
太湖流域直湖港水系不同水质状况的河道水体对麦穗鱼卵孵化的影响
2022-05-25 07:16:00王金庆李伟忠高峰涛封佳丽刘长娥
淡水渔业 2022年3期
王金庆,李伟忠,高峰涛,封佳丽,席 丹,刘长娥
(1.潍坊学院生物与海洋学院/现代设施渔业研究院,山东潍坊 261061;2.上海市农业科学院,上海 201403)
随着太湖流域经济社会的快速发展,太湖流域的工农业废水和生活污水排放量持续增加,流域水质近30年来呈现不断下降的趋势。太湖水质综合评价为劣Ⅴ类,而氨氮(NH-N)为Ⅱ类,TP为Ⅳ类,TN为劣V类,化学需氧量(COD)为Ⅲ类。太湖流域水污染和富营养化的各种因素中,主要是面源污染的贡献。鱼类产卵、孵化、仔鱼、幼鱼至成鱼等生活史各阶段均具有特定的环境需求,对水环境理化性质变化比较敏感。很多外界环境因素都对鱼类群体的结构产生影响,例如河道坡岸硬质化、船舶通行和工农业氮磷排放,都会引起鱼类栖息地质量下降和鱼类物种多样性的降低。长江中下游地区,工农业生产活动对水生态系统的干扰同样是不断加剧,鱼类等水生生物栖息地被显著改变,对鱼类群落生态产生负面影响。龙延河的水体污染引起了营养盐含量升高、溶解氧和透明度降低,对多种鱼类具有不良影响。
鱼类是河道水生态系统的消费者,对于清理有机质、净化水质具有重要作用,鱼类的生物多样性决定了水域生态系统完整性和水生食物链稳定性。麦穗鱼()是太湖流域广泛分布的小型鱼类,产粘性卵,是研究水生栖息地质量、水质改善效果的良好的指示物种。麦穗鱼摄食水中的有机碎屑,进行废弃物循环利用和净化水质,是重要的清水型鱼类。麦穗鱼生存力强,资源量丰富,在内陆湖泊和河道中具有较高的生态优势度。麦穗鱼鱼苗在室外水泥池中可以顺利生长和存活,开展人工养殖和资源开发也具有很高的可行性;鱼体上部有新月形黑斑,具有观赏价值,喜食残饵、易于饲养,也可以在水族箱内作为功能性鱼类(“清道夫”)或原生态观赏鱼进行养殖。
本研究以麦穗鱼为试验对象,探讨直湖港河道不同富营养状况的水体对麦穗鱼的受精卵孵化、鱼苗成活的影响,分析光照、水体TN、TP、NH-N、COD、pH、溶解氧(DO)、电导率(EC)、氧化还原电位(ORP)等环境因子与水生类群密度变化的响应关系,评价不同水质参数和营养盐含量对于提高鱼苗孵化效率的重要性。本研究归纳了不同水体营养盐指标的生态作用,为开展水域生态修复工作提供参考资料和基础数据,也提高了河道治理过程中水生动物资源保护的针对性。
1 材料与方法
1.1 水体取样与测定
依据以前的水质监测结果,分别在重污染、中度污染和轻度污染水域采集水样。2011年6月从江苏省无锡市直湖港各级河道里取不同污染程度的水体,共设置4个样点(表1),取样点分别位于直湖港、朱家浜中部、人工湿地、龙延河西部(图1)。同时选取室内曝气自来水(ZLS)作为对照,进行麦穗鱼受精卵孵化实验。
