
藏羊和牦牛的生理特征及高原适应性分子遗传学研究进展
2022-05-27 10:38孙珂欣阿旺措吉王小龙吴玉江
家畜生态学报 2022年4期
德 吉,孙珂欣,罗 琪,阿旺措吉,王小龙,吴玉江*
(1. 西藏自治区农牧科学院 畜牧兽医研究所,西藏 拉萨 850009;2. 西北农林科技大学 动物科技学院,陕西 杨凌 712100)
高原常指海拔高度高于500 m的平原,相较于其他地理环境,高原地貌有着特殊的环境特征。以中国青藏高原为例,平均海拔高度在4 800 m以上,总面积将近300×10hm,该地区大气稀薄,光照强度高,紫外辐射强,且具有低氧、低压等特点。稀薄的空气和寒冷的环境致使该地区植被覆盖较少,进而导致青藏高原的环境蓄水保持能力较差,常年干旱,昼夜温差极大。即使环境恶劣,青藏高原仍存有非常丰富的野生物种资源如藏羚羊、藏野驴、雪豹和岩羊等,以及一些驯化物种,如牦牛、藏羊、藏鸡、藏猪等。
牦牛()是在中国高原地区广泛分布的珍稀牛种之一,其人工驯养历史十分悠久,至少在7 300年前就已被游牧民族驯化。如今,牦牛养殖已经成为藏区人民重要的生活来源,不仅作为重要的肉、奶等食物资源,还可用作驼畜使役。据统计,约有2 200×10头家养牦牛为西藏居民提供食物、交通、住所和燃料等必需品。
西藏羊(Tibetan sheep)是青藏高原地区饲养的主要草食家畜之一,主要分布在平均海拔3 500 m以上的青藏高原,是中国典型的粗毛羊品种,具有抗逆性强、耐粗饲、体格较大的优点。藏羊为毛皮用或毛肉兼用型羊,其羊肉氨基酸总量以及必需氨基酸的含量都更高,脂肪含量较低;羊毛长度长、强度高,是优质毛毯制品的重要原料来源。现有研究证明,在藏区动物中存在的HIF通路及氧气感应通路是高原动物适应低氧低温环境的最主要途径,也是现代低氧研究的热点;同时,桥粒芯蛋白3(Desmoglein3,3)、YTH N6甲基腺苷RNA结合蛋白1(YTH N6-methyladenosine RNA binding protein 1,1)等基因、解剖学及血液生理学特征也为藏羊与牦牛在低氧环境中的生存提供了有力保障。除此之外,严厚的被毛及独特的瘤胃微生物群落组成,也使得藏羊与牦牛在低温、饲料短缺的环境中具有强大的生存能力。
1 藏羊与牦牛的高原适应性进化
高寒、低氧、强紫外线是藏区环境最为明显的特征。随着海拔高度增加,氧气浓度几乎呈线性下降,且海拔每升高1 km,气温也会下降约6 ℃。除对动物生存造成巨大挑战以外,青藏高原的低温环境也导致了饲料供应不足。高原动物为抵抗严酷的环境条件,进化出了独特的生理特征以及高原适应性基因。
1.1 藏羊与牦牛适应藏区环境的特殊生理特征
