
2种常见CRISPR-Cas系统在苹果中的适用性特征
2022-05-09 02:37岳鹏高海琴李哲米志波郑博赵彦敏
江苏农业科学 2022年7期
关键词:苹果
岳鹏 高海琴 李哲 米志波 郑博 赵彦敏

摘要:运用生物信息学方法初步分析CRISPR-Cas9和Cpf1在苹果中的整体适用性特征,以期为苹果基因组编辑和CRISPR-Cas在苹果研究中的推广使用提供一定的参考和便利。结果表明,苹果染色体中有数量可观的PAM,平均间隔7 bp碱基有1个5′-NGG、间隔3 bp有1个5′-TTN;也就是说5′-TTN比5′-NGG的出现频率高。SpCas9、FnCpf1分别有29.0%、26.9%的作用位点几乎覆盖了所有染色体基因,个别不能被SpCas9识别的基因能被FnCpf1识别,反之亦然。苹果的CRISPR靶序列有大量重复,单一靶序列被视为能被Cas蛋白特异识别并有效编辑。在靶序列长度为20 nt时,99.5%的染色体基因可至少被其中1种Cas蛋白编辑,分别具有不同的可编辑度搭配;其中的237个基因只能被1种Cas蛋白编辑,填补了另一种Cas蛋白留下的编辑空白,另有220个染色体基因(0.5%)不能被任一种Cas蛋白编辑,即2种Cas蛋白同时留下编辑空白,没有互补。
关键词:苹果;CRISPR-Cas;适用性特征;PAM出现频率;基因编辑
中图分类号:S661.101 文献标志码: A
文章编号:1002-1302(2022)07-0043-08
收稿日期:2021-06-04
基金项目:中国成人教育协会“十四五”成人继续教育科研规划重点课题(编号:2021-323ZB);河北省张家口市科技项目(编号:18110300043);河北省张家口市社会科学立项研究课题(编号:2021121)。
作者简介:岳 鹏(1984—),男,河北张家口人,硕士,讲师,从事农林生命科学类和开放教育研究。E-mail:yppolymerase@foxmail.com。
通信作者:赵彦敏,博士,副教授,从事农林生命科学类和开放教育研究。E-mail:zym319@163.com。
