
外源亚精胺对低温胁迫下烟草种子萌发及幼苗生理特性的影响
2022-05-09 02:37魏晓凯何佶弦俞世康张启莉顾会战雷强喻保华李俊举景延秋
江苏农业科学 2022年7期
魏晓凯 何佶弦 俞世康 张启莉 顾会战 雷强 喻保华 李俊举 景延秋





摘要:为揭示外源亚精胺对低温胁迫下烟草种子萌发率及幼苗生长发育的调控机制,以云烟87为供试品种,设置不同浓度(200、400、600、800 μmol/L)亚精胺对烟草种子进行浸泡处理,探究低温胁迫下外源亚精胺浸种对烟草种子萌发率、幼苗抗氧化酶活性、渗透调节物质及膜质过氧化物含量的影响。结果表明,低温胁迫下烟草种子萌发率降低,幼苗的生长发育受到抑制,200、400 μmol/L亚精胺处理对低温胁迫的缓解作用有限,而600 μmol/L 亚精胺处理能够有效提高烟草种子萌发率,增强烟草幼苗抗氧化酶(SOD、POD、CAT)活性和净光合速率,增加渗透调节物质含量,降低活性氧自由基积累和丙二醛(MDA)含量,从而缓解低温胁迫对烟草幼苗造成的损害。
关键词:亚精胺;低温胁迫;烟草幼苗;缓解;种子萌发率
中图分类号:S572.01 文献标志码: A
文章编号:1002-1302(2022)07-0094-06
收稿日期:2021-07-10
基金项目:中国烟草总公司四川省公司项目(编号:2019410102000453);四川省烟草公司广元市公司科技攻关项目(编号:202051080024063)。
作者简介:魏晓凯(1994—),男,河南漯河人,硕士研究生,研究方向为烟草栽培,E-mail:1351262779@qq.com;共同第一作者:何佶弦(1988—),男,四川广元人,硕士研究生,研究方向为烟草栽培,E-mail:w1351262779@163.com。
通信作者:景延秋,博士,教授,研究方向为烟草化学。E-mail:jinglaoshi202105@ 163.com。
烟草作为一种喜温经济作物,原产于热带和亚热带,对温度变化较为敏感[1-2]。烟草生长发育最适温度为25~28 ℃,当温度低于12 ℃时,叶片光合速率会受到抑制,生长发育变缓,甚至停止生长,当温度高于35 ℃时,烟苗徒长,壮苗率低,也可能造成烧苗现象[3]。张嘉雯等研究发现,低温胁迫时烟草幼苗的抗氧化酶活性和渗透调节物质含量均显著升高,并产生大量的活性氧,造成植株生理代谢紊乱,生长受到抑制[4]。我国南方地区普遍存在因气温波动较大而出现“倒春寒”等低温伤害,不利于壮苗培育,导致幼苗成活率降低,缓苗期延长,烟叶产量和质量受到影响。如何缓解低温胁迫对烟株的危害已成为人们亟待解决的问题。