如何应对低氧低温环境是牦牛面对的最严峻的问题。为此,牦牛进化出了一系列适应藏区环境的解剖及生理学特征:如体型大而紧凑,脖子较短,前躯较后躯发达;被毛粗且硬,密且长,皮下脂肪厚,有助于抵御寒冷侵袭及紫外照射;缺氧环境导致其呼吸频率高,体内血液流动快,血红蛋白含量高,心肺发达,肺泡增大。此外,牦牛缺乏缺氧性肺血管收缩,这一生理性状有助于提高氧气的摄取并提高氧气的利用率。陈秋生等通过系统观测牦牛肺脏的微细结构发现,与正常动物相比,构成牦牛肺泡壁的肺泡Ⅰ型上皮较薄且有间断处,气血屏障较薄,各级呼吸系统都具有较好的弹性,为牦牛呼吸时增加气体交换、防止低氧条件下肺动脉压急剧升高提供了保障。为了应对低温环境带来的饲料短缺问题,牦牛的瘤胃微生物相较于其他牛科动物具有较大差异,瘤胃上皮细胞独特的肽运输环境能够促使其在青藏高原恶劣的环境下更加高效地利用蛋白,以达到充分利用饲料的目的。
藏羊同样具有独特的抗逆特征。相较于普通绵羊,藏羊体型更大且面容粗犷,四肢粗壮有力,具有极强的攀岩能力;被毛粗且密,能够有效的抵御严寒、风沙及强紫外照射。王欣荣和吴建平通过对藏羊与滩羊的脑组织解剖对比,发现两者脑动脉系统结构虽然大致相同,但藏羊具有大脑后交通动脉、上颌动脉以及其他脑动脉较粗的特点,使得藏羊具有格外强的供血能力,在低氧环境下仍能保证脑部所需氧气的正常输送。为进一步探索藏羊高原适应性机理,卢晓丽等对藏羊的血液生理学特征进行了研究,发现藏羊的白细胞总数、血红蛋白等指标显著高于林芝河谷型绵羊,证明藏绵羊能够通过增加白细胞数增强免疫力,增加血红蛋白携氧能力。
1.2 藏羊与牦牛对低氧条件的适应性进化
在高海拔地区的诸多恶劣因素中,对生物影响最大的是低氧。随着海拔高度的增加,大气压急剧下降,但大气中氧气始终维持在一定比例保持不变,高海拔地区氧分压较低,表现出强烈的低压缺氧。动物机体的活动,尤其是大脑的运转离不开氧气的供应,长期的低氧环境还会造成昏迷、癫痫甚至脑死亡。同时,低氧环境会引起胃肠粘膜上皮细胞缺氧,导致缺氧诱导因子(hypoxia inducible factor-1,HIF)的激活。不仅如此,低氧还与大量的疾病发生相关。如Sun等发现,在低氧条件下,阿茨海默症的发病率明显上升,且与糖尿病与癌症等各种人类疾病密切相关。因此,对在藏区生活的哺乳动物来说,如何适应低氧环境,保持正常生活,成为了巨大的挑战。
1.2.1 低氧适应过程的主要通路——HIF通路 在牦牛、藏獒等藏区哺乳动物中存在着一条共有的HIF低氧诱导途径,根据其共有性推测,这可能是长期在低氧环境中生存的动物所共有。HIF是一个异二聚体,由1个共同的β亚基(HIF-β,由基因编码)和3个α亚基HIF-1α、HIF-2α和HIF-3α之一组成,2(或1)、egl-9家族低氧诱导因子1(egl-9family hypoxia-inducible factor 1,1)(或2)和脯氨酸羟化酶3(egg-laying deficient nine-like protein 3,3)(或3)基因编码的酶能够使HIF-α在正常氧气浓度下羟基化,HIF-α与VHL(von Hippel-Lindau)蛋白结合,从而激活泛素连接酶系统,导致HIF-α蛋白酶降解;HIF-α中脯氨酸残基的羟基化对于VHL结合至关重要,二者结合还依赖于α-酮戊二酸依赖性双加氧酶、脯氨酰羟化酶(prolyl hydroxylase,PHD)和天冬酰胺基羟化酶;在缺氧期间,PHD不活跃,导致HIF-α稳定并与HIF-1β二聚。