
Intraspecific variation in shoot flammability in Dracophyllum rosmarinifolium is not predicted by habitat environmental conditions
2022-06-10 07:34XingliCuiArinPtrsonGorgLWPrrySrhWysAzhrulAlmCongHungShixingZhouLinXioChnghongLiFngDongyuCoKtMrshllTimothyCurrn
Forest Ecosystems 2022年2期
Xingli Cui, Arin M. Ptrson, Gorg LW. Prry, Srh V. Wys, M Azhrul Alm,Cong Hung, Shixing Zhou, Lin Xio, Chnghong Li, Fng H, Dongyu Co,Kt Mrshll, Timothy J. Currn
a Department of Pest-management and Conservation, Lincoln University, Lincoln, 7647, New Zealand
b National Forestry and Grassland Administration Engineering Research Centre for Southwest Forest and Grassland Fire Ecological Prevention,College of Forestry,Sichuan Agricultural University, Chengdu, 611130, China
c School of Environment, University of Auckland, Auckland, 1142, New Zealand
d Bio-Protection Research Centre, Lincoln University, Lincoln, 7647, New Zealand
e Sichuan Province Forestry and Grassland Environmental Monitoring Centre, Chengdu, 610036, China
Keywords:Dracophyllum Fire Flammability Habitat
ABSTRACT Background: Flammability is a compound plant trait that can vary significantly across natural populations within species. Investigating intraspecific variation in flammability provides insights into the evolution of plant flammability and inform understanding of wildfire risk and behaviour in different habitats.Methods:We measured four flammability variables,representing ignitibility(time to ignition),sustainability(total burning time),combustibility(maximum temperature during burning)and consumability(percentage of biomass consumed by fire)to assess the shoot-level flammability of Dracophyllum rosmarinifolium(G.Forst.)(Ericaceae),a polymorphic endemic species distributed throughout New Zealand. We examined the relationship between flammability components and a suite of climatic and geographic variables (elevation, latitude, mean annual temperature (MAT), mean annual rainfall (MAR) of the sample locations, etc.).Results: We measured shoot-level flammability components of 62 individuals across eight populations. Burning time, maximum temperature and burnt biomass were positively correlated with each other, while ignition score was independent of other flammability components. All flammability components varied significantly across the eight populations. The habitat conditions we considered were not related to any of the shoot-level flammability components of D. rosmarinifolium.Conclusions:Intraspecific variation in flammability in D.rosmarinifolium may be a byproduct of selection on other functional traits, such as leaf size, shoot lipid content, indicating that plant flammability is an incidental result,rather than selected for, at least in ecosystems without fire as a selective force.
1. Introduction
Climate change is exacerbating the chance of wildfires in many parts of the world.Plant flammability,which indicates the ability of plants to burn and sustain a flame when exposed to an ignition source, is an important determinant of fire behavior in terrestrial ecosystems and plays a fundamental role in ecosystem dynamics (Pausas et al., 2017;Prior et al.,2017).Plant flammability varies widely among(Calitz et al.,2015; Wyse et al., 2016; Cui et al., 2020a) and within species (Pausas et al.,2012;Battersby et al.,2017).Identifying the factors that influence plant flammability will inform prediction of changes in wildfire risk and fire behaviour in a given environment, and provide insights into the evolution of plant flammability.
The micro-evolutionary approach, which involves investigation of variation in traits within species or populations,is useful for identifying the factors that may have influenced the evolution of plant traits(Pausas,2015).Pausas et al.(2012)showed that individuals of the Mediterranean shrub species Ulex parviflorus(Mediterranean gorse,Fabaceae)from sites with a history of frequent fire were more flammable than those growing in sites without recurrent fire (based on fire records since 1978). Flammability of U. parviflorus has likely increased due to recurrent fires(Pausas et al., 2012; Moreira et al., 2014). In Leptospermum scoparium(m-anuka, Myrtaceae), serotinous populations were rarely found at sites with no history of fire during the Holocene, and serotiny was generally more common (higher proportion of closed capsules) in populations where the landscape had experienced some fire(Battersby et al.,2017).However, no association was found between m-anuka shoot-level flammability and Holocene site fire history, nor flammability and serotiny(Battersby et al.,2017).