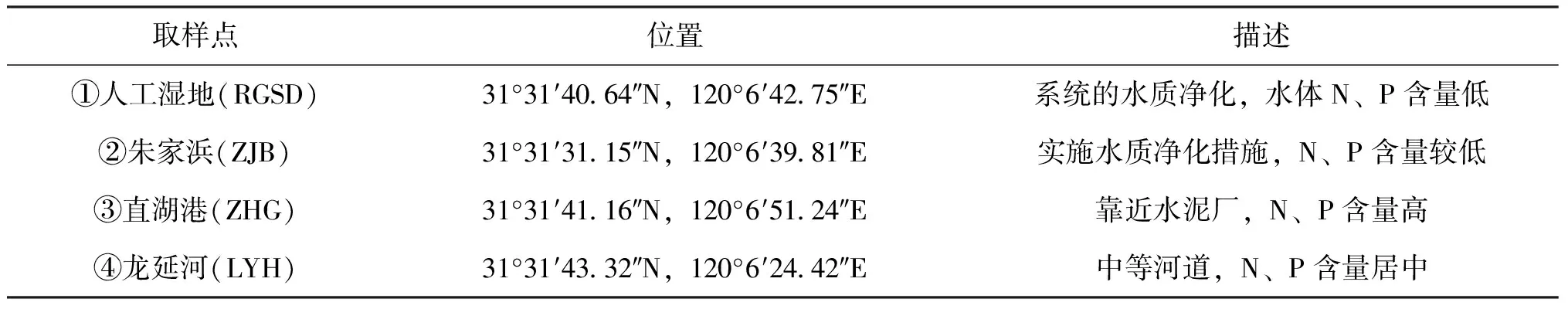
表1 水样采集地点坐标
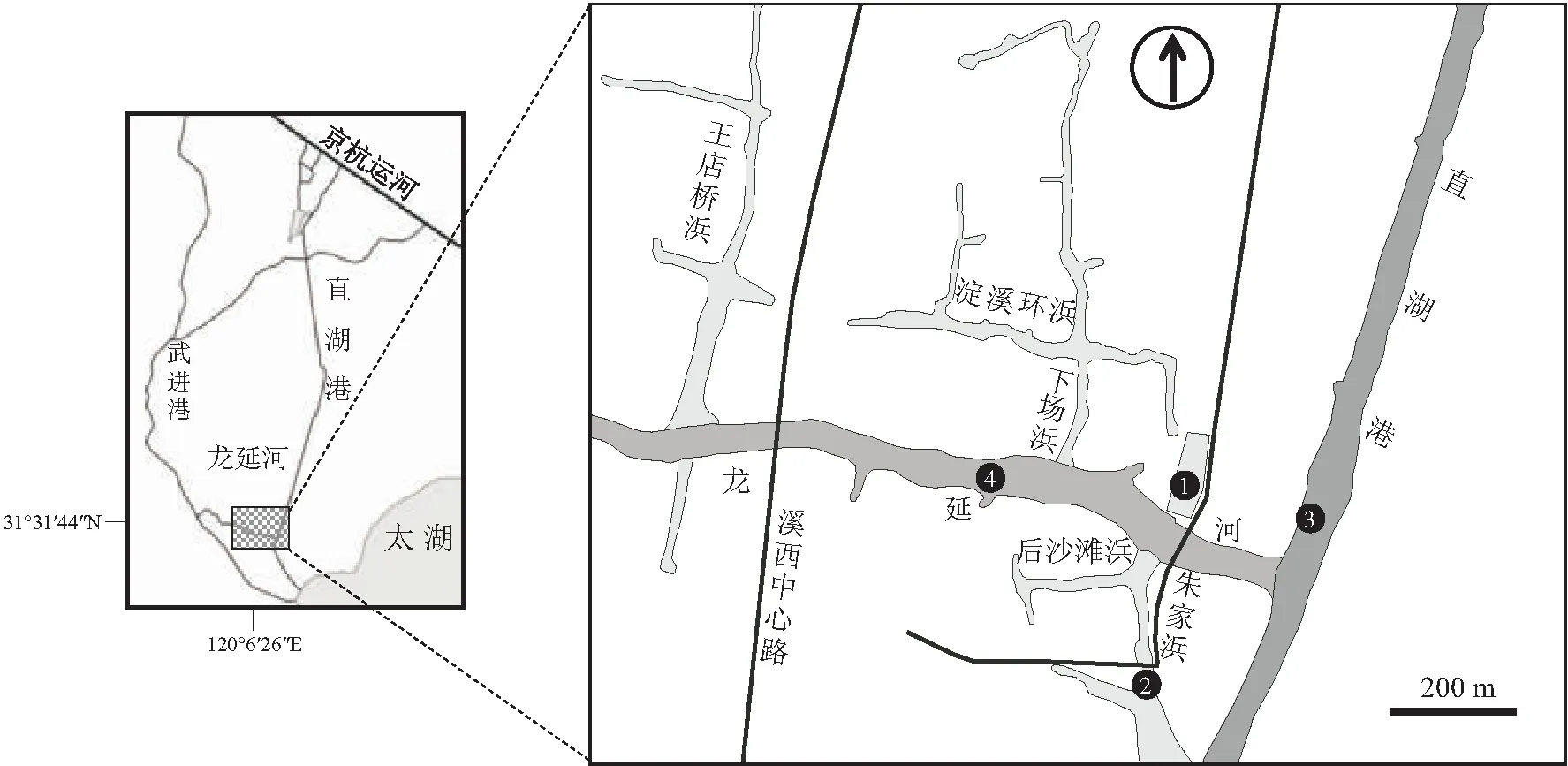
图1 研究区域和样点示意图
1.2 鱼卵的取样与孵化
来自4种不同水源的水样分别存放于4个25 L塑料桶中,当天运至实验室。鱼卵采自直湖港、龙延河及其分支河道,麦穗鱼在繁殖季节频繁在茭草()、水花生()、芦苇()等根茎上洒卵。鱼卵连同水草根茎(茭草茎、水花生根、芦苇茎等)一同被采集放入塑料瓶中并装入河水,水瓶上部钻孔并定期晃动用以增氧,运至实验室开展培养实验。水样分为RGSD、ZJB、ZHG、LYH、ZLS 5个处理,设置4个重复,倒入20个1 000 mL烧杯开展实验。每个烧杯内放入20粒卵,每5 d计数鱼苗的孵化数、存活数和死亡数,直至实验结束。实验期间定期补充蒸发损失的水分,每天用玻璃棒搅动增氧。计数鱼苗时间分别为6月28日(孵化第5 d)、7月3日(孵化第10 d)、7月8日(孵化第15 d)、7月13日(孵化第20 d)共4次。同时中午12:00记录光照、室温和水体理化指标。监测指标包括TN、TP、NH-N、COD、pH、DO、EC、ORP和光照(Light)。静置期间水体营养盐指标的变化动态参考文献[15]。
使用多参数水质分析仪(HI9828,Hanna,Italy)测定DO、pH、EC、ORP,室内光照采用照度计测定。TN的测定方法为碱性过硫酸钾氧化紫外分光光度法、TP为钼锑抗分光光度法、NH-N为纳氏试剂比色法、COD则采用高锰酸盐指数法测定,测定方法均按《水和废水监测分析方法》进行。
1.3 统计分析
各参数计算公式为:
孵化率=孵化个体数/受精卵个数×100%;
成活率=成活个体数/孵化个体数×100%。
采用Statistica 7.0进行统计分析,采用One-way ANOVA分析20 d实验期间不同水样的水质理化指标,以及鱼卵孵化率和鱼苗存活率的差异,分析了水生种类密度(鱼苗孵化率、轮虫密度、剑水蚤密度)与环境变量的相关性。实验数据均采用平均值±标准误(Means±SE)表示,采用Levene’s法进行方差齐性检验,Cochran C法进行正态性检验,当不满足齐性方差时,对百分比数据进行反正弦或平方根处理,采用ANOVA对实验结果进行方差分析,采用Tukey’s法进行多重比较,以<0.05为差异显著性标准。