二聚化后,HIF易位至细胞核,以结合启动子区域内的E-box 缺氧反应元件(hypoxia response elements,HRE);HIF激活控制细胞氧稳态的基因,包括与耗氧、红细胞生成、血管生成和线粒体代谢有关的基因。对藏区动物来说,细胞通过HIF调控,提高糖酵解降低氧化代谢来降低耗氧量,并通过减少细胞分裂等细胞过程的能量需求来适应低氧环境(图1)。除对缺氧适应性调节以外,HIF通路也被发现与多种肿瘤发生相关,为使用小分子干扰提供了理想靶标。

图1 低氧适应过程的主要通路-HIF通路的作用机制(改自文献[16])
1.2.2 氧气感应途径 藏羊长期生活并广泛分布于青藏高原,是研究高原适应性的良好材料。Wei等通过对7个分布于不同高海拔和低海拔地区的绵羊品种进行基因分型,检测其全基因组范围内 单核苷酸多态性(single nucleotide polymorphism,SNP),发现了一系列与高原低氧环境适应性有关的候选基因,包括内皮PAS1蛋白(endothelial PAS domain-containing protein 1,1)、αA晶状体蛋白(crystallin alpha A,)、线粒体离子肽酶1(mitochondrial ion peptidase 1,1)、神经纤维瘤1型(Neurofibromatosis,1)、二肽基肽酶-4(Dipeptidyl peptidase-4,4)、超氧化物歧化酶1(Superoxide dismutase,1)、过氧化物酶体增殖物激活受体(Peroxisome Proliferator-activated Receptor gamma,)以及细胞因子信号转导抑制因子2(suppressor of cytokine signaling 2,2),揭示了绵羊高原环境适应的遗传机制,尤其是1基因在高原适应性中的作用;1基因为重要的高原低氧适应性基因,与稳定HIF-1α因子的EGLN1(EGL9同系物)不同,在低氧条件下,1将被激活,其产物能够与DNA结合,从而激活低氧反应基因,包括促红细胞生成素(Erythropoietin,)、血管内皮生长因子基因(vascular endothelial growth factor,)和血小板衍生生长因子B基因(platelet-derivedgrowthfactor,)等,用以调节血管内稳态(图2)。
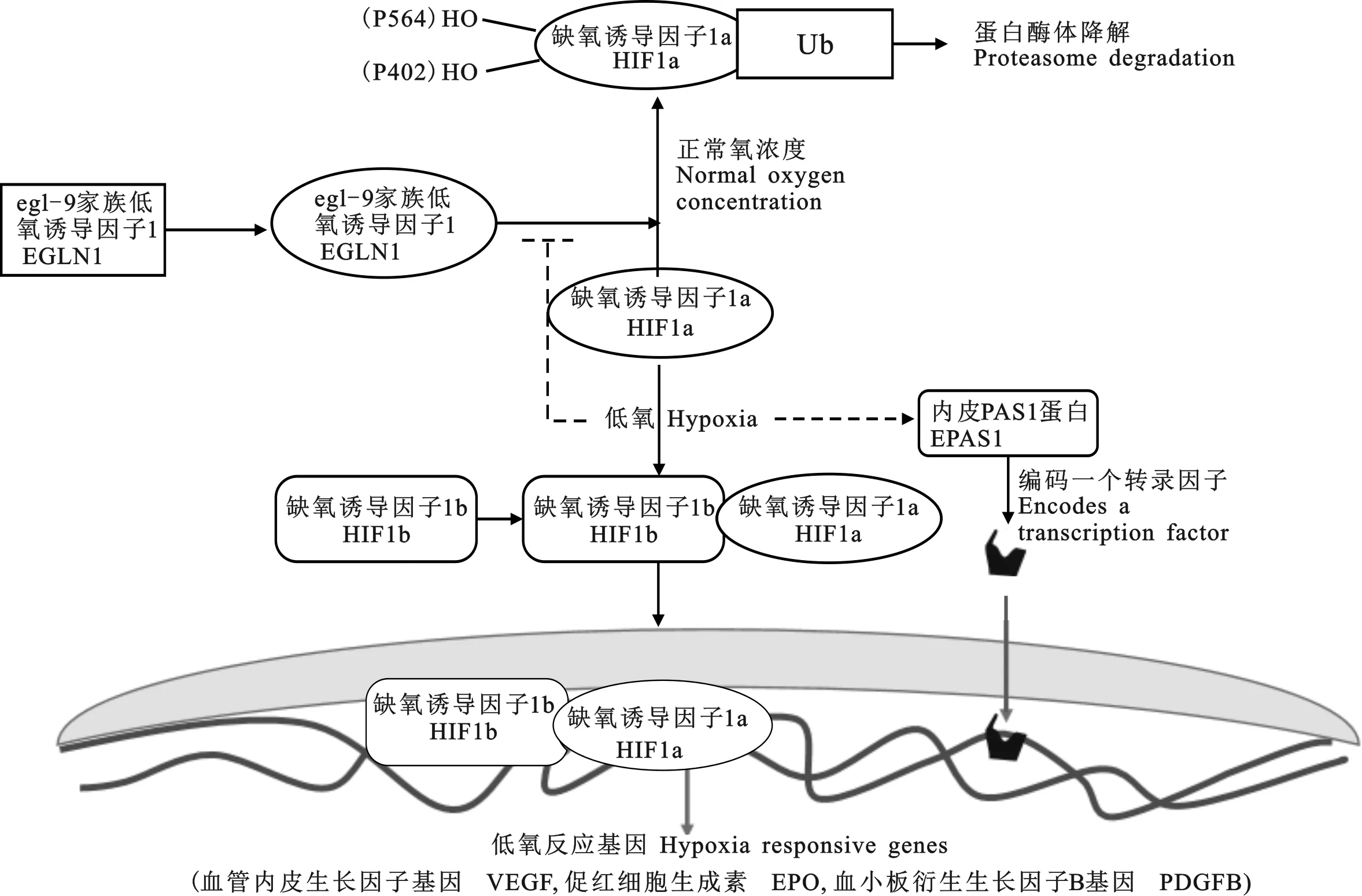
图2 氧气感应途径(改自文献[18])
1.2.3 藏羊与牦牛的高原适应性基因研究 上述两条通路在山羊与绵羊中具有高度的相似性。肖骁等通过对比检测藏山羊与乐至黑山羊的HIF-1α、VEGF165及其受体基因的表达量,发现藏山羊目的基因表达量明显低于乐至黑山羊,表现出明显的低氧环境适应性;Song等通过对位于330个不同地理区域的绒山羊外显子测序,获得了118 700个常染色体SNP,并挖掘了可能处于高海拔选择的339个基因,发现这339个基因中有极大一部分是与心血管发育系统的基因,其中最有代表性的是1基因,这一结果与藏羊中的研究相符合;Kumar等对5个中国羊品种的外显子进行测序,在高地与低地绵羊之间发现了27个SNP位点,发现3基因与西藏绒山羊的高原适应性具有密切关系。