Nevertheless, micro-evolutionary studies of plant flammability are rare, and most have attempted to relate flammability to fire regimes in fire-prone regions (Pausas et al., 2012; Moreira et al., 2014). Whether and how environmental conditions, other than fire regime, have influenced the intraspecific variation of plant flammability is poorly understood. Although the hypothesis that flammability is selected by the ecosystem fire regime has received support (Bond and Midgley, 1995;Archibald et al., 2018), it is possible that flammability is an emergent property, whereby increased flammability is an incidental or secondary byproduct of selection for other traits.Such traits may include water-use efficiency,nutrient retention,etc.,which increased individual fitness and incidentally changed the flammability (Gould and Vrba, 1982; Snyder,1984; Midgley, 2013). Micro-evolutionary studies of plant flammability in ecosystems in the absence of fire as a selective force may offer insights into the evolution of plant flammability.In addition,understanding how environmental conditions can affect the flammability of plant can facilitate fire management under climate changing. However, such studies have scarcely been reported.
We investigated whether environmental conditions, other than fire regime, are correlated with intraspecific variation of plant shoot flammability. We collected and measured the shoot flammability of 62 D. rosmarinifolium individuals from eight populations across New Zealand, a region with a history of low fire frequency (Perry et al., 2014;Kitzberger et al.,2016)and where most ecosystems are unlikely to have been influenced by fire prior to human arrival(Ogden et al.,1998;Lawes et al., 2014; Perry et al., 2014). We examined variation in shoot-level flammability among populations and sought to identify the geographic and environmental factors(elevation,latitude and bioclimatic variables)that predict intraspecific variation in plant flammability.
2. Materials and methods
2.1. Sample collection
Dracophyllum Labili.(Ericaceae)occurs in Australia,New Zealand and nearby oceanic islands, and reaches its highest species richness and morphological diversity in New Zealand, ranging from low-growing subshrubs to trees (Wagstaff et al., 2010; Venter, 2021). Dracophyllum rosmarinifolium is an extremely polymorphic species and occurs in different habitats across New Zealand:mountain gullies,mountain slopes ranging from 0 to 80°in steepness, ridges, bluffs, plateaus and valley floors from ca. 150–2,100 m elevation (Venter, 2009). Although it is unclear whether D. rosmarinifolium has adapted to local conditions, it shows considerable variation in morphological characters among populations, such as leaf length, branching habit and plant height (Venter,2009). These polymorphic characteristics and its occurrence in diverse habitats make D. rosmarinifolium a good model for micro-evolutionary study.
We collected shoot samples of D. rosmarinifolium from eight sites across the South Island, New Zealand based on information on species distributions from Venter (2009), iNaturalist (https://inaturalist.nz/),and the Allan Herbarium. The eight collection sites spanned a large proportion of the latitudinal range of D. rosmarinifolium in the South Island, New Zealand and showed obvious variation in environment(Fig. 1); All sites are believed to have a historical low fire frequency, as fires were infrequent across most of New Zealand prior to human settlement,with the exception of some wetland systems in the North Island(Ogden et al.,1998;Rogers et al.,2007;Perry et al.,2014).Eight samples of D.rosmarinifolium were collected from each site(except for two of the sites, where seven samples were collected). Healthy terminal shoots of 70-cm length with leaves were cut from healthy individuals, preserving branch architecture, and kept in separate sealed plastic bags to prevent moisture loss.Shoot samples were kept cool during collection and were then stored at 4–8°C on the same day. All samples were collected over one summer (2018–2019) to eliminate the influence of seasonal and annual variability,and were burned within one week of collection.
2.2. Flammability measurement

Fig.1. Map of New Zealand displaying collection sites in this study.Left-top of the figure shows the environmental condition of collection and observation sites. Blue circles indicate the collection sites. Orange diamonds indicate the observations (until May 2019 on iNaturalist) of D. rosmarinifolium, showing the range of D. rosmarinifolium.