多胺是一种广泛存在于植物体内并具有逆境胁迫响应的低分子脂肪族含氮碱,主要包含腐胺、精胺和亚精胺,对植物生长发育及分化具有重要调节作用,其中亚精胺因其多聚阳离子特性调节作用效果尤为显著,它可以与质膜上的蛋白质等大分子结合,降低膜的流动性,维持植物体内酸碱平衡,同时亚精胺也可以作为信号分子参与激素的代谢[5-7]。近年来,越来越多研究表明,亚精胺对于缓解植物逆境胁迫具有显著的作用,如高温胁迫、硝酸钙胁迫、盐胁迫、干旱胁迫以及重金属胁迫等[8]。李书涵等研究发现,外源亚精胺能够显著增加生菜叶绿素含量,增强光合能力和抗氧化酶活性[9]。周晨楠等研究表明,外源亚精胺能够显著降低硝酸钙胁迫下番茄幼苗的丙二醛(MDA)含量和超氧阴离子(O- 2·)产生速率,进而减轻对幼苗的伤害[10]。赵东晓等研究表明,盐胁迫下外源亚精胺可作为信号分子诱导植物细胞中逆境蛋白等相关基因的表达,还能够通过激活脯氨酸合成的限速酶和其他关键酶活性,促进脯氨酸的积累,进而增强植物抗盐胁迫能力[11]。目前,关于亚精胺对植物的抗逆作用已成为人们关注的焦点,在许多农作物上均有研究,但对于低温胁迫下烟草生理机制却鲜有报道。鉴于此,本研究以云烟87为供试材料,通过设置不同浓度亚精胺浸种处理来探索低温胁迫下外源亚精胺对烟草种子萌发率和幼苗生理特性的影响,以期为烟草抗低温栽培提供理论依据。
1 材料与方法
1.1 试验材料
供试烟草品种为云烟87,由四川省烟草公司广元市公司提供;外源亚精胺(纯度为98%)购自山东福瑞达生物科技有限公司。
1.2 试验方法
试验共设置6个处理:CK,蒸馏水浸种;T1,低温15 ℃+蒸馏水浸种;T2,低温 15 ℃+200 μmol/L亚精胺浸种;T3,低温15 ℃+400 μmol/L亚精胺浸种;T4,低温15 ℃+600 μmol/L亚精胺浸种;T5,低温15 ℃+800 μmol/L亚精胺浸种。
试验于2020年11月在河南农业大学人工气候箱(RXZ-1000)中进行。每个处理选取50粒大小均匀、籽粒饱满的种子,按照上述试验处理在20 ℃黑暗条件下浸种24 h,然后用蒸馏水冲洗后放入盛有Hoagland营养液、pH值为5.7±0.2并垫有2层滤纸的培养皿上,每个处理3次重复。培养条件:光照周期为15 h/d,温度为(15±1) ℃,光照度为 4 000 lx。
点种后1~6 d,每天统计烟草种子萌发数量,并计算发芽率。待烟草幼苗长至5叶1心时,取从上到下第3张叶片进行各项指标的测定。
1.3 测定指标与方法
1.3.1 种子萌发率测定