Hu等研究了分布于整个青藏高原地区的986个藏羊样品的全基因组序列、SNP序列、线粒体DNA以及Y染色体在藏羊中的基因组变异情况,确定盘羊在氧气运输、感知和形态学相关通路相关基因等在藏羊中有渗入现象,且进一步确定了藏羊的迁移路线和时间,为深入了解早期藏区牧民和藏羊高原适应性以及人类在青藏地区定居历史的研究提供了参考。
从进化角度看,环境越极端,在相同时间段内,环境压力对于动物选择的强度越大。Wang等以藏羚羊和牦牛为研究对象,发现二者之间存在着细胞因子信号传导抑制因子4(suppressor of cytokine signaling 4,4)基因的趋同进化,4基因是对低氧适应相关调控因子HIF-1α具有调控作用的基因之一;除此之外,生活在高原与低海拔的同一物种表现出明显的趋异进化,如比较基因组学分析发现红尾沙蜥与低地区沙蜥在与低氧耐受、能量代谢以及紫外损失反应相关的基因中存在着非同义变化率。
从牦牛基因组与牛基因组的对比分析中可以看出,牦牛基因组的GC含量与普通牛相似,基因同源性较高;然而在对印度本地牛外周血单个核细胞的转录组研究中,高海拔地区牛的-1、1、等基因的表达明显更高,这可以用图2所示的氧感应途径解释;与此同时,EPO(促红细胞生成素)同样在低氧适应性中发挥重要的作用。缺氧条件下,EPO产生并增加,通过促使红细胞进入血液而降低机体对低氧的不利反应,不同地区的牦牛基因具有一定差异,但同源性仍然较高;Qiu等通过对比牦牛和普通牛的基因,鉴定出牦牛具有85个正选择基因,且这些基因主要集中在与高原适应性有关的低氧适应和能量代谢通路中;周大鹏等对藏獒肺组织进行研究时,同样发现了藏獒肺组织富含胶原纤维与弹性纤维,肺泡管的数量较多,肺泡隔内毛细血管丰富。
1.2.4 藏羊与牦牛高原适应性基因的挖掘及应用 为了尽可能地利用藏区动物基因发掘缺氧适应性相关基因,将其应用于临床医学研究中,Shi等对人类与生活在高原上的狗、马、猪、牛、绵羊、山羊进行分析,并将其与生活在低海拔地区的相应物种进行了比较,发现高原动物的1(m6A修饰的mRNA结合蛋白基因)是缺氧适应性基因,在缺氧耐受性和癌症发生方面都起着至关重要的作用,为了解癌症发生机制提供了新的思路;同时,研究人员发现,藏区的家养动物与野生动物均具有较长的寿命,藏区老人的长寿(>100岁)比率较平原老人高,且发现衰老相关基因在藏区人群与动物中都表现出快速进化,与平原相应物种具有显著差异。
为了进一步提高对低氧环境的适应能力,机体运氧、用氧能力较强也是藏区动物显著的生理特征。从血液学上分析,藏区动物表现出明显的红细胞(erythrocyte,RBC)和血红蛋白(hemoglobin,Hb)含量升高,同时,平均红细胞容积(mean corpuscular volume,MCV)和血细胞比容(hematocrit,HCT)较低,平均红细胞血红蛋白浓度(mean corpuscular-hemoglobin concentration,MCHC)较高,使得藏区动物在提高运氧能力的同时避免因RBC、Hb升高而造成的血液粘稠。