To assess the flammability of a plant, four components of flammability are often considered: ignitibility (how easily a plant ignites), sustainability(the length of time a plant sustains flame),combustibility(the intensity at which a plant burns) and consumability (the percentage of biomass consumed by fire) (Anderson, 1970). Here, shoot flammability was measured following the methods described by Jaureguiberry et al.(2011) and Wyse et al. (2016), using the same device as Wyse et al.(2016). Prior to burning, all shoot samples were air-dried at room temperature for 24 h to match the sample moisture content to the ignition source(Wyse et al.,2016,2018).Samples were first placed on our device for preheating for 2 min at 150°C.Then,a blowtorch was turned on for 10 s to ignite the samples. Ignitability was represented by an ignition score (Padull′es Cubino et al., 2018), and was recorded first as time to ignition (between 0 and 10 s), which was subsequently converted to an ignition score by subtracting the time to ignition from 10;e.g.,a sample that took 1 s to ignite (i.e. rapid ignition) had an ignition score of 9.Samples that did not ignite after 10 s were given a zero value.The reason why we use ignition score(but not ignition time)to represent ignitability is that a high ignition score represents a fast ignition and high ignitibility.The maximum temperature of flames during burning was recorded using an infrared laser thermometer (Fluke 572; Fluke Corp., Everett, WA,USA) to represent combustibility. We start measuring when samples begin to burn until flame extinguished. The maximum temperature during the burning was recorded by the infrared laser thermometer.Samples that failed to ignite were given a value of 150°C, representing the grill temperature (Padull′es Cubino et al., 2018). Sustainability was measured as the period that a sample remained burning (had flaming combustion) after the blowtorch was turned off. Consumability was measured as the mean percentage of burnt biomass after flaming combustion ceased,assessed by visual observation by at least two observers.Samples that did not sustain a flame after the blowtorch was turned off were assigned zero for sustainability and consumability.
2.3. Data collection
Latitude,longitude and elevation of the collection sites were recorded during collection by using Garmin GPSMAP 64. Bioclimatic variables,including mean annual temperature (MAT), mean temperature of warmest quarter, mean temperature of coldest quarter, mean temperature of wettest quarter,mean temperature of driest quarter,mean annual rainfall(MAR),mean rainfall of warmest quarter,mean rainfall of coldest quarter, mean rainfall of wettest quarter and mean rainfall of driest quarter, for each collection site were obtained from WorldClim version 2.1(https://www.worldclim.org/) at a spatial resolution of 1 km2(Fick and Hijmans,2017).
To calculate the moisture content of each sample, a sub-sample of twigs and leaves was collected and weighed to determine their fresh mass(FM)before 24-h drying.These sub-samples were oven-dried at 65°C for at least 48 h until reaching a constant weight and weighed for dry mass(DM).Moisture content(MC;%)of the sub-samples was calculated as:

2.4. Statistical analysis
Partitioned variance (%) for the flammability traits at species and individual level was calculated from Analysis of Variance (ANOVA),using the aov function in R. Principal components analysis (PCA) of the four flammability components was performed to obtain composite flammability metrics,using the princomp function in R.The flammability components, and the PC1, and PC2 scores, were compared across the eight populations using one-factor ANOVA and Tukey's tests, using the aov function in R. Geographic and environmental factors were used for correlation analysis with flammability variables of each population by using Pearson correlation, using cor function in R. To preserve the familywise error-rate, the Benjamini and Hochberg method (Benjamini and Hochberg,1995)was used for multiple testing correction with R package ppcor (Kim, 2015). All statistical analyses were conducted in R 3.5.0 (R Core Team,2018).