萌发率=萌发种子数/种子总数×100%。
1.3.2 光合特性指标和叶绿素含量测定
使用便携式光合测定仪(Li-6400,美国LI-COR公司)于09:00—11:00测定各处理叶片的净光合速率( P n)、气孔导度( G s)、胞间CO 2浓度( C i)和蒸腾速率( T r);叶绿素含量测定参照李合生的方法[12]。
1.3.3 抗氧化酶活性测定
超氧化物歧化酶(SOD)、过氧化物酶(POD)和過氧化氢酶(CAT)的活性采用北京索莱宝科技有限公司生产的检测试剂盒测定。
1.3.4 根系活力和渗透调节物质含量测定
根系活力采用2,3,5-三苯基氯化四氮唑(TTC)法[13]测定,可溶性蛋白含量采用考马斯亮蓝G-250染色法[14]测定,脯氨酸含量采用水合茚三酮法[15]测定。
1.3.5 MDA含量和 O- 2·产生速率测定
MDA含量采用北京索莱宝科技有限公司生产的检测试剂盒测定;O- 2·产生速率采用苏州科铭生物技术有限公司生产的检测试剂盒测定。
1.4 数据处理
运用SPSS 17.0以及Excel 2016进行数据分析整理,采用最小显著差异法比较不同处理在0.05水平上的差异显著性。
2 结果与分析
2.1 外源亚精胺对低温胁迫下烟草种子萌发的影响
由图1可知,与正常萌发的烟草种子相比,低温处理的烟草种子萌发速率和萌发率均大幅降低。低温胁迫下外源亚精胺浸种影响了烟草种子的萌发率,且不同浓度亚精胺处理对种子的萌发影响不同。其中,点种后3 d,浓度为600、800 μmol/L的亚精胺处理烟草萌芽速率最快,均达到5.56%;其次为200、400 μmol/L亚精胺浓度处理,萌芽率分别为4.44%、2.22%。在点种后6 d,浓度为600 μmol/L的亚精胺处理烟草萌发率最高,达到92.22%,其次是800、400、200 μmol/L亚精胺处理。与单独低温处理(T1)相比,不同浓度的亚精胺处理,烟草种子萌发率分别提高了10.45%、14.92%、23.89%、2090%,说明适宜浓度的亚精胺浸种能够促进低温下烟草种子的萌发。
2.2 外源亚精胺对低温胁迫下烟草幼苗光合特性的影响
由表1可知,与CK相比,低温胁迫下烟草幼苗叶绿素含量明显降低,降幅为90.83%。与单独低温胁迫相比,不同浓度亚精胺处理能够不同程度提高低温胁迫下烟草幼苗叶片叶绿素含量,且随着亚精胺浓度的增加,叶绿素含量呈先升高后降低的趋势。其中600、800 μmol/L亚精胺处理时烟草幼苗叶绿素含量增幅较大,分别提高了174.07%、17284%,说明低温胁迫下外源亚精胺能够显著提高烟草幼苗叶绿素含量,促进净光合速率。