使用Canoco 4.5软件(Microsoft,美国)对物种密度(鱼苗孵化率、轮虫密度、剑水蚤密度)与环境因子(光照、水体TN、TP、NH-N、COD、pH、DO、EC、ORP)间的关系进行冗余分析(RDA),并利用基于499次置换的Monte Carlo检验判断各环境因子与RDA排序轴的显著性。
2 结果
2.1 不同来源水体的水质状况变化
鱼卵孵化期间水体理化指标的变化如图2所示。实验期间的光照强度为室内无灯光时的自然光照,均在200~500 lx之间,各处理的光照情况基本一致。在实验的第5天至第15天的光照较强,其余时间光照较弱。
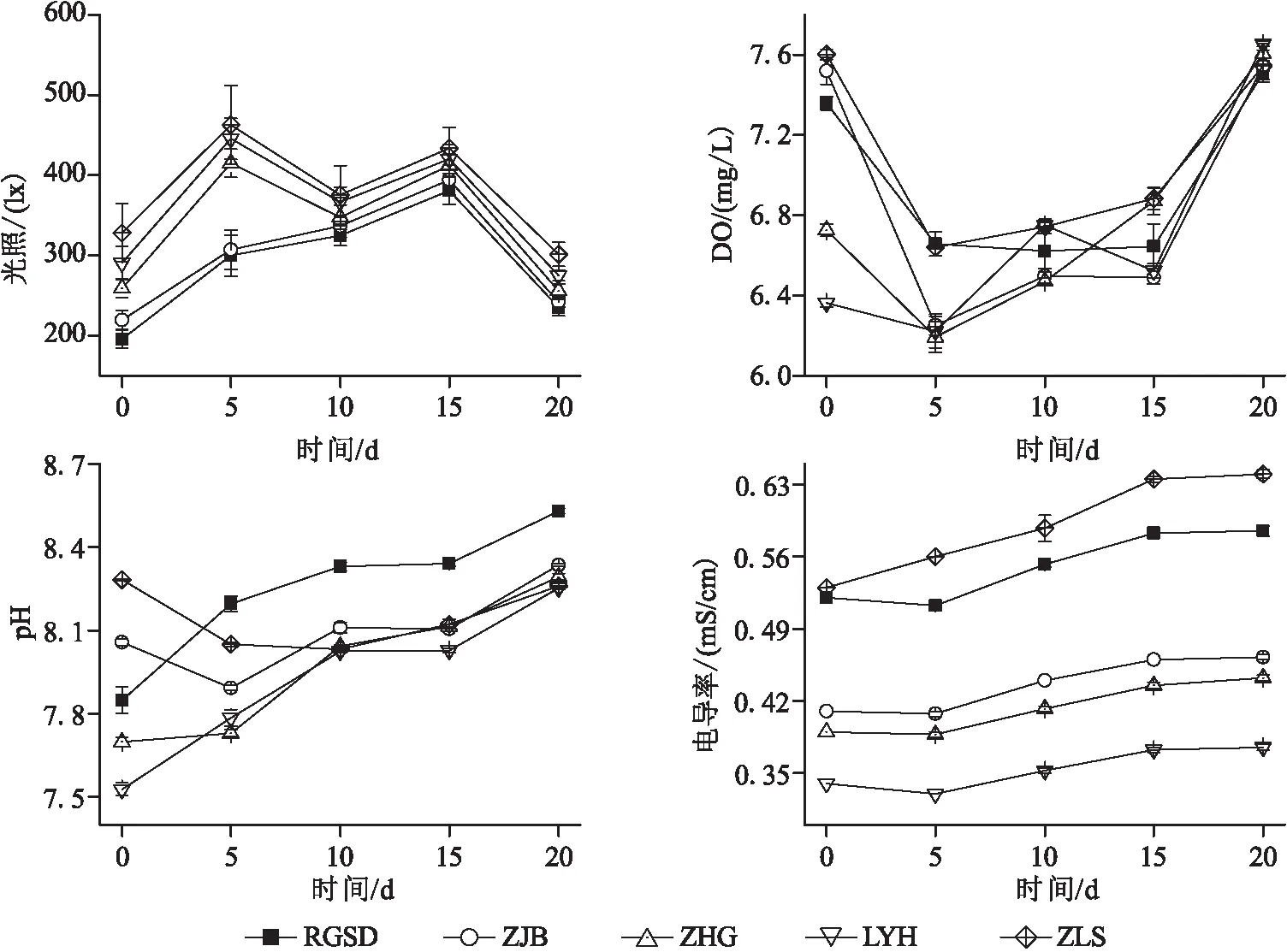
图2 鱼卵孵化期间环境和水体理化指标的时间变化
DO波动比较大,第0天,水体DO以ZLS最高,其次ZJB、RGSD,ZHG和LYH最低;第15天的ZLS和ZHG显著高于LYH和ZJB。曝气使得ZLS的DO明显增加,但在第5天至第15天期间DO含量降低,由于孵化出膜的鱼苗数量增加,耗氧率增大所致,鱼苗数量降低后DO含量在第20天显著回升。
实验期间水体pH和EC均逐渐增加,这与实验期间的水分蒸发有关,尽管在实验期间不断给烧杯内补充水分,水体的pH和EC仍然出现了增加。在0 d,pH从高到低排序依次是ZLS、ZJB、RGSD、ZHG、LYH;在5 d,排序变为RGSD、ZLS、ZJB、LYH=ZHG;在10 d,为RGSD、ZJB、ZHG=ZLS=LYH;在15 d,为RGSD>ZLS=ZHG=ZJB>LYH;在20 d,为RGSD>ZJB>ZHG=ZLS>LYH。实验期间RGSD的pH逐步上升并保持最高,ZLS的pH逐步降低,LYH保持最低。EC从高到低排序为ZLS、RGSD、ZJB、ZHG、LYH,EC高低顺序不随着取样时间改变。从0~20 d的EC逐步升高,尤其是15~20 d升高速度最快。
实验期间水体ORP的变化如图3所示。依据水体的氧化还原特征与周围环境状况,评估作为鱼苗繁殖与生长栖息地的水环境质量。ORP表示了水体的氧化能力,14 d时ORP从大到小依次是ZLS、LYH最高,其次ZHG,最低为ZJB和RGSD。后来由于鱼苗死亡耗氧,18 d时ORP在RGSD、ZJB、ZHG、LYH间无显著差异,ZLS显著最高。随着时间延长,其中LYH、ZHG、ZLS的ORP降低,而RGSD和ZHG水样ORP升高。本研究中,14 d时,ZLS、LYH和ZHG本身ORP值较高,LYH和ZHG河道宽和流速大,水体具有良好的水质自净和恢复潜力。LYH与ZLS在18 d时ORP出现显著降低。本研究中繁育鱼苗实验水体保持静置,水体有机物质含量高时氧气消耗增加,导致呈还原态有机物大量积累,ORP降低,有机物长碳链上的氮、磷、硫等被还原,最终产生氨气、甲烷和硫化氢等。ZLS有机质含量低,同时ZLS消毒用二氧化氯、漂白粉和次氯酸钠含量高,故水体ORP值持续较高,保持较高的氧化能力。