3. Results
3.1. Shoot flammability varies significantly across D. rosmarinifolium populations
We collected 62 shoot samples across the eight sites, ranging from 840 m to 1,310 m in elevation and from 41.198°to 44.814°S in latitude(Fig. 1, Table 1). All the samples ignited successfully during the flammability measurements. The proportion of variance in shoot moisture content and all the shoot flammability traits,except burning time, were higher between populations (shoot moisture content: 64.6%; ignition score:60.1%;maximum temperature:70.9%;burning time:44.7%;burnt biomass: 87.5%) than within populations (shoot moisture content:35.4%; ignition score: 39.9%; maximum temperature: 29.1%; burning time: 55.3%; burnt biomass: 22.5%). All flammability components and shoot moisture content varied significantly among the populations(Fig. 2, ANOVA: P < 0.001). Flammability components were not significantly related to shoot moisture content(Table 2).Individuals from Mt Rochford had the lowest value in mean shoot moisture content, while individuals from Alex knob had the highest value. Individuals from Homer Tunnel had the lowest mean value of ignition score(7.31±0.09),maximum temperature (439.50°C ± 29.34°C), and burnt biomass(33.75%±4.51%).The Lewis Pass population had the shortest burning time(23.14 s±3.89 s).The maximum temperature and burnt biomass of Homer Tunnel and Lewis Pass were both significantly lower than other populations (Fig. 2). The Lewis Pass individuals had low values for burning time (23.14 s ± 3.89 s), maximum temperature (456.00°C ±31.86°C)and burnt biomass(36.43%±2.37%),and had a high ignition score(8.71±0.10).Samples from Mt Arthur were the most flammable,with the highest mean value of ignition score (8.81 ± 0.09), maximum temperature (724.38°C ± 30.56°C), and burnt biomass (88.75% ±1.57%). The longest average burning time occurred in the Alex Knob population,with a mean of 93.88 s ±18.12 s.
A PCA was conducted with the four flammability components of all samples(Fig.3a).The first two axes of PCA explained 61.6%and 24.5%of the variation, respectively. Burning time, maximum temperature and burnt biomass were positively correlated with each other. The ignition score was independent of other flammability components(Table 2).The first axis scores represent an integrated measure of combustibility, sustainability and consumability,and the second axis describes ignitability.Multiple comparisons of PC1 and PC2 across populations showed that they varied significantly(ANOVA:P<0.001,Fig.3).The PC1 scores for the Homer Tunnel and Lewis Pass populations were significantly lower than for the other six populations, which had a similar value of PC1.Homer Tunnel samples has the lowest value of both PC1 and PC2,indicating individuals from Homer Tunnel were least flammable.Lewis Pass samples have a low value in PC1; however, these individuals also have the highest values in PC2, showing that these individuals ignited most readily.
3.2. Intraspecific variation in shoot flammability in D. rosmarinifolium is not related to habitat conditions
To explore which factors may influence intraspecific variability in flammability of D. rosmarinifolium, bioclimatic variables (10 variables)and geographic location(elevation and latitude)of collection sites were analyzed against the flammability variables. The habitat conditions we considered were not related to any of the shoot-level flammability components of D. rosmarinifolium(Fig.s1).
4. Discussion
Plant trait can change rapidly in response to altered environmental conditions (Bone and Farres, 2001). Such rapid evolutionary changes allow us to observe evolution over a short period of time and to identify its potential drivers. Many micro-evolutionary studies have facilitated our understanding of evolutionary mechanisms for different plant traits(Vazquez-Gonzalez et al.,2019;Depardieu et al.,2020;Weemstra et al.,2021). The studies of intraspecific variation in plant flammability are rare,and have focused on the influence of the fire regime(Pausas et al.,2012; Battersby et al., 2017). However, flammability could be an incidental byproduct of selection on other functional traits in response to theenvironment(such as cold and dry conditions),rather than a trait directly selected for by fire(Snyder,1984;Midgley,2013;Cui et al.,2020b).How the flammability of a species may change without fire as a selective force is still unclear. Exploring the change of flammability due to environmental factors, rather than fire regimes, can confer insights into the evolutionary mechanisms of plant flammability and facilitate fire management under climate changing. However, such studies have rarely been reported.