苹果( Malus domestica )是世界上主要果树作物之一,其产量和质量易受生物和非生物胁迫的影响[1]。因此,了解抗逆性相关基因的功能及调控规律,对培育抗逆性强的品种至关重要[2]。与其他作物相比,果树具有高度杂合的多倍体基因组且繁育周期长,导致传统的育种研究进展缓慢[3];而随着基因组测序工作的完成,对其基因结构、基因通路和基因功能的认识为基因编辑奠定了基础[4]。高效、易用、省时、低成本的基因编辑技术是赋予果树重要经济性状的捷径。常间四文重复序列丛集关联蛋白[CRISPR(clustered regularly interspaced short palindromic repeat)-Cas(CRISPR-associated proteins)]系统具备以上优势,在研究生物多种类的精确分子机制方面具有极高的应用价值[5]。
当前,基因组编辑技术主要集中在第2类CRISPR-Cas系统[6];它可以使用单个效应蛋白剪切DNA,包括Ⅱ型、Ⅴ型、Ⅵ型[7]。其中,Ⅱ型的Cas9及其失去剪切活性的衍生品dCas9被广泛应用于多种生物体的基因组操作,包括靶向基因干扰、转录激活抑制、表观遗传修饰及目标碱基对转换[8-9]。经典的Cas9蛋白须识别衔接在靶序列(+)3′端形如5′-NGG 的前间隔序列邻近基序(protospacer-adjacent motif,PAM,+),并在PAM上游第3个碱基处切割双链,形成平头末端;在人类基因组中约平均间隔8 bp就会有1个5′-NGG[10]。不同于此,Ⅴ型的Cpf1(Cas12a)要识别毗邻在靶序列(+)5′端形似5′-TTN 的PAM(+),并在PAM下游的同向链(+)第18、互补链(-)第23碱基处交错切割,形成有5个突出碱基的黏性末端[11]。作为新型且更小的CRISPR效应蛋白,Cpf1的开发利用有利于突破和克服Cas9在使用中的一些限制,尤其是拓宽了CRISPR-Cas系统的识别范围,使之能更有效地编辑富含AT碱基的基因组[12]。
