
Comparative fish community structure among different types of enclosures of estuarine environments
2022-05-18 13:59SunwnHwngKeunHyungChoi
Aquaculture and Fisheries 2022年3期
Sunwn Hwng, Keun-Hyung Choi
aMarine Bio-Resources Research Unit, KIOST, Busan, 49111, Republic of Korea
bDepartment of Ocean Environmental Sciences, Chung-Nam National University, Daejeon, 34134, Republic of Korea
Keywords:
Fish community
Diking
Cumulative distribution
Estuary
Habitat connectivity
A B S T R A C T
This study explores the differences of fish assemblages in three macrotidal estuaries of Man-gyeong River Estuary(MRE), Geum River Estuary (GRE), and Han River Esturay (HRE) with similar ranges of seasonal water temperature that are diked with different levels of disconnection from freshwater discharge. The distribution of major species in the three estuaries could generally be grouped into three distinctive patterns based on their cumulative distribution with respect to the salinity gradient. The MRE was geographically closer to the GRE than to the HRE, but the fish community of the MRE more closely resembled that of the HRE. Increased freshness of the water inside the dike and the limit of coastal fish to the river may have led to this result. The results provide clues to how enclosure patterns of estuarine waters could affect fish communities over a long-term period. In addition, the information may provide guidance for how a fish community may return once environmental conditions are restored.
1.Introduction
Estuarine environments support high productivity and diversity of fauna (Cowley & Whit field, 2002; Day et al., 2013). They are important habitats for fish that provide nursery and foraging grounds, refuges, and migration routes (Day et al., 2013; Fujita et al., 2002; Hwang et al.,2011). Fish communities may consist of resident and migratory fish (i.e.,catadromous, anadromous, and amphidromous fish), including many juveniles, and re flect the environmental structure of the estuarine ecosystems (Elliott & Dewailly, 1995; Hwang et al., 2011).
Fish composition and assemblage structures are commonly affected by fluctuations in freshwater flow. In particular, variations in the scale,frequency, and seasonality of freshwater flows have profound effects,especially on fish species that require flood pulses for successful breeding or survival in early larval stages (Drinkwater & Frank, 1994;Ferguson et al., 2008). Connectivity between freshwater and marine ecosystems may be indicated (Hall et al., 2011), and alteration of the connectivity could lead to changes in the fish community over time.
Estuarine dikes and dams are probably the most significant alterations of estuarine habitat connectivity. They cut off migration passages,fragment habitats, alter flow patterns, and thus change the connectivity between freshwater and marine ecosystems (Hall et al., 2011). They can also restrict migration, reduce the abundance of diadromous fish species(Hwang et al., 2015; Kim et al., 2011), and affect fish communities overall.
Most studies on the effects of dams on fish communities have focused on the resulting changes in fish assemblages. Although this type of study provides direct information on the changes following estuarine reclamation, many estuaries may be lacking from data prior to reclamation.Fish monitoring is often not available at all or only begins once the reclamation is expected to be in a starting phase. Therefore, the beforeand-after effect of diking on fish community cannot be properly assessed. In such cases, a comparative hindsight analysis of fish communities of adjacent estuaries in similar environmental settings may provide insight or additional perspectives on the effects of reclamation.
The present study explores the differences of fish assemblages in three macrotidal estuaries of similar environmental settings on the West Coast of Korea. Qualitative and quantitative data were collected and compared to determine whether the composition and community structure of fish assemblages in the estuaries may re flect the different settings of the dikes, such as the location of the dike in the estuarine systems.
2.Materials and methods
2.1.Description of three river estuaries
The three macrotidal estuaries examined are all on the west coast of Korea (Fig. 1). The estuaries of the west coast of Korea present similar environmental settings with macrotidal waters of generally high turbidity. They also show similar patterns of seasonal temperature fluctuation (Lim et al., 2007, 2008). Almost all of the estuaries have been reclaimed, and no estuary is intact and free from human development. In the Han River to the north, at least 5 dams and 2 submerged dams have been constructed in the upper reach since 1943, which are mainly for flood control and supplying drinking water for the Seoul metropolitan area (Park, 2004).