图3 水样ORP的时间变化
2.2 不同来源水体的麦穗鱼卵孵化率和水生种类密度的差异
从孵化率来看(图4),ZHG的鱼苗孵化率最高,其次是ZJB、LYH、ZLS,而以RGSD最低,水体营养盐含量高时鱼苗的孵化率相应提高。图5中可以明显看出,在第5天,出苗量最高的是ZHG水体培育的鱼卵,其次是ZJB、LYH、ZLS和RGSD。在第10天,出苗量高低顺序与第5天一致,也是ZHG最高;但在第15天,仍有鱼苗存活的是ZJB和ZLS。在孵化第5~10天时,苗数达到最大值,孵化数量从大到小依次是ZHG、ZJB、LYH、ZLS和RGSD。为评价水环境质量对鱼苗抗逆性的影响,培育期间未投喂饵料,在第15天时,在RGSD、ZHG、LYG水样内鱼苗均已死亡,而ZJB和ZLS培养鱼苗还有存活;在第20天时,所有水体内鱼苗均已死亡。

图4 实验期间各来源水体麦穗鱼卵最大孵化率比较
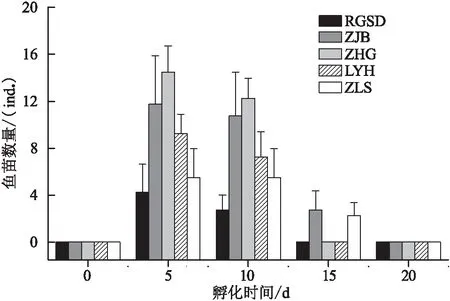
图5 孵出鱼苗数量的时间变化
在鱼苗孵化后期10~20天,鱼苗开始死亡,而浮游动物大量繁殖,在孵化鱼苗的水体中,浮游动物主要包括轮虫和剑水蚤两种;轮虫为萼花臂尾轮虫(),剑水蚤为广布种剑水蚤()。剑水蚤的大小为1.22 mm×0.36 mm,无节幼体为0.49 mm×0.17 mm,轮虫为0.44 mm×0.34 mm。图6表明,RGSD水体中轮虫数量明显高于其他来源水体,剑水蚤也有相似的现象;ZJB、ZHG、LYH水体中轮虫数量为0,剑水蚤数量较高;ZLS中轮虫数量略高,剑水蚤略低。在不同来源水体中剑水蚤的密度没有显著差异。非参数检验表明,轮虫数量存在显著差异,RGSD里轮虫数量最高,其次是ZLS,其他水源没有轮虫(Chi-Square=10.625)。

图6 不同来源水体浮游动物的密度
2.3 环境因子与水生种类密度的相关性
DCA分析显示坐标轴的最大梯度长度为1.86,意味着水生种类与环境因子之间的关系呈线性,因此选择RDA分析,RDA结果显示前两轴解释了66.6%的种类密度变化,前4轴共解释水生动物类群结构总变异的91.4%,环境因子有效区分了不同水源的水生类群密度。种类-环境关系的累积方差值第1、2轴达到88.7%,很好地拟合了种类和环境之间的相关性(表2)。

表2 针对鱼苗孵化率、轮虫和剑水蚤密度与环境因子相互关系的RDA分析