CRISPR-Cas9已被大量使用在植物基因功能分析及育種研究中[13-15],其中包括多种果树[14]。有关苹果的几项研究,依次是首次利用CRISPR-Cas9敲除内源 PDS (phytoene desaturase)基因[16],高效传送CRISPR-Cas9核糖核蛋白到苹果原生质体操纵DIPM1( DspA/E -interacting proteins of M.×domestica )、DIPM2和DIPM4基因[17],优化CRISPR-Cas9的应用条件并敲除PDS和TFL1(terminal flower)基因[3],运用CRISPR-Cas9敲除DIPM4基因的同时又减少外源DNA的残留[18]。相对地,CRISPR-Cpf1的技术特点更加鲜明,但在2016年首次应用于水稻和烟草后[19],在植物基因组项目中使用的报道较少,目前还未见于果树研究[2]。
已有的研究结果表明,CRISPR-Cas系统可实际应用于苹果基因编辑,但只关注了少数几个基因,且仅限于使用CRISPR-Cas9。本研究对苹果全基因组序列进行初步分析,尝试探索2种流行的CRISPR-Cas系统,即CRISPR-Cas9和CRISPR-Cpf1,在苹果基因组编辑中的整体适用性,首次从PAM的数量和频率、靶序列的重复和分布及2种Cas蛋白的互补等几个方面开展讨论,并形成PAM位点和靶序列信息库,以期为苹果基因组编辑和CRISPR-Cas系统在苹果研究中的推广使用提供一定的参考和便利。
1 材料与方法
1.1 数据获取
苹果全基因组代表性数据ASM211411v1下载自NCBI网站[20],17条染色体(chromosome,chr)和1条线粒体(mitochondrion,mt)的DNA序列以FASTA格式分别存储在文本文件中;所有下载的DNA都只有单链(+)。相应的基因文件也从NCBI网站下载,其中以列表形式记录基因在DNA序列上的起止位点等信息。最新发布在NCBI网站上的苹果基因组数据ASM411538v1质量更高[21],但相应的基因信息不够完善,只在本研究中作对比和补充。在诸多CRISPR-Cas系统中,效应蛋白SpCas9[化脓性链球菌( Streptococcus pyogenes ,Sp)]和FnCpf1[弗郎西斯菌( Francisella novicida ,Fn)]识别的PAM较有代表性,分别为5′-NGG和5′-TTN[10-11];本研究围绕这2种典型的PAM,尝试使用个人计算机分析苹果全基因组。