The far lower reach of the Man-gyeong River Estuary (MRE) in the middle of the west coast was blocked with the completion of the Saeman-geum Dike (33 km in length) in 2006, which is the largest reclamation project ever in Korea (Lie et al., 2008). It has since allowed partial exchange of seawater only through gates and has radically changed the brackish water environment inside the dike as well as the seawater environment outside it (Lie et al., 2008). A dike of much smaller scale was constructed about a decade earlier in 1994 in the mouth area of the Geum River Estuary (GRE), which is located adjacent to the MRE. This dike blocked tidal propagation and the intrusion of seawater upstream (Lee et al., 1999). The Han River Estuary is generally considered human altered the least, with the dams being located near upper reach of the river for drinking water purposes.
2.2.Sampling of fish assemblages
Quantitative fish data were collected to determine the composition and structure of fish assemblages in the estuaries. Monthly fish samples were collected from the estuarine waters of HRE (2 stations, Fig. 1) in 2009 and 2010, as well as GRE and MRE (1 station each, Fig. 1) in 2003 and 2004. Sampling was carried out using stow nets tied to the stern of commercial vessels at depths of about 5 m from the surface level in the mainstream in the two estuaries (GRE and MRE). The 30-m long stow nets had an opened-mouth area of 125 m2(openings: 5 m vertical, 25 m horizontal) for the GRE net and 100 m2(openings: 5 m vertical, 20 m horizontal) for the MRE net with stretched mesh cod ends of 1–5 mm. In HRE, stow nets of 50-m length with an opened-mouth area of 30 m2(openings: 3 m vertical, 10 m horizontal) were used with stretched mesh cod ends of 25 mm (Hwang & Rhow, 2010). We accounted for potential differences in abundance due to different timing of sampling and the size of sampling gear by calculating cumulative fraction of the fish distribution (that is, normalizing the abundance data) and comparing them among the three estuaries.
2.3.Statistical analysis
We computed the cumulative distribution with respect to the salinity gradient of the relative abundance (percentage of total number of individuals) of major species in different estuarine habitats. Fish community structures were visualized using non-metric multidimensional scaling (NMDS) plots. ThenMDS was based on Bray–Curtis similarity estimates between pairs calculated from the log10-transformed abundance data (i.e., log10(abundance +1)) by using the metaMDS function in the Vegan Package within the R environment (Oksanen et al., 2007).Axes are rank order and thus unscaled. Juxtaposition but not distance between samples is an indication of similarity based on species composition of the samples.
3.Results
3.1.Hydrographic conditions
There are similar hydrographic conditions for both water temperature and salinity for the three estuaries (Fig. 2). The water temperature ranged from 6.4 ℃ in February to 25.4 ℃ in August in the GRE, from 4.3 ℃ in December to 31.2 C in July in the MRE, and from 3.0 ℃ in December to 27.0 ℃ in August in the HRE. The temperatures were noticeably higher in July (31.2 ℃) and August (30.4 ℃) in the MRE than in the GRE (Fig. 2a).

Fig. 2.Temporal variations in (a) surface water temperature and (b) salinity in the estuarine habitats of the Han River, Geum River, and Man-gyeong River on the mid-west coast of the Republic of Korea. HRE, GRE, and MRE represent the Han River Estuary, Geum River Estuary, and Man-gyeong River Estuary,respectively.
The monthly salinity ranged from 2.2 in September to 24.2 in October in the GRE and from 8.7 in June to 25.9 in November in the MRE. In the HRE, the salinity was highest at 23.4 in April and lowest at 1.4 in August (Fig. 2b). The overall pattern varied between the estuarine habitats. The sharp decline in September (2.2) in the GRE was probably due to the discharge of freshwater following the opening of water gates due to heavy rainfall (Fig. 2b). The higher summer water temperature associated with low salinity in the MRE may indicate the discharge of freshwater of higher water temperature.
Another broad survey of physical properties on the Korean coastal waters made in a recent year (year 2015) also showed that the distribution of water temperature and salinity are similar among the three river estuaries (Fig. 3). The winter temperature difference may be<1 ℃ across the three estuaries, and salinity is also within a few although salinity fluctuate more due to river runoff (Fig. 3).