为了证明环境因子对水生种类解释量的显著性,对实验进行Monte Carlo显著性检验(499组排序),置换检验结果为第1轴=4.8,=0.03,所有轴检验=1.4,=0.24。在环境单因子对水生种类影响的重要性上,排在前6位的是TN、pH、EC、TP、COD、DO,分别解释了水生类群结构变异的22%、18%、15%、11%、11%、8%(表3,表4)。除了分类变量外,环境因子pH、EC、COD、DO与第1轴正相关性较高,RGSD样点与第1轴正相关性较高,与轮虫密度负相关性最高的指标为TN(=-0.54)、TP(=-0.32),而与轮虫正相关性较高的为pH(=0.49)、EC(=0.36)。TN、TP、光照与第1轴负相关性较高,ZHG样点主要分布于该方向。TP、NH-N与第2轴正相关性较高,LYH、ZJB样点与该轴正相关性较高。EC、DO、ORP与第2轴负相关性较高,ZLS样点主要分布于第2轴负方向,与剑水蚤密度正相关性较高为NH-N(=0.31),负相关性较高的指标为TN(=-0.28)、ORP(=-0.27)。与鱼苗孵化率正相关性较高的指标为TP(=0.40)、TN(=0.62)、NH-N(=0.17),而负相关性较高的为EC(=-0.47)、DO(=-0.42)、pH(=-0.47)、COD(=-0.24)(图7)。

表3 环境变量的显著性检验及其RDA各轴的相关系数
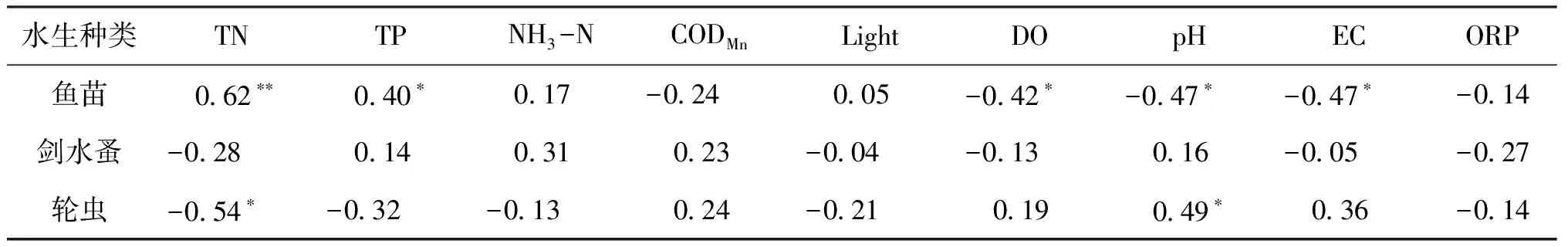
表4 水生种类密度与环境变量的相关性分析
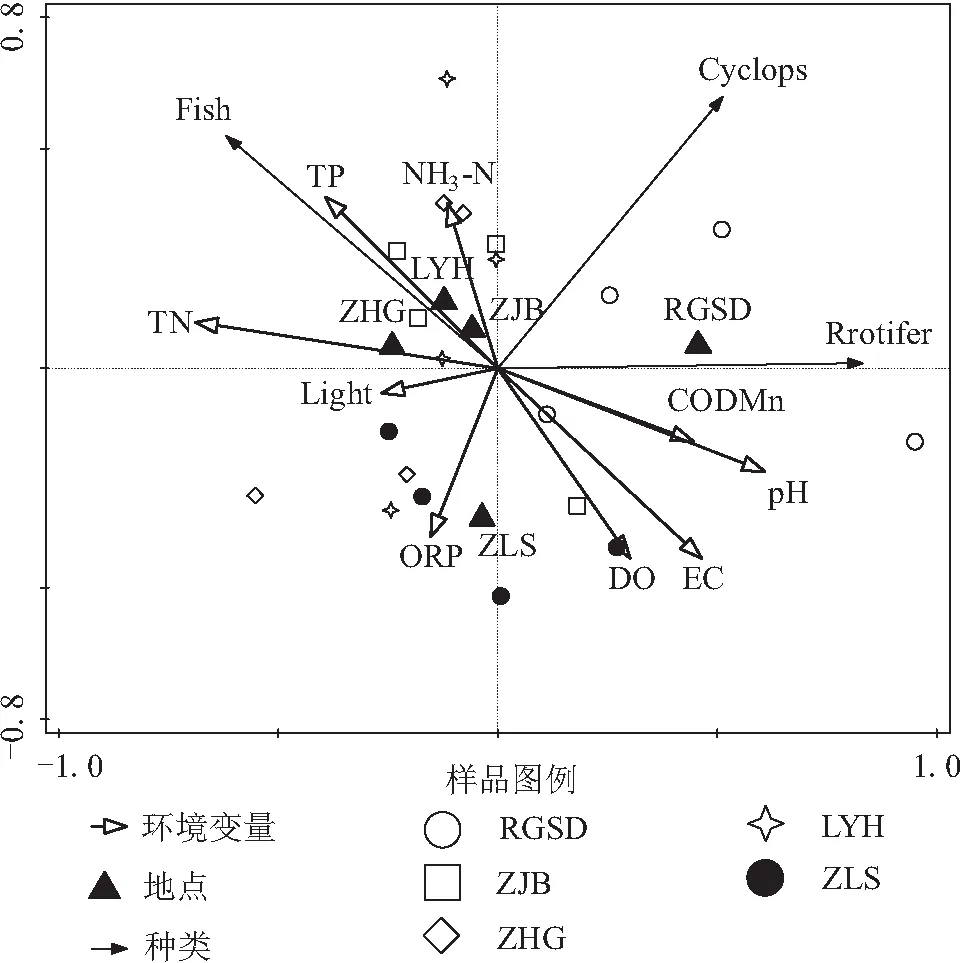
图7 鱼苗孵化率、轮虫、剑水蚤密度与光照和水质等环境指标的多变量RDA分析
3 讨论
3.1 不同来源水体的水质状况

水体TN含量从高到低依次是直湖港[(7.99±0.14)mg/L]、朱家浜[(6.77±0.14)mg/L]、龙延河[(6.15±0.15)mg/L]、自来水[(5.24±0.07)mg/L]、人工湿地[(4.28±0.12)mg/L]。水体NH-N含量从高到低依次是龙延河[(1.20±0.007)mg/L]、直湖港[(0.94±0.006)mg/L]、自来水[(0.66±0.006)mg/L]、人工湿地[(0.26±0.01)mg/L]、朱家浜[(0.16±0.002)mg/L]。水体TP含量从高到低依次是龙延河[(0.337±0.002)mg/L]、直湖港[(0.256±0.002)mg/L]、朱家浜[(0.158±0.003)mg/L]、人工湿地[(0.074±0.004)mg/L]、自来水[(0.017±0.002)mg/L]。氮、磷是构成生物体各组分的重要元素,是决定浮游生物生长的重要营养元素,对水体中的水生生物及水质状况具有重要影响。