从表1中还可以看出,低温胁迫下烟草幼苗的净光合速率( P n)、气孔导度( G s)和蒸腾速率( T r)均显著降低,而胞间CO 2浓度( C i)显著提高。与CK相比,低温胁迫下烟草幼苗的 P n、 G s和 T r分别显著降低了5194%、68.97%和53.10%,而 C i显著增加了8345%。低温胁迫下随着亚精胺浓度的增加烟草幼苗叶片 T r呈增加趋势,而 P n和 G s呈现先升高后降低的趋势,并且亚精胺浓度为600 μmol/L时效果最佳,该浓度下烟草幼苗叶片的 P n、 G s和 T r分别比单独低温胁迫时显著提高了113.03%、13333%、78.79%, C i显著降低了30.59%。说明外源亚精胺浸种可以明显增强低温胁迫下烟草幼苗的光合作用。
2.3 外源亚精胺对低温胁迫下烟草幼苗抗氧化酶活性的影响
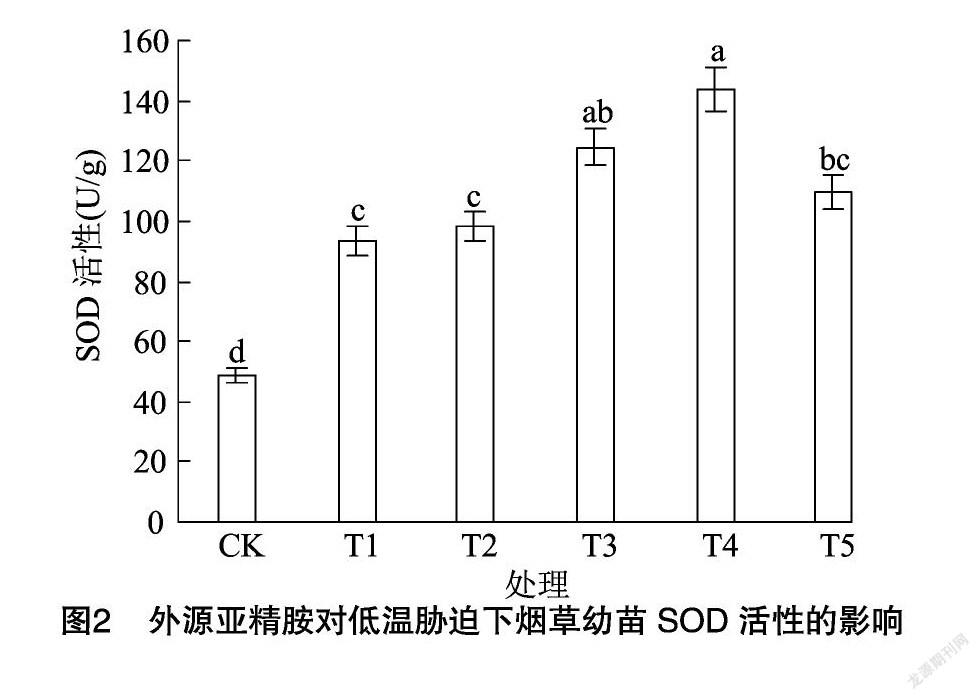
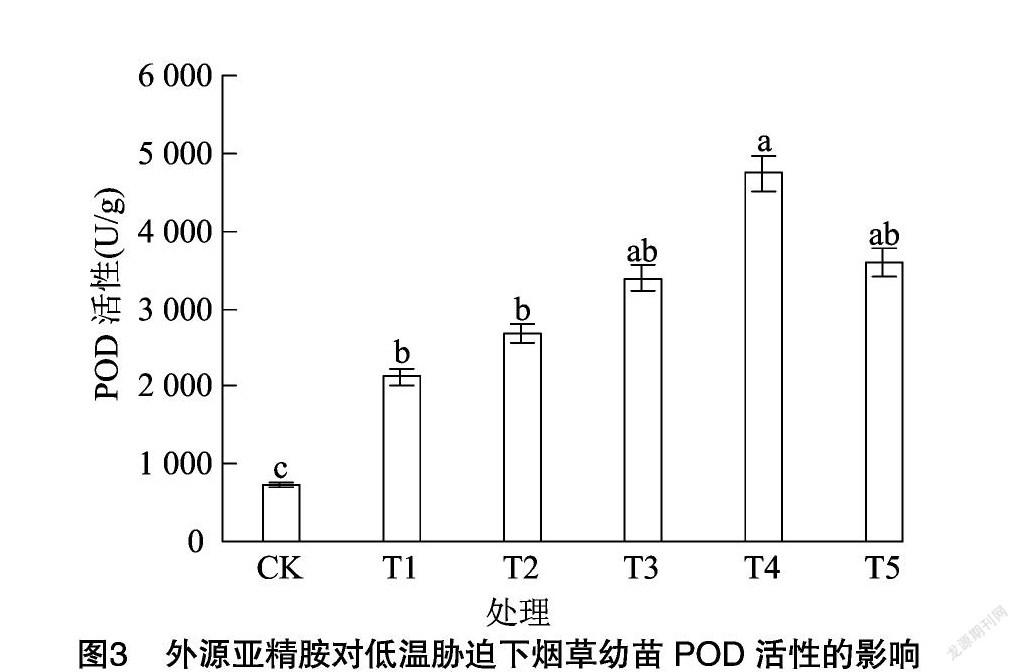
由图2、图3、图4可知,与CK相比,低温胁迫下烟草幼苗抗氧化酶(SOD、POD、CAT)活性分别显著提高了91.31%、189.00%、244.24%。与单独低温处理相比,不同浓度亚精胺浸种对烟草幼苗抗氧化酶活性影响不同,其中600 μmol/L亚精胺处理效果最为明显,抗氧化酶活性分别提高了53.86%、123.18%、25.00%,但随着亚精胺浓度的继续增加,烟草幼苗叶片抗氧化酶活性呈降低趋势,说明适宜浓度的亚精胺浸种可以显著提高抗氧化酶活性,增强抗低温能力。
2.4 外源亚精胺对低温胁迫下烟草幼苗根系活力的影响
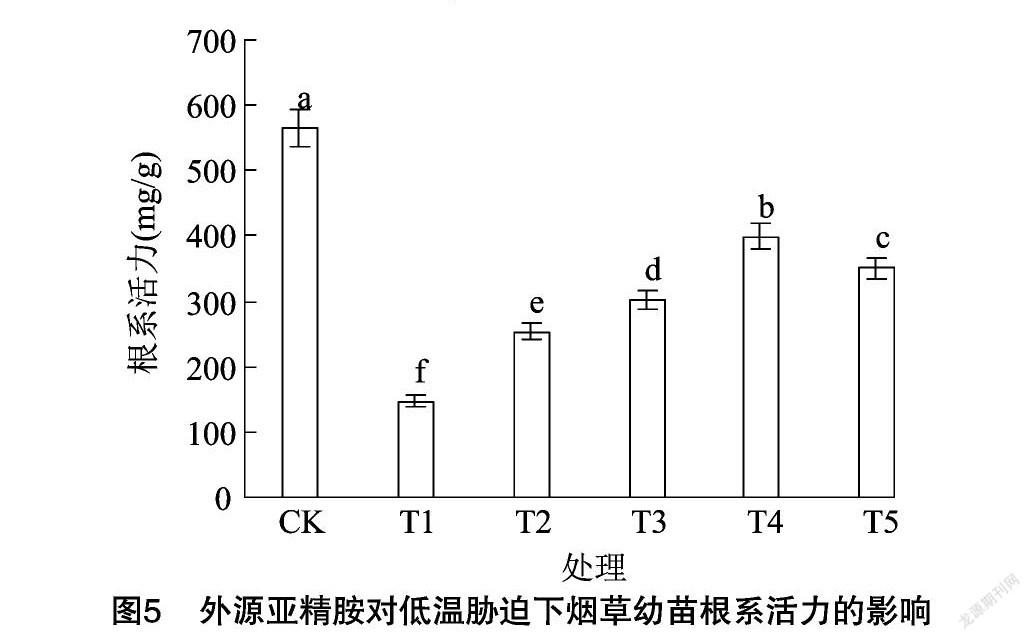
由图5可知,与CK相比,低温胁迫下烟草幼苗的根系活力显著降低了73.86%。与T1处理相比,低温胁迫下外源亚精胺浸种明显增强了烟草幼苗的根系活力,随着亚精胺浓度的增加,根系活力呈先增加后降低的趋势,T2、T3、T4、T5处理的根系活力较单独低温处理分别提高了71.86%、104.35%、171.13%、137.52%,且均达到显著水平,说明外源亚精胺浸种可以有效降低低温胁迫对烟草幼苗根系活力的损害,尤以600 μmol/L亚精胺处理效果最佳。
2.5 外源亚精胺对低温胁迫下烟草幼苗渗透调节物质含量的影响
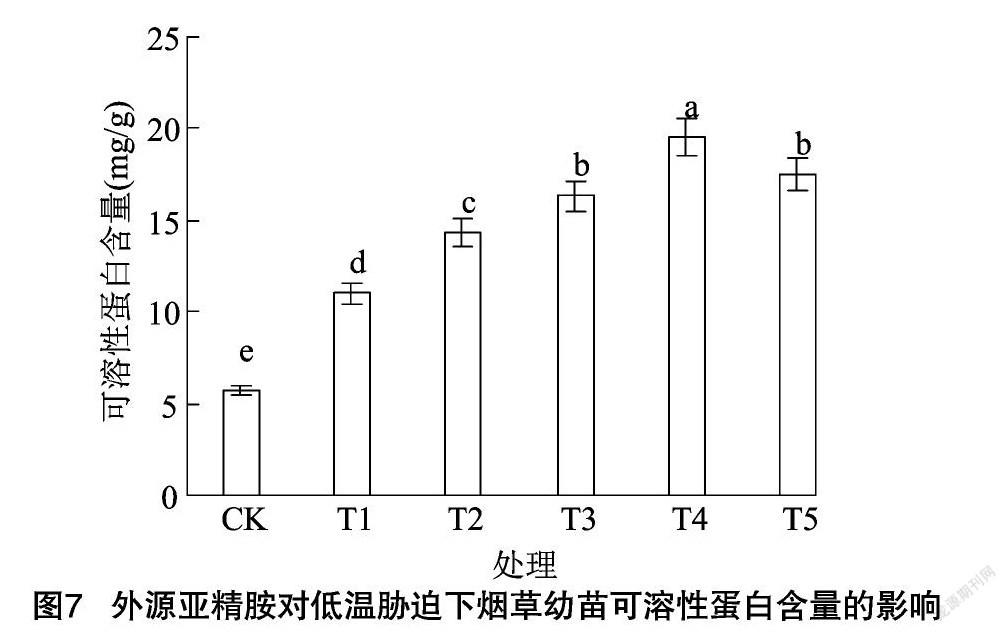