Fig. 3.Distribution pattern of seawater properties around the west and south coasts of Korea: a) Temperature distribution at 1.5 m seawater depth in a winter day, b)Salinity distribution at 1.5 m depth in a winter day.
3.2.Fish composition
The major fish species caught during the survey is shown in Table 1,which shows some distinctive patterns. Japanese anchovy (Engraulis japonicus) was only dominant in the GRE, ranking the first, while dotted gizzard shad (Konosirus punctatus) was dominant only in the MRE,ranking the first. Estuary tail fin anchovy (Coilia nasus) and big-eyed herring (Sardinella zunasi) were abundant in all three estuaries. Both big head croaker (Collichthys lucidus) and Osbeck’s grenadier anchovy(Coilia mystus) were the two most abundant species in the HRE but not in the top 10 species in the other two estuaries (Table 1).
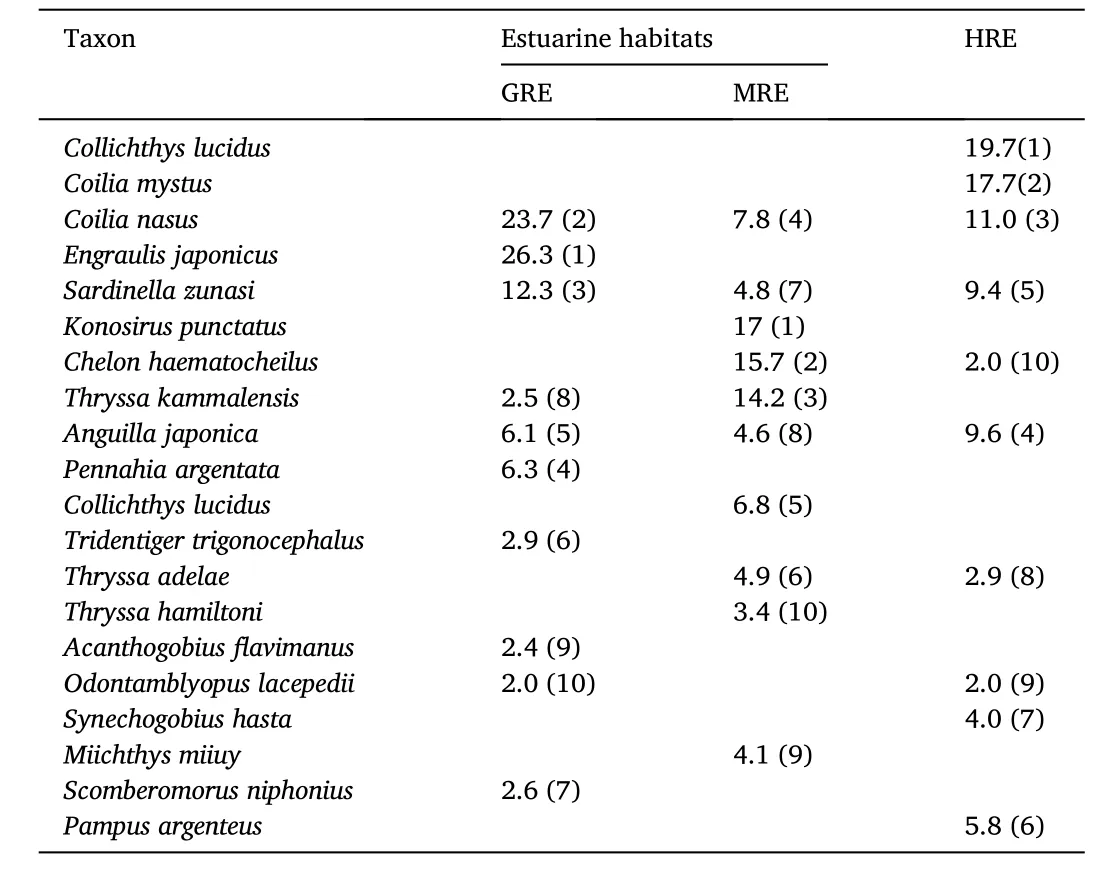
Table 1Ten most abundant fish species for each river estuary, with percent contribution and the rank in parenthesis GRE: Geum River Estuary, MRE: Man-gyung River Estuary, HRE: Han River Estuary.