1.2 PAM计数和间距计算
对苹果各DNA序列出现的5′-NGG或 5′-TTN 计数,并记录中间碱基在DNA序列上的位点作为PAM位点;还需要同时累计5′-CCN或 5′-NAA,以实现对互补链的搜索。计数期间,单独计算N所代表的各种碱基占比,并用PAM位点数量与DNA长度的比值表示序列的PAM密度。除线粒体外,合计染色体DNA的各项数据以考量全基因组。
同样地,分别计算各DNA序列和全基因组的PAM出现频率。将每2个相邻PAM位点之差作为间距(用字母 d 表示),其意义为间隔 d 个碱基对存在1个PAM,代表PAM的出现频率;并累计每个 d的出现次数(用字母n表示),再把全部d升序排列(以t为排列序号),记录不同d的数量(用字母m 表示)。PAM出现频率的均值( d mean)和中值( d median)计算如下:
d mean=∑ mi=1d in i∑mi=1n i;若∑ti=1n i≥∑mi=1n i2>∑t-1i=1n i,d median =d t 。
1.3 计算剪切位点
在DNA序列上从5′-NGG前溯3个碱基或从5′-CCN后推3个碱基,获得Cas9剪切位点。通过判断剪切位点是否处于基因的起止位点之间,将对应的PAM位点划入相应的基因范畴;不属于任何基因范畴的PAM位点不做标记。
依据Cpf1的剪切特征,在DNA序列上从5′-TTN后推18个碱基或从5′-NAA前溯23个碱基,获得同向剪切位点,用来判断相应的PAM位点是否属于同向链基因;从5′-TTN后推23个碱基或从 5′-NAA 前溯18个碱基,获得逆向剪切位点,用来判断相应的PAM位点是否属于互补链基因。
1.4 靶序列的截取
根据2种PAM的特点,在DNA序列上分别截取长度为20 nt(不含PAM)的靶序列(target),用PAM位点命名,以FASTA格式储存;PAM在互补链上的,还需要按照碱基配对原则转换碱基并逆序。对具有同种PAM的靶序列进行重复性搜索,找到单一序列(singleton)和相同序列簇(cluster);簇中的PAM位点有属于染色体基因的计1分,有属于染色体基因间隔的计2分,有属于线粒体基因的计4分,有属于线粒体基因间隔的计8分,4种分值任意组合可将所有簇归入15个重复类(用repeat N 命名, N 为不大于15的正整数)。重点关注靶序列单一且属于基因范畴的PAM位点,其数量与基因长度的比值表示基因的可编辑度;按照“基因ID-PAM位点-靶序列”的模式建立简易信息库,找到可编辑度最高和最低的基因。