由图6、图7可知,与CK相比,低温胁迫下烟草幼苗叶片可溶性蛋白和脯氨酸含量显著提高,分别提高了93.00%、86.04%。随着外源亚精胺浓度的增加,可溶性蛋白和脯氨酸含量均呈先升高后降低的变化趋势,与单独低温处理相比,施用200、400、600、800 μmol/L亚精胺,可溶性蛋白含量分别提高了30.04%、48.46%、77.72%、59.26%,脯氨酸含量分別提高了21.11%、66.97%、11234%、6421%,且均存在显著差异,说明外源亚精胺浸种能够通过增加烟草幼苗叶片内渗透调节物质含量,增强抗低温能力。
2.6 外源亚精胺对低温胁迫下烟草幼苗MDA含量和 O- 2·产生速率的影响
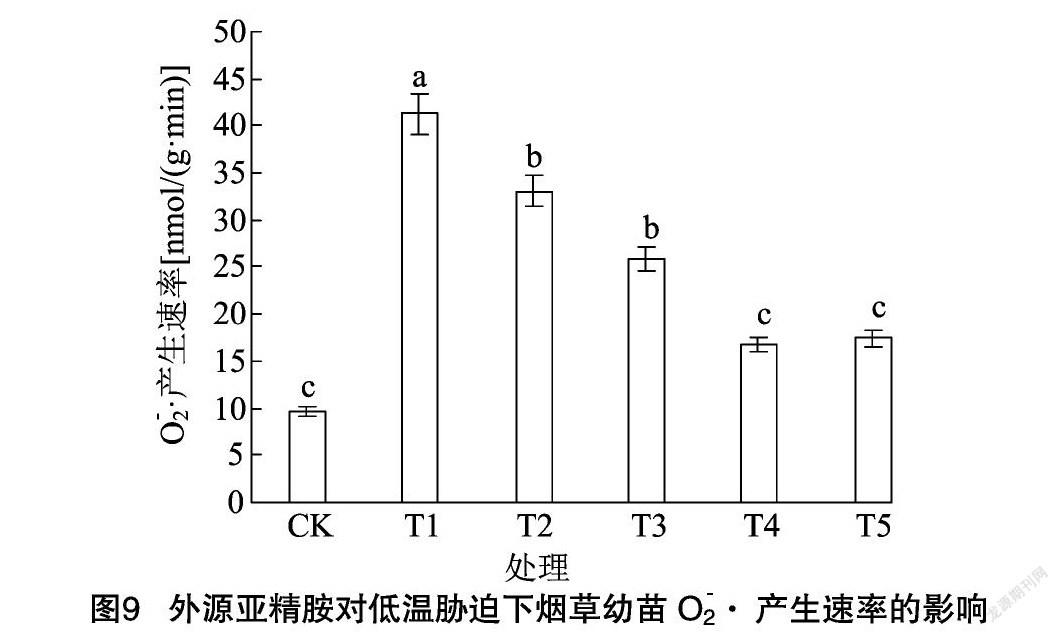
由图8、图9可知,低温胁迫下烟草幼苗中的MDA含量和 O- 2·产生速率显著高于CK,与CK相比分别提高了567.58%、330.58%。低温胁迫下经过亚精胺浸种的烟草幼苗叶片MDA含量和 O- 2·产生速率均显著降低。与低温处理相比,T2、T3、T4和T5处理的烟草幼苗MDA含量分别降低了35.75%、44.00%、55.76%、47.51%,O- 2·产生速率分别降低了19.78%、37.45%、59.30%、57.84%,说明外源亚精胺浸种能够有效减少MDA和 O- 2·的产生和积累,缓解低温胁迫对烟草幼苗造成的毒害,其中以浓度为600 μmol/L时的作用效果最为显著。
3 讨论与结论
种子发芽率可作为植物耐低温的重要指标。低温胁迫时种子萌发吸胀阶段的相对吸水量和吸水速率显著降低,种子活力也受到抑制,进而影响种子的萌发速率和萌芽率[15-17]。本研究结果表明,低温胁迫降低了烟草种子的萌发速率和萌芽率,这是由于低温胁迫导致种子相对吸水量和吸水速率大幅降低,种子活力和代谢能力减弱,萌芽受到抑制。已有研究表明,外源亚精胺能够提高植物种子的发芽率、发芽势和发芽指数[18]。本研究也发现,低温胁迫时外源亚精胺浸种能有效提高烟草种子的发芽率,尤以浓度为600 μmol/L时效果最佳,这与上述研究结果一致,说明低温胁迫下外源亚精胺能够通过增强烟草种子的活力,提高吸水能力,来维持细胞代谢平衡,进而提高种子发芽率。
光合作用是植物体内有机物合成的主要方式,直接反映了植物生理代谢的持续能力[20-21]。叶绿素作为植物主要的光合色素和功能物质,其含量直接反映了植物的光合能力[22-24]。大量研究表明,低温胁迫会导致植物叶片气孔导度降低,光合色素合成受阻,光能利用率降低,光合作用減弱,严重影响了植物的生长发育[25]。