Fire was infrequent in New Zealand prior to human arrival in the late 13th century(Wilmshurst et al.,2008),and the evolution of New Zealand indigenous plants was not strongly influenced by fire(Ogden et al.,1998;Lawes et al., 2014; Perry et al., 2014). Dracophyllum rosmarinifolim is a New Zealand indigenous species that appeared 1.1–2.2 million years ago,probably due to climatic changes during the Late Tertiary, and now shows considerable intraspecific variation in morphological characters(Wagstaff et al., 2010; Venter, 2021). We found that shoot-level flammability components varied significantly among different populations of D. rosmarinifolim, although it is unclear whether the variation in flammability is heritable or a result of phenotypic plasticity (e.g., whether there is a genetic difference across the populations) (Schlichting, 1986;Sultan,1995).Further studies,such as reciprocal transplant experiments and genetic studies, are needed to explore whether intraspecific variability in flammability is heritable.To identify the potential factors that influenced the intraspecific variation in flammability components of D. rosmarinifolim, we examined the relationship between flammability components and habitat conditions, including elevation, latitude, MAT,MAR of the sample locations,etc.
Elevational and latitudinal gradients are common geographic factors examined in ecological and evolutionary studies(K¨orner,2007;Swenson and Enquist, 2007). A change in elevation or latitude is usually accompanied by changes in a range of environmental variables (Blumthaler et al., 1997; K¨orner, 2007), and can cause genetic and phenotypic changes within species, which may result in local adaptation (Ronghua et al., 1984; Giordano et al., 2007; Swenson and Enquist, 2007; Gonzalo-Turpin and Hazard,2009;Scheepens et al.,2010;Montesinos-Navarro et al.,2011; Halbritter et al., 2018). For example,Festuca eskia Ramond(Poaceae),a perennial alpine grass common in the Pyrenean Mountains,was found to be adapted to local climate and has a reduced plant stature with increasing elevation (Gonzalo-Turpin and Hazard, 2009). Temperature and precipitation are major climatic determinants of plant trait variation (Choat et al., 2007; Franks et al., 2007; Sandel et al., 2010;Moles et al., 2014). Krix and Murray (2018) reported that leaves of sheltered forest species(moist habitat)were significantly less flammable than exposed woodland species (dry habitat). Whether intraspecific variation in plant flammability is influenced by these habitat conditions has received little attention.
Habitat conditions did not predict shoot flammability components of D.rosmarinifolium(Fig.s1).In short,shoot flammability variation within D. rosmarinifolium may not be explained by habitat conditions. Similar findings have been reported at the Dracophyllum genus level:shoot-level flammability varies significantly across 21 Dracophyllum species,but was not correlated with latitude, elevation, MAT and MAR of habitats (Cui et al., 2020b). The apparent decoupling between flammability and habitat environment may be explained if flammability is an incidental byproduct of selection on many functional traits,rather than a trait that is selected for. The functional traits that have influenced different plant flammability components(ignitibility,sustainability,combustibility and consumability)were potentially selected for by different environmental factors.Another explanation for lack of correlation between flammability and habitat environment is that shoot-level flammability of D. rosmarinifolium was influenced by other factors, environmental or otherwise, such as soil conditions (Gibson et al., 2015) that we did not consider. A broader range of habitat conditions and more populations need to be measured to help ascertain why flammability differs among populations (Scarff and Westoby, 2008; Alam et al., 2020). A third possible explanation is that shoot flammability can change quickly due to dynamic environmental conditions,such as rainfall and drought.
The evolutionary mechanisms of flammability have been debated since the Mutch hypothesis was posited half a century ago(Mutch,1970;Snyder, 1984; Bond and Midgley, 1995; Midgley, 2013; Bowman et al.,2014; Cui et al., 2020a). Our results are consistent with plant flammability in local populations being an incidental property not selected primarily by fire, at least in environments that have not experienced frequent fire historically (Snyder, 1984; Midgley, 2013; Cui et al.,2020b). Environment selected plant traits and incidentally changed the plant flammability.However,the hypothesis that flammability has been selected for under different fire regimes has also received support in previous studies (Gagnon et al., 2010; Pausas et al., 2012; Cui et al.,2020a).We suggest that plant flammability has likely both been selected for by fire and emerged as an incidental byproduct of selection on other functional traits, depending on the context and whether a species or population has evolved in a fire-prone habitat or non-fire-prone habitat.In future studies, understanding the evolution of plant flammability should be considered in the light of identifying the roles of both adaptation and incidental result, potentially based on the fire regimes of species’ habitats.
