥的作用下能有效提高各处理的苗木叶绿素质量分数,利于促进苗木光合作用以及营养物质的积累;黄栀子幼苗叶片光合速率日变化呈“双峰”变化趋势(峰值出现的时间分别为09:00和13:00),存在“光合午休”现象,且净光合速率,与气孔导度呈正相关性、与胞间CO2摩尔分数呈负相关性;T3处理与其他施肥处理组相比,净光合速率最高,光饱和点也较高,补偿点和暗呼吸速率较低,光合潜力最强。本研究可为黄栀子良种壮苗培育及精准栽培提供参考,同时也可为黄栀子产业健康发展提供技术支撑。
猜你喜欢
生态学报(2024年1期)2024-01-25 07:05:16
湖北农业科学(2021年13期)2021-08-04 03:56:18
新疆农业科技(2021年2期)2021-07-03 06:39:34
生态学报(2021年6期)2021-04-25 08:24:42
农业工程学报(2021年3期)2021-04-15 10:03:14
节水灌溉(2020年9期)2020-09-16 00:32:12
中学生物学(2020年12期)2020-03-30 07:27:46
新课程·下旬(2017年7期)2017-08-14 11:12:50
西北林学院学报(2015年5期)2015-01-03 05:12:20
生物技术通报(2014年10期)2014-03-21 06:56:38