1.5 程序实现
以上操作已被整合到几个Perl脚本中,并尽量优化算法降低时间复杂度和空间复杂度[22]。其中,为避免耗费大量时间,在判断PAM位点是否影响基因时,用数组模拟DNA序列,数组索引表示位点,基因区间内的数组元素由基因ID填充,其他为未定义(undef)值;当剪切位点对应的数组元素为基因ID时,将PAM位点划入此基因范畴。为避免占用大量内存空间导致程序卡顿,在搜索重复靶序列时,把总文件分割成大小合适的几个子文件,使用散列快速剔除子文件内的相同序列;然后利用每个子文件中序列唯一的特点,使用散列剔除子文件间的相同序列,最终合并成没有重复序列的总文件。
2 结果与分析
2.1 PAM含量分析
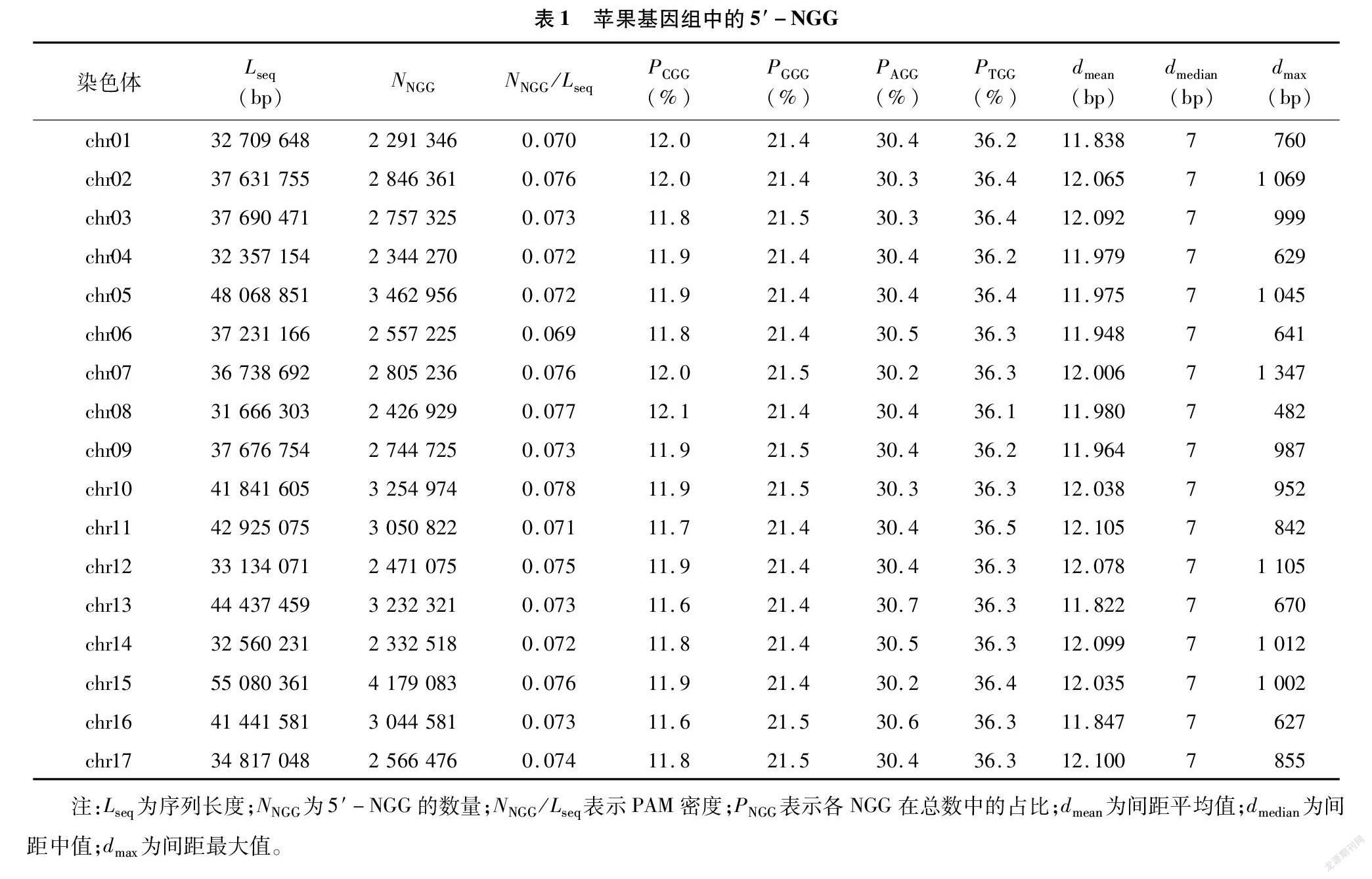
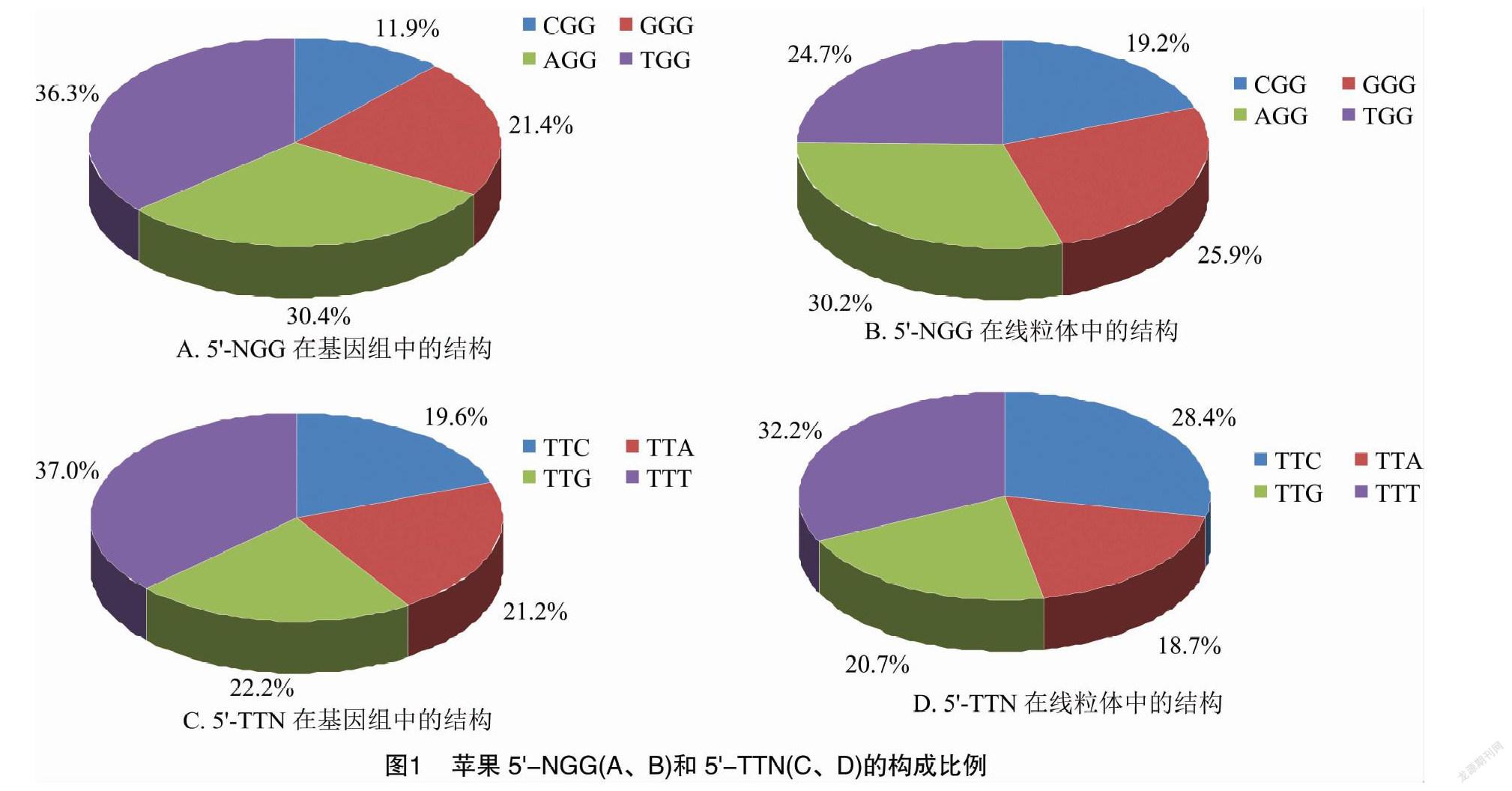
苹果基因组(ASM211411v1)中5′-NGG总量为48 368 223,chr15的数量最多(4 179 083),chr01的最少(2 291 346),各染色体间差异较大;再结合序列长度估算序列的PAM密度,基因组是0.074,chr10最高(0.078),chr06最低(0.069),相差不大(表1)。在基因组中,CGG、GGG、AGG和TGG在NGG总量中的比例分别为11.9%、21.4%、30.4%、36.3%(图1-A);在各染色体中的比例与此一致,略有微小波动(表1)。线粒体中的5′-NGG含量为 46 468,但密度达到0.117,高于基因组最高值;CGG、GGG、AGG和TGG的比例分别为19.2%、259%、30.2%、24.7%,与基因组中的同项数据也有明显差异(图1-B)。
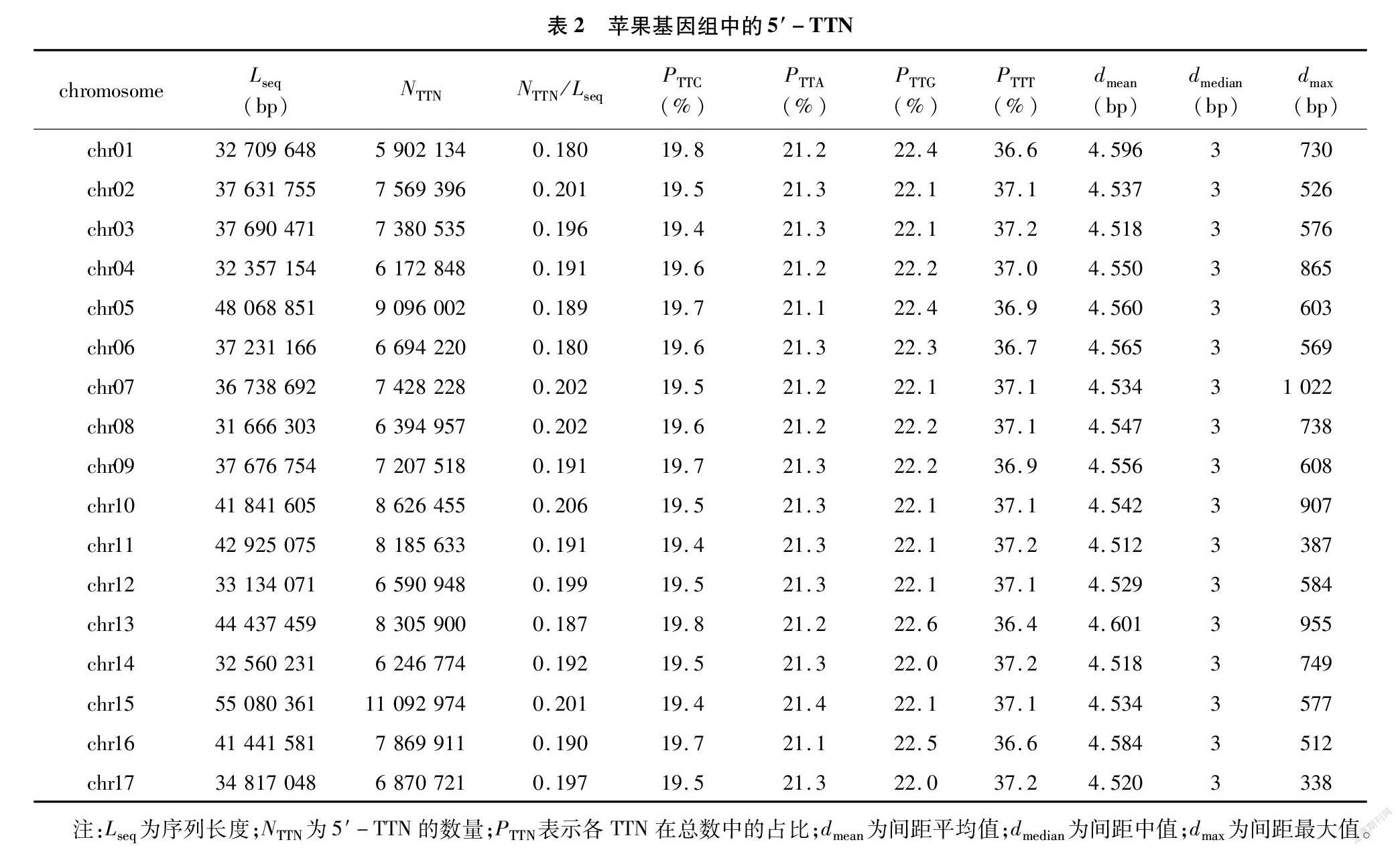