1.3 藏羊与牦牛低温适应性与强紫外适应性的研究进展
除低氧外,藏区环境还具有低温的特点。以青藏高原为例,年均气温常年维持在0 ℃以下,成为除低氧以外动植物生长的第二大逆性条件。对大多数动物来说,增加其绒毛、被毛厚度,增厚脂肪层,是其抵抗寒冷的重要手段;除此之外,藏区动物通过解偶联蛋白(uncoupling protein,UCP)的解偶联机制进行产热。UCP是位于线粒体内膜上的解偶联蛋白,可促进氧化呼吸链驱出的质子重新进入线粒体,从而减少三磷酸腺苷(adenosine triphosphate,ATP)合成的能量消耗,转而维持产热;目前,高原低温相关研究常常被锁定于能量代谢相关基因,包括重组人丙酮酸激酶(recombinant human PKLR protein,)、烯醇化酶3(enolase 3,3)、轴丝动力蛋白重链9(axin dynamin heavy chain 9,9)等,均已被研究人员在高原非人类哺乳动物中鉴定。
低温同时导致藏区饲草短缺、来源匮乏的现象。因此,藏羊等藏区家畜需要利用有限的饲草保证自身正常代谢。Ma等等对藏羚羊、藏野驴、藏绵羊的肠道微生物进行比较后发现,与生活在低海拔的绵羊相比,藏羚羊的肠道菌群中具有更高的瘤胃菌科(Ruminococcaceae)、梭菌目(Clostridiales)和拟杆菌门(Firmicutes)的富集,这些结果与宏基因组学分析共同反映出高海拔食草动物的肠道微生物组成相似,且功能相似,表明这些肠道菌群可能与高海拔适应性有关;此外,与小尾寒羊相比,在负能量平衡的情况下藏羊脂肪分解代谢较低但脂质合成能力却更强;藏羊瘤胃微生物中,分解纤维真菌属的比例较高,这或许与藏羊适于青藏放牧环境有关。同时,宏基因组测序结果显示藏羊具有大量和碳水化合物代谢相关的独特的菌群,说明藏羊肠道微生物在藏羊适应青藏高原环境过程中也发生演化。
藏区环境强烈的紫外辐射,驱使动物进化出了强紫外适应性的相关基因,藏区动物具有在强紫外线下特有的皮肤保护基因,或可作为临床治疗人类相关疾病的材料;除对皮肤的损伤以外,强紫外照射造成了藏区动物DNA修复相关基因的迅速进化——高辐射使得藏羚羊与DNA修复相关的基因表现出明显的正选择信号,这些基因的特殊表达使得藏羚羊具有强大的DNA修复能力,能够正常生存繁衍。
2 藏羊与牦牛生产性能的改良
2.1 藏羊的改良
藏羊是最广泛分布于青藏高原地区的粗毛羊品种,具有较强的自然气候条件适应能力,但同时也具有生长速度缓慢、个体成熟较晚、产毛量不足的缺点,如何改进藏羊、利用藏羊改良其他优良羊品种,是亟需解决的问题。