已有研究表明,亚精胺内带电荷的氨基和亚氨基能与蛋白质、核酸以及带电荷磷脂共同作用,来维持叶片类囊体膜结构的稳定,抑制光合色素的降解,减轻光化学反应受到的抑制[26]。本研究结果表明,低温胁迫下烟草幼苗的光合作用受到抑制,这主要是由叶片气孔导度降低、光合色素合成受阻造成的。在本研究中,外源亚精胺浸种能够明显提高烟草幼苗叶片的 P n、 G s、 T r和叶绿素含量,降低 C i,这与张永平等的研究结果[27]一致,这可能是因为外源亚精胺能够维持低温胁迫下烟草幼苗叶片类囊体膜结构的稳定,促进了光合色素的合成,保护了光反应系统,提高了碳同化能力,从而提高了低温胁迫下烟草幼苗的光合能力。
脯氨酸和可溶性蛋白都是植物主要的渗透调节物质,植物在正常生长时,渗透调节物质含量处于较低水平,而逆境胁迫时其含量会大量积累,以此来应对细胞渗透势的升高,进而增强植物的逆境适应能力[28]。姜秀梅研究发现,脯氨酸可以通过线粒体电子传递链,诱导植物体内抗氧化酶、泛素和保护蛋白等保护性物质活性或含量增加,同时启动抗低温代谢途径,从而缓解植物遭受的低温伤害[29]。已有研究表明,低温胁迫下外源亚精胺能够显著提高植物渗透调节物质含量[30]。本研究结果也表明,低温胁迫下烟草幼苗的渗透调节物质含量显著增加,而外源亚精胺浸种后,烟草幼苗渗透调节物质含量进一步显著增加,说明外源亚精胺能够通过增加植物体内渗透调节物质含量来降低细胞渗透势,维持细胞渗透平衡,进而保持细胞的完整,增强植物对低温胁迫的适应能力,这与上述研究结果一致。
低温胁迫会降低植物细胞膜流动性,引起细胞内活性氧清除系统紊乱,导致活性氧自由基大量积累,加剧脂质过氧化反应,其中过氧化产物MDA含量是反映膜损伤程度和植物抗逆能力的重要指标[31-32]。贺嘉豪等研究表明,逆境胁迫下烟草幼苗的脂质过氧化程度与 O- 2·产生速率和MDA含量呈正相关关系[33-34]。本研究结果表明,低温胁迫下烟草幼苗的 O- 2·产生速率和MDA含量显著增加,原因可能是低温胁迫使细胞内活性氧的产生和清除失衡,引起 O- 2·大量积累,加剧脂质过氧化反应,造成MDA含量大量积累。已有研究表明,低温胁迫下外源亚精胺能够显著提高植物对活性氧自由基的清除能力,减轻脂质过氧化损害。本研究结果也表明,低温胁迫下外源亚精胺浸种显著降低了 O- 2·产生速率和MDA含量,这是因为外源亚精胺可以诱导抗氧化酶活性提高,增强了植物对活性氧自由基的清除能力,减轻了脂质过氧化程度。