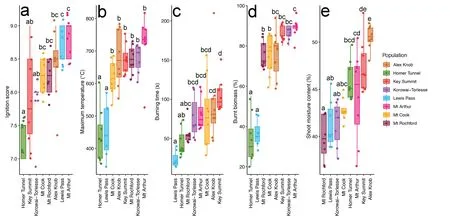
Fig.2. Variation in falmmability components and shoot moisture content across populations.a:ignition score,b:burning time,c:maximum temperature,d:burnt biomass,e:shoot moisture content.Points indicate the individual samples.Point colour indicates the populations.Whiskers indicate the 95%percentile.Populations with the same letter code are not signifciantly different,based on Tukey's multiple comparisons of means(P< 0.05).

Table 2 Pearson's correlation coefficient among four flammability variables and shoot moisture content. n =62,NS: P>0.05; ***: P< 0.001. P values were adjusted using the Benjamini and Hochberg method.
5. Conclusions
Shoot-level flammability varies significantly across natural populations of D. rosmarinifolium, but is not consistently related to habitat conditions. Intraspecific variation in flammability in D. rosmarinifolium may not be explained by contemporary environmental conditions, suggesting that shoot-level flammability of plants may be an incidental byproduct of selection on other functional traits, at least in largely firefree ecosystems. Environment selected plant traits and incidentally changed the plant flammability. Our findings contribute to understanding of plant flammability evolutionary mechanisms and provide empirical support for the hypothesis that flammability of plants can be an incidental result of selection on other functional traits.
Ethics approval and consent to participate
Not applicable.
Authors’ contributions
This project was developed and designed by XC, TC and AP. Plant samples and flammability data were collected by XC,AA,DC and KB.XC did the statistical analyses with advice from AP,TC,SW.XC,TC&AP led the writing of the manuscript, with input from all co-authors.
Funding
This work was supported by Lincoln University faculty funding(NO.3601-AGLS-45401-1128912).
Availability of data and materials
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Consent for publication