龙延河、直湖港作为通航河道,沿岸有厂房分布和工农业的污水排放,水体富营养化程度高,TN、TP、NH-N和COD等营养盐指标比较高;同样原因,直湖港水体培育的鱼苗孵化率也是最高的,轮虫和剑水蚤数量较低,一种解释是鱼苗对浮游动物的捕食作用引起的,另一种解释是浮游动物本身不适应富营养化的水体出现密度降低。人工湿地和朱家浜河道采用生态浮床、人工礁石等措施集中开展了水体生态修复和水质净化,有效削减了水体N、P,水体景观效果改善显著,然而COD仍然较高,人工湿地水体pH、COD、DO、EC较高,鱼苗孵化率最低,轮虫数量和剑水蚤数量达到最高。自来水由于水处理工艺特点,TP、COD、TN含量低而Cl含量高,导致EC、ORP比较高,具有较高的轮虫数量和较低的剑水蚤数量,同样不利于鱼苗的孵化和存活。
河道鱼类物种多样性受到多种环境因素的影响,往来船只对鱼类的觅食、生长、游动行为具有不利影响,河岸混凝土硬质化改造也对水域生态造成很大破坏,从而降低了栖息地质量和鱼类的多样性。东太湖的鱼类群落结构也处于中度干扰状态,鱼类个体小型化趋势明显,鱼类多样性水平较低,总磷含量、pH、DO含量等是影响东太湖鱼类群落的主要环境因子。平凡等预测北京市西南部界河拒马河2018~2060年河道内麦穗鱼适宜的栖息地面积将逐渐增加,且秋季栖息地面积明显增加。而在太湖部分支流河道存在岸带狭窄、硬质化严重、生境单一的问题,部分岸坡水生植物群落受到破坏,麦穗鱼适宜栖息地面积有一定下降,影响了水域生态稳定性,应适度恢复河岸挺水植物群落,逐步修复退化的水生态系统,才能有效促进鱼类群体的繁育和增殖。
3.2 影响鱼苗存活的主要因子
鱼卵的孵化和鱼苗的存活受到某些关键水质指标的显著影响。韩萃等报道Mg在大西洋鲑()早期发育阶段具重要影响,在养殖水体添加5 mg/L Mg显著提高鱼卵存活率和仔鱼开口率,添加8 mg/L的K损害鱼卵孵化。张家海等用循环水孵化法孵化斑点叉尾鮰()鱼卵,孵化率达到94.15%,显著高于常规流水孵化法的孵化率57.35%。斑马鱼(var)鱼卵的孵化与水温、DO等均有一定的关系。许秋瑾等报道沉水植物影响水质指标并间接影响斑马鱼鱼卵的孵化,沉水植物叶片附着生物可作为食物促进仔鱼的生长。在内陆河流中鱼类主要以鲤科淡水鱼类居多,鲤科小型鱼类具有很强的生命力,具有摄食有机碎屑并促进其分解循环的生态作用。DO是鱼苗存活重要的水质条件,本研究鱼苗孵出后显著消耗水体的DO,引起DO在5~15 d的明显降低,在15 d鱼苗死亡后DO又明显回升。直湖港和龙延河水体营养盐含量较高,营养分解消耗的DO相应较多,而朱家浜、人工湿地的营养盐含量较低,消耗DO的量较小,所以朱家浜、人工湿地的水体DO水平高于直湖港与龙延河。