低温胁迫下烟草幼苗生长发育受到抑制,而外源亚精胺浸种能够有效缓解低温胁迫对烟草幼苗的抑制程度,尤以浓度为600 μmol/L时作用效果最佳。低温胁迫下外源亚精胺浸种能够诱导烟草幼苗抗过氧化酶活性提高,增加渗透调节物质含量,降低 O- 2·产生速率和MDA含量,缓解光合系统的损害,提高叶绿素含量,促进烟株的生长发育,进而提高烟草幼苗的抗低温能力。
参考文献:
[1]杨利云,段胜智,李军营,等. 不同温度对烟草生长发育及光合作用的影响[J]. 西北植物学报,2017,37(2):330-338.
[2]侯 爽,陈锦芬,刘溶荣,等. 外源水杨酸对烟草幼苗低温胁迫的缓解效应[J]. 湖南农业大学学报(自然科学版),2020,46(1):14-20.
[3]崔 翠,李君可,周清元,等. 温度胁迫下烤烟幼苗的生理生态响应研究[J]. 农机化研究,2012,34(1):185-189.
[4]张嘉雯,卢绍浩,赵 喆,等. 外源褪黑素对低温胁迫下烟草幼苗生理指标的影响[J]. 中国农业科技导报,2020,22(9):78-86.
[5]杨 妮,万绮雯,李逸民,等. 外源亚精胺对盐胁迫下茶树光合特性及关键酶基因表达的影响[J]. 园艺学报,2022,49(2):378-394.
[6]王汉忠,赵福庚,张国珍.多胺延缓植物衰老的机制[J]. 山东农业大学学报,1995,26(2):227-232.
[7]林必博,王 锋,周济铭. 亚精胺提高植物抗旱性作用机理的研究进展[J]. 贵州农业科学,2020,48(9):6-10.
[8]刘 球,李志辉,吴际友,等. 外源亚精胺对不同干旱胁迫程度下红椿幼苗生理功能的修复调节[J]. 中南林业科技大学学报,2017,37(7):66-72.
[9]李书涵,韩莹琰,郝敬虹,等. 高温胁迫下外源亚精胺对生菜抗氧化酶系统的影响[J]. 北京农学院学报,2020,35(4):63-67.
[10]周晨楠,施晓梦,袁颖辉,等. 外源亚精胺对Ca(NO 3) 2胁迫下番茄幼苗光合特性和抗氧化酶活性的影响[J]. 西北植物学报,2012,32(3):498-504.
[11]赵东晓,董亚茹,孙景诗,等. 外源亚精胺对桑树盐胁迫的缓解效应[J]. 蚕业科学,2019,45(4):484-493.
[12]李合生. 植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[13]刘 领,王艳芳,宋久洋,等. 生物炭与氮肥减量配施对烤烟生长及土壤酶活性的影响[J]. 河南农业科学,2016,45(2):62-66.
[14]田 佳,李 佳,孟清波,等. 不同苹果品种叶片耐热阈值及高温下生理生化响应[J]. 河南农业科学,2021,50(1):121-128.