Not applicable.
Declaration of competing interest
The authors declare that they have no competing interests.
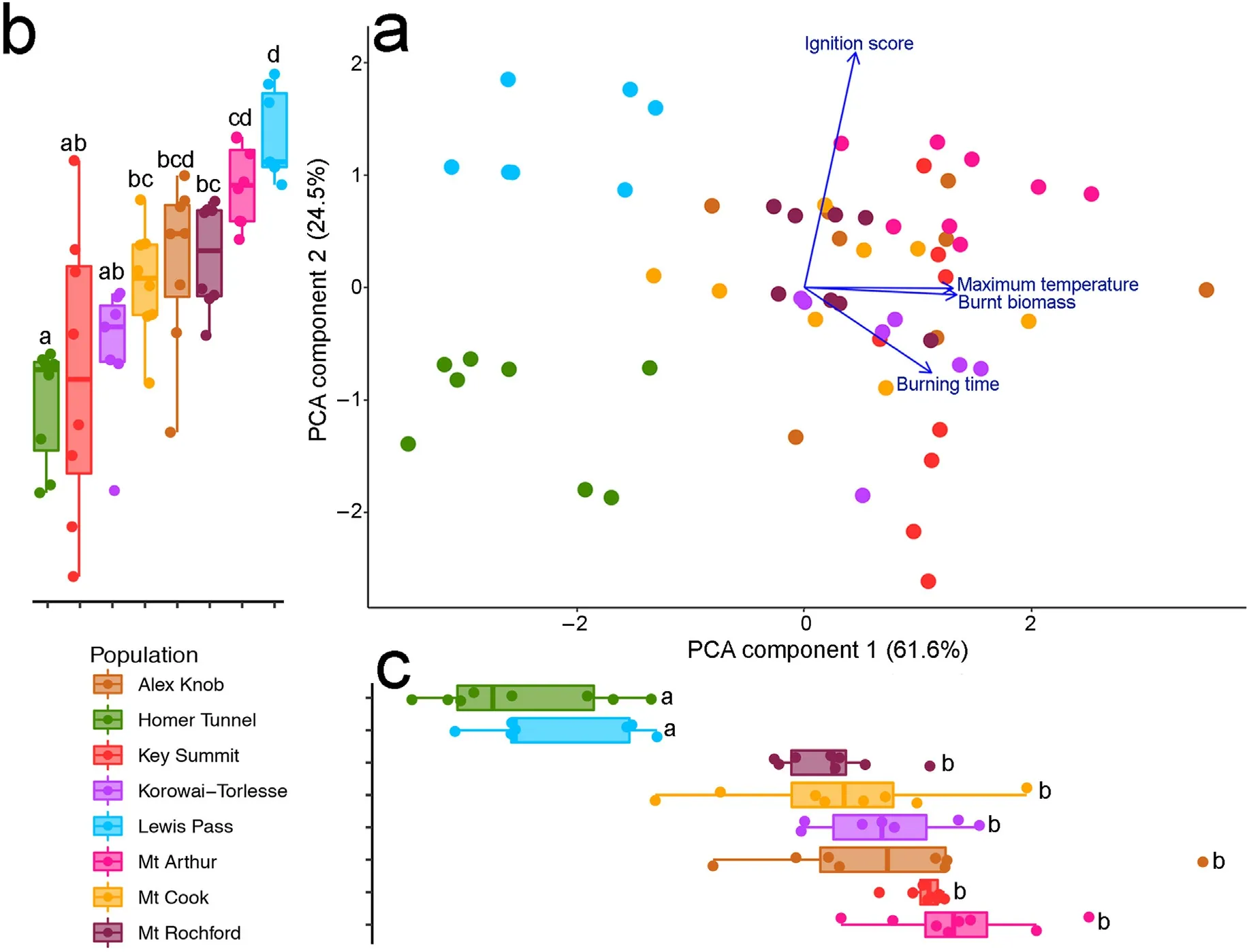
Fig.3. aa::Principal components analysis(PCA)of flammability components(ignition score,burning time,maximum temperature and burnt biomass),b:The variation of PC2 across the populations,c:the variation of PC1 across the populations.Points indicate the individual samples.Point colour indicates the populations.Whiskers indicate the 95% percentile. Populations with the same letter code are not significantly different, based on Tukey's multiple comparisons of means (P < 0.05).
Acknowledgments
We want to thank the staff at the Allan Herbarium(CHR)for access to their collection of Dracophyllum species and site information on where to collect D. rosmarinifolium. We also want to thank the staff of the Department of Conservation of New Zealand,Myles Mackintosh and Eva van den Berg for the help in the sample collection. The collection of samples was authorized by the New Zealand Department of Conservation under collection authorization 65543-FLO.
Appendix A. Supplementary data
Supplementary data to this article can be found online at https://do i.org/10.1016/j.fecs.2022.100017.

- Forest Ecosystems的其它文章
- Removing harvest residues from hardwood stands affects tree growth,wood density and stem wood nutrient concentration in European beech (Fagus sylvatica) and oak (Quercus spp.)
- Modeling of cold-temperate tree Pinus koraiensis (Pinaceae) distribution in the Asia-Pacific region: Climate change impact
- Carbon stocks in a highly fragmented landscape with seasonally dry tropical forest in the Neotropics
- Current climate overrides past climate change in explaining multi-site beta diversity of Lauraceae species in China
- Ecological niche modeling of the main forest-forming species in the Caucasus
- Impact of nitrogen input from biosolids application on carbon sequestration in a Pinus radiata forest