直湖港和朱家浜水体的TN含量最高,与鱼苗孵化率的提高有很大关系;而其pH、EC、ORP、DO都是居中的,说明不是鱼卵孵化的决定因素。自来水的pH、EC、ORP、DO都是最高的,鱼苗的存活力最强,产生的浮游生物数量也最少,可能是被鱼苗摄食殆尽。范思林等发现宜昌鳅鮀()的鱼卵水平分布受到空间位置的影响,整体水平分布两岸高于中间。方刘等培育翘嘴鳜()鱼苗期间受到鱼苗规格增长和代谢旺盛的影响,TN和NH-N持续升高。本研究中环境因子对水生种类影响的重要性上,排在前6位的是TN、pH、EC、TP、COD、DO。RDA分析表明,与鱼苗孵化率正相关性较高的指标为TN(=0.62)、TP(=0.40)、NH-N(=0.17),而负相关性较高的为EC(=-0.47)、DO(=-0.42)、pH(=-0.47)、COD(=-0.24)。麦穗鱼苗孵化率最高的是直湖港和朱家浜水样,最低是人工湿地和自来水水样,而培养15 d后只剩下朱家浜和自来水存活有鱼苗,说明这两种水源培育的鱼苗具有较强的抗逆性。
3.3 不同水生类群的生态关系
小型鱼类是水域食物网的重要一环,与浮游动物、螺类、虾类共同消耗掉水体中的有机碎屑,保持水生食物链的多样性和稳定性,促进物质循环转化和能量流动。麦穗鱼栖息于水流速小或静水环境中,食性为杂食性,并摄食绿色藻类。刘飞等认为麦穗鱼更喜好浮游动物和摇蚊幼虫、水丝蚓等动物性食物,对红色有选择偏好。杨晓鸽等报道长麦穗鱼()的消化道形态结构与其食性相适应,口咽腔上皮可见球形味蕾分布,肠长比为0.82,口裂/吻长比约为0.25,口裂特征属于温和杂食性鱼类。赵晓丹等报道断线麦穗鱼()种群具有较高的生态位宽度,能够缓解捕食者带来的压力,全年摄食绿藻门、甲藻门、桡足类、枝角类、轮虫、原生动物和昆虫碎屑,而仅在春季可摄食扁卷螺,在夏季可摄食水熊,在秋季摄食水螨。
降低水体N、P可以提高水体的透明度、清洁度和水色等景观效果,但“水至清则无鱼”,本研究发现过低的N、P导致营养物质匮乏、食物链的中断和水生动物数量的大幅降低,不利于鱼苗的孵化存活;从适度保持小型鱼类资源丰度的角度,不能一味追求水体N、P的降低,藻类可以利用水体N、P营养盐,应适度保持水体营养浓度以维持藻类密度,从而增加小型鱼类的食物资源。
富含营养的水体会自发形成藻类,出现藻类到轮虫到桡足类的种群繁盛期。孙枭琼等报道营养盐和叶绿素a浓度会影响浮游动物摄食,从而影响其丰度数量和群落结构组成。孙晓梅等指出TP、NH-N、COD是浮游动物群落分布的主要影响因子,很多浮游动物优势种与TP、NH-N呈正相关性,说明营养盐类能显著影响浮游动物群落结构。在草鱼()养殖池塘轮虫丰度与TN和NH-N显著相关,枝角类和桡足类与环境因子存在一定的相关性。本研究有相似的结论,与轮虫密度负相关性最高的指标为TN(=-0.54)、TP(=-0.32),而与轮虫正相关性较高的指示为pH(=0.49)、EC(=0.36)。