[15]朱 伟,房卫平,谢德意,等. CaCl 2对盐胁迫下棉花幼苗生长和生理特性的影响[J]. 河南农业科学,2014,43(6):35-38.
[16]薛晓梦,吴 洁,王 欣,等. 低温胁迫对普通和高油酸花生种子萌发的影响[J]. 作物学报,2021,47(9):1768-1778.
[17]张陇艳,程功敏,魏恒玲,等. 陆地棉种子萌发期对低温胁迫的响应及耐冷性鉴定[J]. 中国农业科学,2021,54(1):19-33.
[18]沈 虹,孟佳丽,吴绍军,等. 低温胁迫对西瓜种子萌发的影响[J]. 北方农业学报,2020,48(5):95-103.
[19]倪秀珍,徐基森,蔡佶烨,等. 外源亚精胺浸种对NaCl胁迫下三种牧草种子萌发特性的影响[J]. 黑龙江畜牧兽医,2019(9):118-121.
[20]胡胜男,王 波,李铁华,等. 叶面喷施亚精胺对干旱胁迫下闽楠幼苗生理的影响[J].西南林业大学学报(自然科学),2021,41(6):31-38.
[21]周紫燕,李晓斐,丁雪丹,等. 干旱对烤烟团棵期的光合特性影响研究[J]. 江西农业大学学报,2011,33(6):1037-1042.
[22]王 磊,汤家鑫,高兴国,等. PEG模拟干旱胁迫条件下光叶珙桐幼苗叶片叶绿素含量变化[J]. 安徽农业科学,2018,46(32):91-92,95.
[23]李鹏辉,向金友,王 林,等. 干旱胁迫下外源褪黑素对烟草幼苗生理特性的影响[J]. 中国农业科技导报,2019,21(5):41-48.
[24]武 佶,王禹佳,張欣玥,等. 玉米毛状根再生植株叶片光合特性对干旱胁迫和复水条件的响应[J]. 江苏农业科学,2020,48(15):106-111.
[25]周 建,杨立峰,郝峰鸽,等. 低温胁迫对广玉兰幼苗光合及叶绿素荧光特性的影响[J]. 西北植物学报,2009,29(1):136-142.
[26]李 冬,王艳芳,申洪涛,等. 外源亚精胺对渗透胁迫下烤烟幼苗生长及生理特性的影响[J]. 烟草科技,2019,52(9):1-9.
[27]张永平,张红梅,顾海峰,等. 外源Spd对低温胁迫下甜瓜幼苗光合与抗氧化特性及其相关基因表达的影响[J]. 西北植物学报,2018,38(4):671-681.
[28]后有丽,苏世平,李 毅,等. 外源脱落酸对红砂叶片渗透调节物质含量及抗氧化酶活性的影响[J]. 草业科学,2020,37(2):245-255.
[29]姜秀梅. 外源物质处理对低温胁迫下辣椒种子萌发及幼苗抗冷性的影响[D]. 乌鲁木齐:新疆农业大学,2014.
[30]牛云然. 外源物质对低温下辣椒幼苗抗冷性的影响[D]. 邯郸:河北工程大学,2020.
[31]李素华,余 佳,韩浩章,等. 干旱胁迫下外源ABA处理对孔雀草幼苗生理特性的影响[J]. 内蒙古农业大学学报(自然科学版),2021,42(4):8-11.
[32]朱春权,徐青山,曹小闯,等. 不同属性特征基质对早稻秧苗耐低温的影响[J]. 中国水稻科学,2021,35(5):503-512.
[33]贺嘉豪,陈建中,徐坚强,等. 外源褪黑素对烟草幼苗抗旱性生理机制的影响[J]. 中国农业科技导报,2020,22(2):50-57.
[34]赵永长,宋文静,董建新,等. 黄腐酸钾对干旱胁迫下烤烟幼苗活性氧代谢的影响[J]. 中国烟草科学,2017,38(4):29-36.
猜你喜欢
江苏农业科学(2017年1期)2017-02-27
艺术评鉴(2016年18期)2017-02-22
现代园艺(2016年7期)2017-01-09
山东农业科学(2016年11期)2016-12-17
知音励志·社科版(2016年9期)2016-11-09
中国实用医药(2016年4期)2016-02-23
江苏农业科学(2015年8期)2015-09-10
江苏农业科学(2015年8期)2015-09-10
安徽农学通报(2014年12期)2014-07-28