3.3.Fish distribution patterns with respect to salinity gradient
The distribution patterns of major species in the three estuaries could generally be divided into three distinctive groups based on their cumulative distribution with respect to the salinity gradient (Fig. 4). First,some species showed a much higher mean percentage at lower salinity in the HRE than the other two estuaries (Fig. 4a–d), including the four speciesSardinella zunasi,Thryssa kammalensis, Coilia mystus, andJohnius grypotus. In particular,S. zunasicomprised>90% of fish abundance at salinity<5 (Fig. 4a).
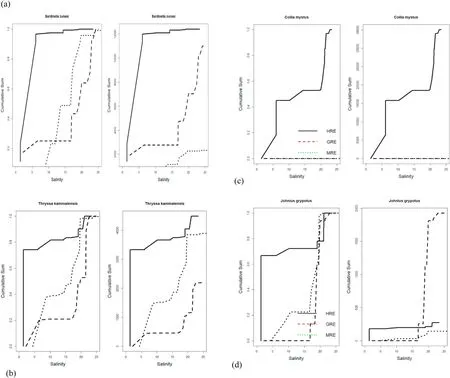
Fig. 4.Cumulative distribution with respect to salinity gradient of major fish for the three estuarine environments. The y-axis represents the cumulative fraction of the abundance, which should be 1 (that is, 100% of the abundance) at the end.
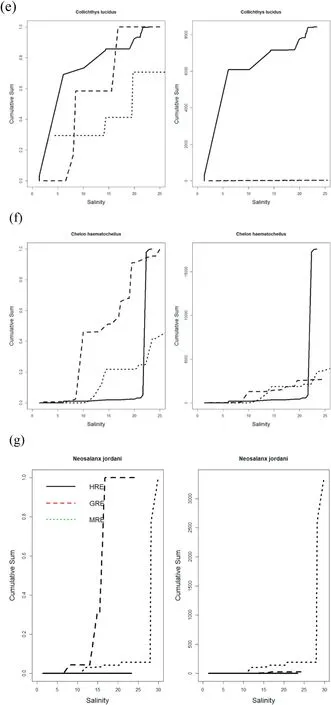
Fig. 4. (continued).
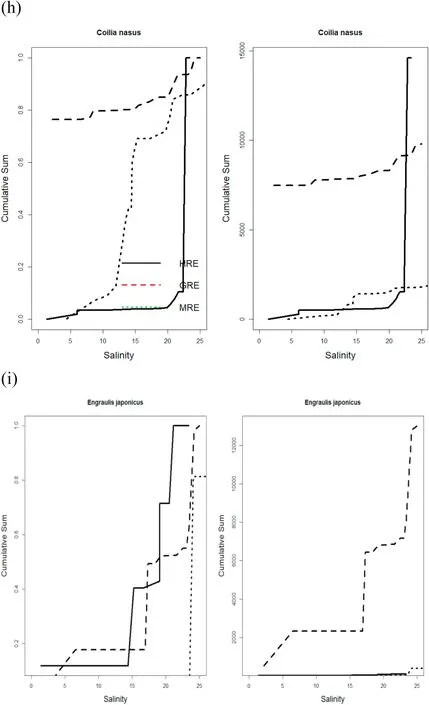
Fig. 4. (continued).

Fig. 4. (continued).
The second group of major fishes included species that had median abundance at lower salinity in the GRE than the other two estuaries.These fishes includedCollichthys lucidus,Chelon haematocheilus, Neosalanx jordani, andCoilia nasus(Fig. 4e–h). The third group of major fishes comprised species that showed more or less similar distribution patterns with respect to the salinity gradient (Fig. 4i–k). These includedEngraulis japonicus,Cynoglossus joyneri, andKonosirus punctatus. These fishes occurred in midranges of salinity or narrower ranges in salinity for the median abundance with respect to the salinity gradient compared with the other groups of fish. Among the major fishes,Pennahia argentatais probably the only fish showing median abundance at lower salinity at the MRE than the other two estuaries (Fig. 4l).