基因组中5′-TTN的总数为127 635 154,同样是chr15最多(11 092 974),chr01最少(5 902 134);估算基因组的密度是0.194,依然是chr10最高(0206),chr06最低(0.180),两者比较接近(表2)。在基因组中,TTC、TTA、TTG和TTT在TTN总量中的比例分别为19.6%、21.2%、22.2%、37.0%(图1-C);同样,在各染色体中的比例与此一致(表2)。线粒体中5′-TTN含量为69 296,密度只有0.174 6,低于基因组最低值;TTC、TTA、TTG和TTT的比例分别为28.4%、18.7%、20.7%、32.2%,与基因组中的同項数据也差异明显(图1-D)。
作为对比,另一基因组数据(ASM411538v1)的5′-NGG密度为0.084,CGG、GGG、AGG和TGG的比例分别为11.7%、21.5%、30.6%、36.3%;5′-TTN的密度为0.219,TTC、TTA、TTG和TTT的比例分别是19.6%、21.2%、22.4%、36.8%。使用2个不同版本的基因组数据计算出的结果很接近,特别是NGG和 TTN的构成比例几乎完全一致,可以用同一组饼状图表示(图1-A、图1-C)。
2.2 PAM出现频率
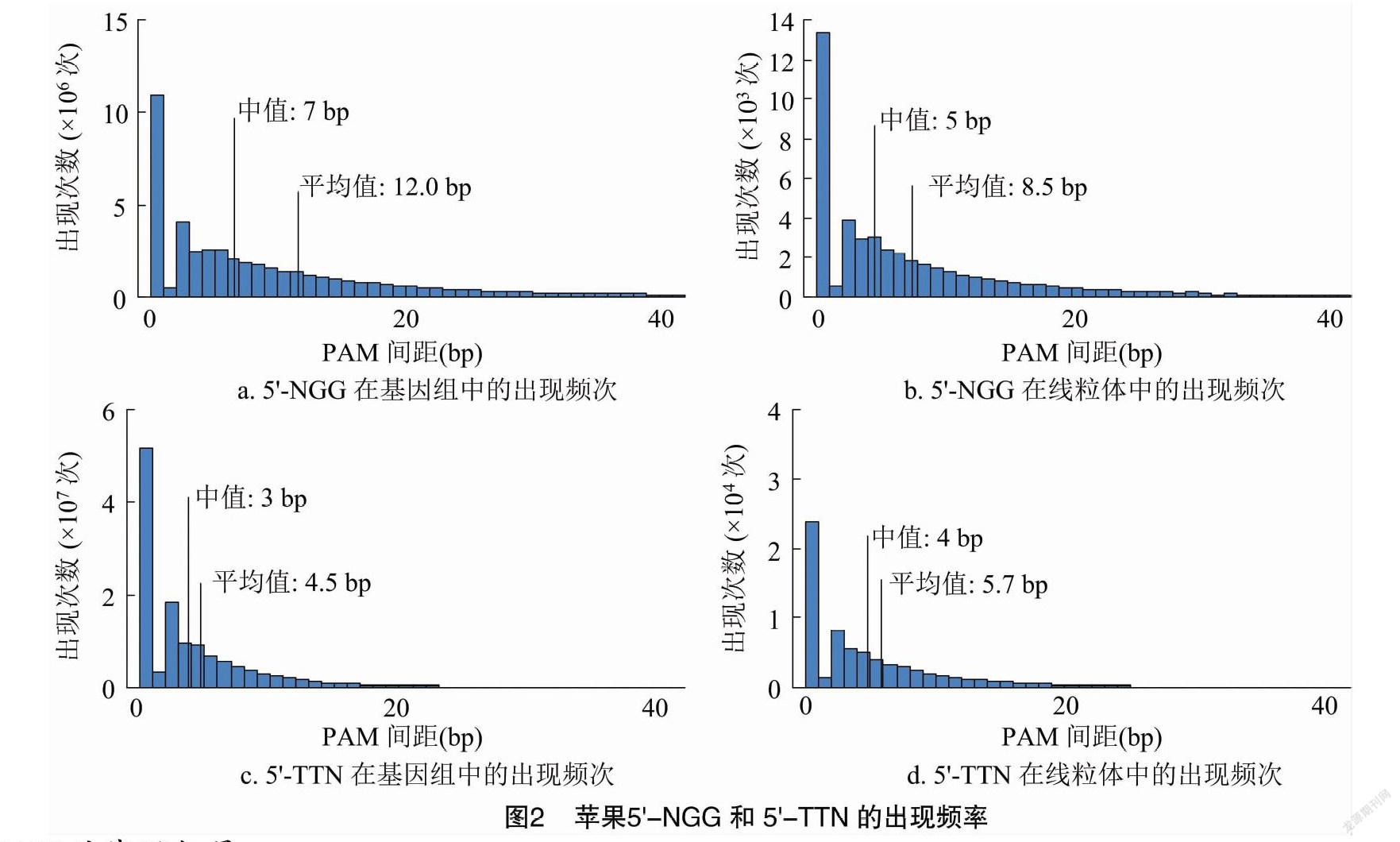
在苹果基因组(ASM211411v1)中,相邻5′-NGG的间距最小为1 bp,出现次数最多(图2-A);最大为1 347 bp,出现在chr07的19 065 740~19 067 087位点间(表1)。间距中值是7 bp,表示平均约间隔7 bp碱基就有1个5′-NGG,也说明半数以上的间距不大于7 bp;间距均值为12.0 bp,是受到最大值的影响而产生了偏移(图2-a)。各染色体情况与此高度一致(表1)。而在线粒体中,间距中值和均值分别为5、8.5 bp,与基因组完全不同(图2-b)。