与剑水蚤密度正相关性较高的指示为NH-N(=0.31),负相关性较高的指标为TN(=-0.28)、ORP(=-0.27)。后续研究中也注意到,藻类吸收利用水体N、P无机营养盐,浮游动物采食藻类为食,螺蛳[如梨形环棱螺()、方形环棱螺()等]、河虾如中华小长臂虾()、秀丽白虾()利用多余的残饵和有机碎屑,水丝蚓[如中华颤蚓()、淡水单孔蚓(Hatai,1898)、苏氏尾鳃蚓()]、线虫、微生物等加速营养物质的分解转化,形成了完整的营养级关系。类似的,鱼蚌综合养殖试验中发现三角帆蚌()通过滤食水体中浮游植物、有机碎屑等悬浮颗粒增加水体透明度,降低水中TN和有机质含量。
鱼苗对浮游动物的捕食使鱼苗密度与轮虫、剑水蚤密度出现相反的变化。一种可能原因是鱼苗与轮虫、剑水蚤对水质条件需求的不同,造成各种类种群密度的差异;另一种可能的原因是,人工湿地水体孵化鱼苗数量较少,对轮虫和桡足类的捕食作用也较弱,因而轮虫数量增加。剑水蚤在自来水中没有出现,说明浮游动物的卵和幼体是由河道实验水体自身携带而来的,而不是通过卵块附着的水草根茎或叶鞘带入水体的。
相对于沿海地区和大江大湖,内陆河汊湾塘宽度一般较小,适宜湿生植物生长的坡岸浅水区域面积所占比例较高,同时淡水充足不存在咸水和海水盐碱对植物的生理限制,各类湿生植物高效利用底泥和水体营养盐呈现茂盛的生长态势。水生维管束植物提供有机碎屑成为水生动物适宜的食物资源,同时为鱼苗、螺蛳、昆虫等提供良好栖息场所和附着基质,并吸引鸟类、两栖类等类群,湿生植物和藻类对窄小河道水体生态完整性和多样性、食物网复杂性的影响更为重要。在河道水生态修复过程中,应充分认识河道水环境的独特性和优越性,充分恢复湿生植物群落作为优良的水生动物栖息地,保持高的生物多样性和水生食物网的稳定性,才能成功实现水体生态修复。
猜你喜欢
中国蚕业(2022年1期)2022-03-09 02:59:20
今日农业(2021年6期)2021-11-27 08:05:59
生物学通报(2021年9期)2021-07-01 03:24:44
辽宁化工(2021年5期)2021-06-03 05:15:50
——以“不同咖啡因饮料对水蚤心率的影响”为例
中学生物学(2021年2期)2021-04-30 12:04:06
蚕桑通报(2020年3期)2021-01-07 03:05:20
科学养鱼(2020年10期)2020-11-23 09:04:10
课外语文·中(2020年2期)2020-03-24 22:18:09
课外语文(2020年5期)2020-03-14 08:16:40
知识窗(2019年4期)2019-04-26 03:16:02