3.4.Regional similarity based on fish assemblage structure
Fish assemblages from each estuarine habitat based on a non-metric multidimensional scaling (NMDS) analysis showed a pattern of continuity from the MRE community on one side and the HRE community on the other side, with the GRE community situated in between the two communities (Fig. 5). However, both GRE and MRE are geographically closer to each other than they are to the HRE (Fig. 1). Therefore, the NMDS results may indicate other effects on the fish community other than the effects associated with the geographic location. Such effects may include the way in which the estuaries have been disconnected from freshwater discharge.

Fig. 5.Nonmetric multidimensional scaling (nMDS) ordination plots displaying samples by habitat. The nMDS was based on Bray–Curtis similarity estimates between pairs calculated from the log10-transformed abundance data. Axes are rank order and thus unscaled. Juxtaposition but not distance between samples is an indication of similarity based on species composition of the samples. GRE,MRE, and HRE represent the Geum River Estuary, Man-gyeong River Estuary,and Han River Estuary, respectively. The solid circles represent all the samples taken from all stations from the three estuaries during the study period.
4.Discussion
The three river estuaries were of similar hydrographic conditions,especially for water temperature (Figs. 2 and 3), which has significant effects on fish distribution via thermal limit. Salinity is also similar(Figs. 2 and 3) although it would be more prone to change due to fresh water discharge. The West Coast of Korea is a small area spanning a few hundred kilometers on the north-south direction (Fig. 1). Warm water current during winter is restricted to the southern coastal waters (Fig. 3),with the West Coast is pretty much isolated from the warm current effects. Water mass exchange between the Yellow Sea and the South Sea of Korea is very limited (Naimie et al., 2001). Thus, the West Coast encompassing the three estuaries can be considered a single marine ecoregion (Spalding et al., 2007) that is physically distinct from the other ecoregions surrounding Korea (Fig. 3).
We used fish catch data collected from different years and types of cod end bag for the HRE and GRE/MRE, which can potentially alter the results, especially the fish composition. However, these differences would not affect the cumulative distribution of fishes along the salinity gradient (Fig. 4), that is, spatial distribution. We calculated the cumulative distribution based on the fraction (that is, percent data), not on the abundance (i.e., the y-axis). Since the same sampling gear was used throughout the entire sampling period for each estuary, this would not have affected the pattern of the cumulative distribution based on fractional data (Fig. 4). The spatial distribution would have been driven primarily by the access to the river as indicated by the abrupt changes in the distribution along the salinity (Fig. 4). For instance, bothSardinella zunasi, a sardine, andThryssa kammalensis, an anchovy, are dominant small fishes around Korean coastal waters (Hwang & Rhow, 2010; Jeong et al., 2015), with their larvae adaptable to a wide range of temperatures and salinities (Ying et al., 2011). The distributions of both fish populations collected in the MRE and GRE shifted toward the lower salinity(i.e., the center of mass) compared to those collected in the HRE (Fig. 4).They may able to thrive even at lower salinity, but simply can not move to the upper reach of the estuaries at both the MRE and GRE.
Meanwhile, the fish composition that may well have changed over the time and the difference of sampling gear could have affected the fish composition and subsequently ordination of the community among the three habitats (Fig. 5). However, this difference also may have limited effects on the ordination results. An independent study by Lee et al.(2007) using a small trawl with a cod-end bag of 16 mm in mesh size shows thatKonosirus punctatuswas the most abundant fish during the survey conducted at the MRE in year 2006–2007 (Lee et al., 2007). In a separate study, Park et al. (2013) used stow nets of different lengths(30–50m) with 7–40 mm in mesh size of the cod end bag during a survey in 2012 in the MRE. They observed thatThryssa kammalensisandKonosirus punctatuswere the two most abundant fish both species contributing 16% and 13%, respectively, to the total fish catch (Park et al., 2013). These observation by independent studies are consistent with our earlier study made in 2003 that dotted gizzard shad (Konosirus punctatus) andThryssa kammalensisare the two dominant species in the MRE, ranking the first and third, respectively (Table 1). The HRE results of fish composition were also similar to another independent study (Oh et al., 2012) conducted during a similar period of 2010 at more sampling stations (a total of 4 stations compared to 2 stations in our study, Fig. 1).They showed that Korean anchovy (Coilia nasus), and Japanese grenadier anchovy (Coilia mystus) were the two most dominant species in the HRE, which again is consistent with our result in the region despite the sampling of smaller stations (2 sites, Fig. 1). The potential abundance variation among the different years and sampling gear would be alleviated by log10transformation of the abundance data for the multivariate analysis (Fig. 5).