白萨福克羊具有体型大、生长发育快、产肉性能好的优点。正确合理地利用白萨福克羊的优点,使之成为优良的杂交亲本,对藏羊的遗传改良具有十分重要的意义。为探索白萨福克羊对低氧环境的适应性,将其转运至青藏高原并饲养,扩大其分布范围。周明亮等在高原过渡环境下对27只白萨福克羊的血液生理生化、生长发育等指标进行了比较分析,发现各指标均在正常范围内波动,其繁殖力与抵抗力都较强。这一结果为藏区引入白萨福克羊提供了理论支持,证明白萨福克羊与藏羊杂交改良的可行性;杨杜录等对杜泊羊、萨福克羊与青海藏羊一代杂交羔羊的生长发育情况进行了研究,发现杂交子代表现出较强的跟群放牧、耐粗饲等特点,较为完整地保留了藏羊在抗病、抗逆方面的优良特性,且3月龄与5月龄时的体重、体斜长、胸围等指标明显优于当地藏羊,有效地改良了藏羊生长发育慢、成熟晚的缺点。
2.2 牦牛的改良
牦牛广泛分布于青藏高原地区,具有优良的高原抗逆性,但由于其进化中自然选择比人工选择强烈,使得牦牛虽可适应逆性环境,但其生长速度、生产性能、繁殖性能都较其他牛种更差。使用中国黄牛杂交改良牦牛生产性状是一条改良牦牛的良好思路,二者的后代同时拥有亲本双方的优势,可大大改进牦牛产乳产肉性能。但黄牛与牦牛的后代——犏牛具有雄性不育的缺陷,给改良工作带来了困难。李贤等研究发现,具有调控精子减数分裂同源重组过程作用的DNA减数分裂重组酶1基因(disrupted meiotic cDNA,1)基因正是F1代雄性犏牛高度不育的关键所在;董丽艳等通过检测黄牛、牦牛、犏牛3者睾丸组织中细胞分裂周期基因2(cell division cycle gene 2,2)和细胞分裂周期25A(cell division cycle 25A,25)基因的表达,发现牦牛和黄牛组织中这2种基因的表达量明显高于犏牛,这些发现或可为阐明犏牛不育的分子机理提供理论依据,也为黄牛杂交改良牦牛提供新的思路。
3 展 望
以藏羊、牦牛为代表的藏区家畜,是中国青藏高原上的珍贵基因库。藏区家畜不仅具备优良的产绒、产毛性能,具有极大的改良空间,同时也是研究低氧性疾病、寻找低氧耐受基因的完美实验模型。就家畜饲养来说,目前已有大量关于将低地家畜引入西藏地区饲养的研究,如改良藏羊、藏牛、藏猪等;同时,各种藏区动物的生理解剖、血液、基因组分析为完善动物机体氧气运输机制、寻找低氧耐受机制提供了充足的研究材料,有利于从遗传学的角度理解高原疾病的发生,如1基因可作为未来低氧耐受性改造实验的关键候选基因。在后续的研究中,有必要重点研究这些基因的变异是否可以帮助动物耐受高原低氧环境,并设法改造基因,创造新的高原耐受物种,达到扩大藏羊、牦牛等家畜饲养范围的目的。
猜你喜欢
当代体育科技(2022年14期)2022-11-04
青海湖(2022年3期)2022-06-09
滇池(2022年5期)2022-04-30
汉语世界(The World of Chinese)(2022年1期)2022-02-25
诗潮(2018年3期)2018-03-26
现代交际(2016年20期)2017-02-15
热带农业科学(2016年10期)2016-12-12
山东体育学院学报(2014年6期)2015-04-07
环球时报(2012-01-29)2012-01-29
环球时报(2012-01-29)2012-01-29