基因组中,相邻5′-TTN的间距最小为1 bp,出现次数最多(图2-c);最大为1 022 bp,出现在chr07的26 831 153~26 832 175位点间(表2)。间距中值是3 bp,表示平均约间隔3 bp碱基就有1个 5′-TTN,也说明半数以上的间距不大于3 bp;受最大值影响,间距均值是4.5 bp(图2-c)。同样,各染色体情况与此一致(表2)。线粒体的间距中值和均值分别为4、5.7 bp,也与基因组完全不同(图2-d)。使用另一版本的基因组数据(ASM411538v1)计算,可得出类似的结果:5′-NGG的间距中值和均值分别为7、11.9 bp,5′-TTN的间距中值和均值分别为3、4.5 bp;所展示出的变化趋势也可以绘成同样的图像(图2-a、图2-c)。
2.3 PAM的基因归属
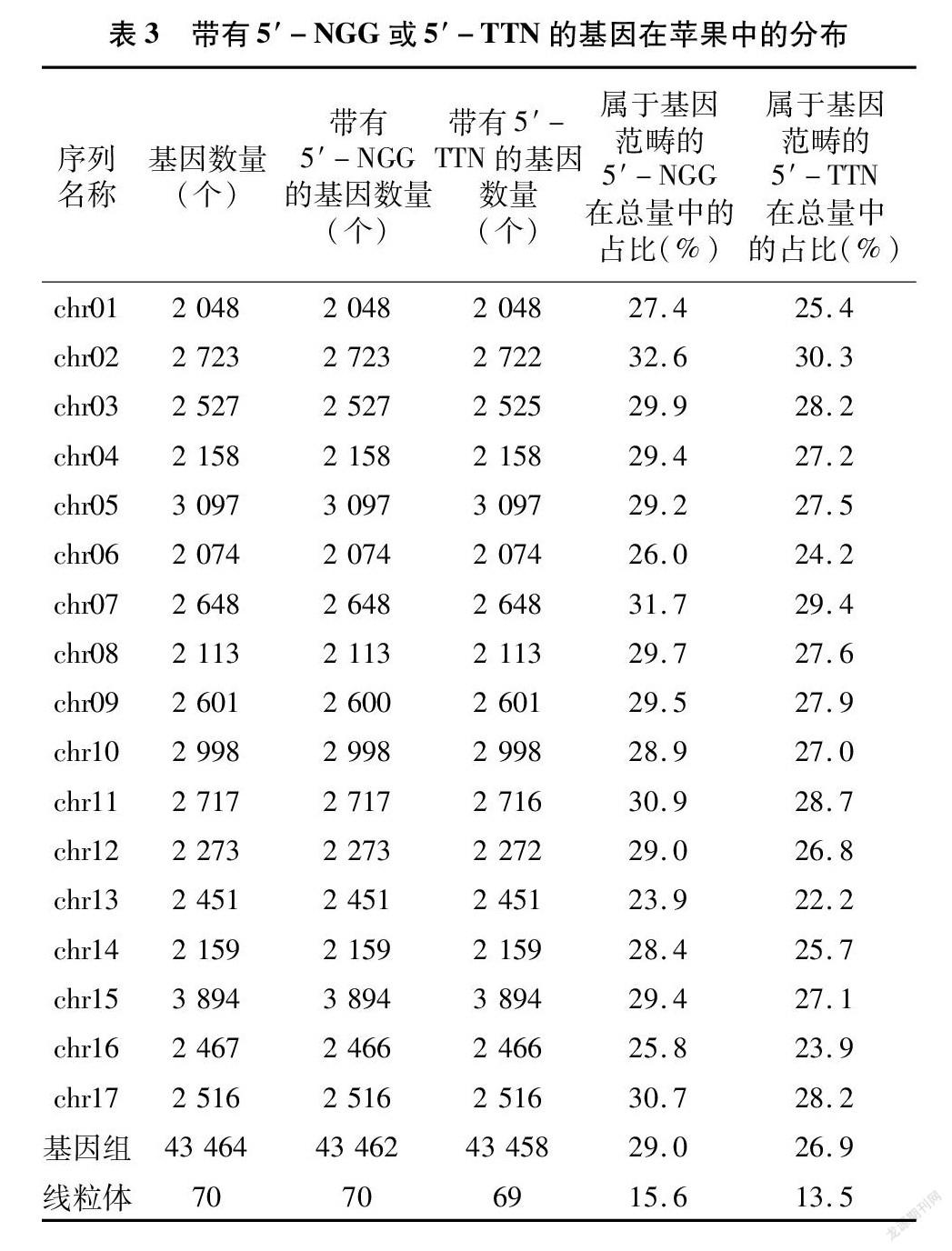
在苹果基因组中,29.0%的5′-NGG和26.9%的5′-TTN能影响到基因,基本覆盖了全部43 464个基因(表3)。其中,chr06上ID為114825448的基因含5′-NGG最多,有14 620个;chr16的114822391(表示基因ID,下同)和chr09的114827208基因不含5′-NGG。chr06的114825448基因含5′-TTN最多,有36 069个;chr16的108169786、chr12的108174957、chr11的108174696、chr02的114823832、chr03的114824289和108171505基因都不含5′-TTN。
在线粒体中,15.6%的5′-NGG能作用于全部70个基因,13.5%的5′-TTN覆盖了98.6%的基因(表3)。其中,ID为13630194的基因含5′-NGG最多,有1 186个;13630239(121 892~121 913位点)和13630229基因含5′-NGG最少,只有5个。13630194基因含5′-TTN最多,有1 337个;13630239基因(121 892~121 913位点)不含5′-TTN。与各染色体相比,线粒体中属于基因间隔的PAM占比明显更大(表3)。
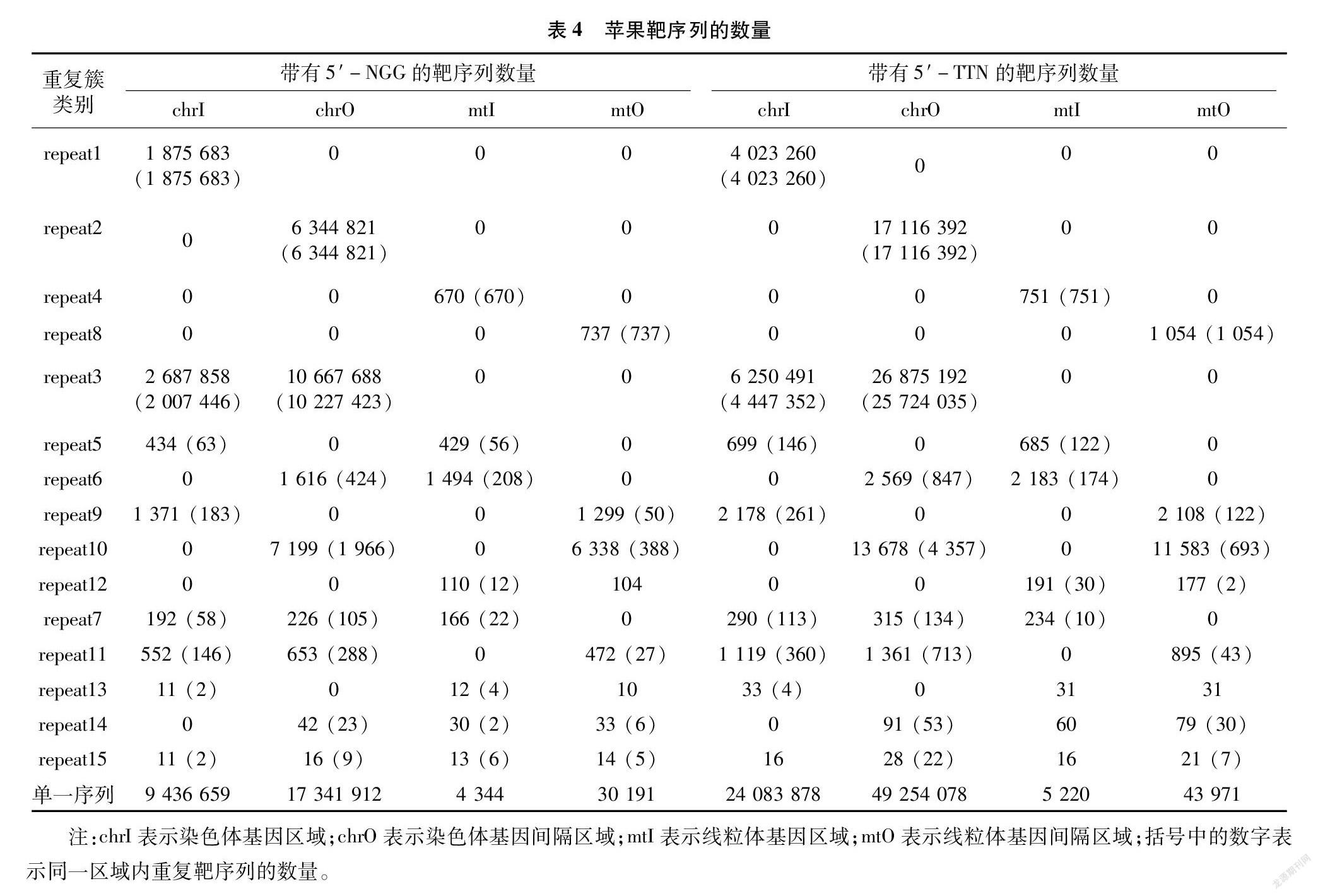
2.4 靶序列的重复簇