In estuarine ecosystems, salinity has been recognized as an important factor for the spatial structure of fish assemblages and is considered to control the abundance of dominant species (Henderson, 1988; Selleslagh et al., 2009). The environmental data suggest that the three macrotidal estuaries still maintain estuarine characteristics and have a large monthly fluctuation of salinity that is influenced by freshwater discharge. Except for some strictly marine fishes, the availability of freshwater can drastically increase the habitat of some estuarine fishes(Fig. 4a–c). Their habitat can expand to almost freshwater environments when available with median salinity<3 (Fig. 4a–c) as these fishes are tolerant to wide range of salinity. Therefore, the fish community variation among the three estuaries (Fig. 5) is mostly likely due to the diking,affecting the limit of the expansion of marine fishes toward the lower saline region and the limit of the expansion of fresh water fishes toward the marine waters. Freshwater fish species in the MRE comprised more than 50% of the fish community, compared with<20% before the dike construction (Hwang et al., 2017). Meanwhile, the number of brackish fish species declined by more than 30% after the dike construction (Kim et al., 2011).
One big contrast in the fish composition among the three estuaries was that the two freshwater fishes of big head croaker (Collichthys lucidus) and Osbeck’s grenadier anchovy (Coilia mystus) were the two most abundant species in the HRE but not even in the top 10 species in the other two estuaries (Table 1). The two freshwater fishes are generally caught during large fresh water discharge only in the GRE (Hwang et al.,2005). Historically they used to be abundant in the GRE and MRE, but largely gone in the lower estuary and just reported in reduced numbers in the river of freshwater inside the dikes (Han & An, 2013; Hur et al.,2009). Increased freshness of the water inside the dike of MRE facilitated the proliferation of freshwater species while limiting the coastal fish to move into the estuary within the dike. Both effects combined may have resulted in a fish community that is more similar to that of the HRE,where the access to freshwater is generally unrestricted.
5.Conclusion
In conclusion, the results of the present study provide clues to how the enclosure of estuarine waters could affect fish communities over a long-term period. In addition, this information may provide guidance on how a fish community may return once environmental conditions are restored.
CRediT authorship contribution statement
Sunwan Hwang: Conceptualization, Methodology, Data curation,Data gathering. Keun-Hyung Choi: Formal analysis, Data analysis,Writing – original draft.
Acknowledgments
This study was partly supported by Chung-nam National University to K–H. Choi and by project PG50850 of the Korea Institute of Ocean Science and Technology (KIOST) to S.W. Hwang.
 Aquaculture and Fisheries2022年3期
Aquaculture and Fisheries2022年3期
- Aquaculture and Fisheries的其它文章
- Anthropogenic temperature fluctuations and their effect on aquaculture: A comprehensive review
- Recent advances in application of moving bed bioreactors for wastewater treatment from recirculating aquaculture systems: A review
- Characterization and expression analysis of gonad specific igf3 in the medaka ovary
- Liver DNA methylation and transcriptome between 1- and 3-year-old grass carp
- Screening and validation of reference genes for qPCR analysis in gonads and embryos of Takifugu bimaculatus
- Assessment of genetic diversity, detection of strain-specific single nucleotide polymorphisms and identification of the Bangladesh and Vietnam strain of Channa striata by PCR-RFLP analysis of the mitochondrial COI gene fragment