苹果的CRISPR靶序列有大量重复,共计15种重复簇(表4)。其中,重复序列只属于染色体基因的簇划入repeat1,只属于染色体基因间隔的为repeat2,只属于线粒体基因的为repeat4,只属于线粒体基因间隔的为repeat8,repeat15是重复的靶序列同时出现在上述4个区域中。可以发现,带有 5′-NGG 的靶序列,染色体基因的648条与线粒体基因中的620条重复;带有5′-TTN的靶序列,染色体基因中的1 038条与线粒体基因中的966条重复(表4,repeat5、7、13、15)。只属于染色体基因的,带有5′-NGG的重复靶序列共有3 883 583条,带有5′-TTN的重复靶序列共有84 71 496条(表4,repeat1、3、5、9、7、11、13、15括号中的数字);而在线粒体基因中,这2个数值分别为980、1 087(表4,repeat4、5、6、12、7、13、14、15括号中的数字)。
2.5 基因可编辑度
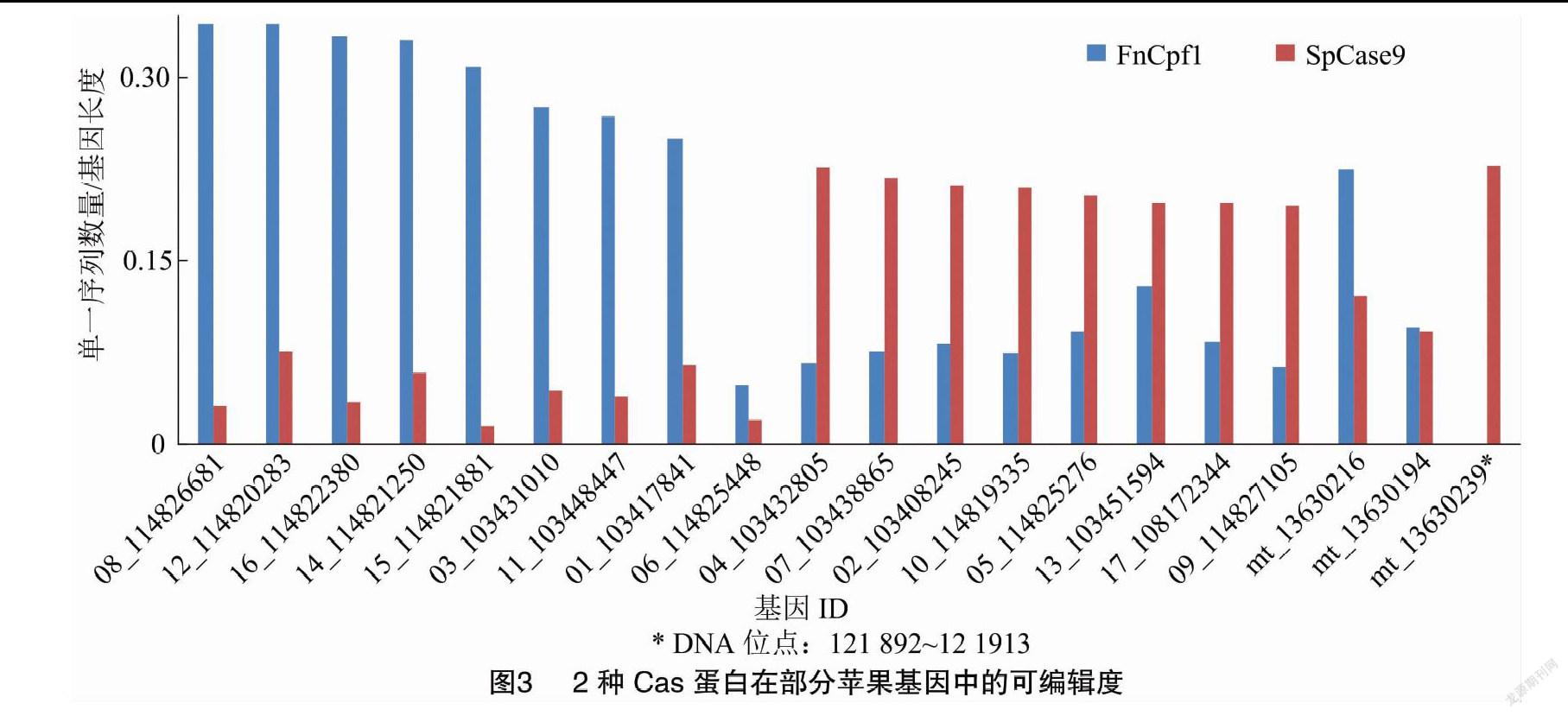
在苹果各染色体中,带有5′-NGG的单一靶序列数量为26 778 571,其中属于基因的有9 436 659,其余都处于基因间隔中;带有5′-TTN的单一靶序列数量为73 337 956,属于基因的有240 838 78(表4)。SpCas9对chr04上的103432805基因有最高的可编辑度,为0.226,含有PAM最多的114825448基因的可编辑度仅为0.020(图3);另有372个基因的可编辑度为0。FnCpf1对chr08上的114826681基因有最高的可编辑度,为0.344,含有PAM最多的114825448基因的可编辑度仅为0.049(图3);另有305个基因的可编辑度为0。
在线粒体中,带有5′-NGG的单一靶序列数量为34 535,属于基因的有4 344;带有5′-TTN的单一靶序列数量为49 191,属于基因的有5 220(表4)。SpCas9对13630239基因(121892~121913位点)可能有最高的可编辑度,为0.227,含有PAM最多的13630194基因的可编辑度为0.092(图3);有4个基因的可编辑度为0。FnCpf1对13630216基因可能有最高的可编辑度,为0.224,含有PAM最多的13630194基因的可编辑度为0.095(图3);有6个基因的可编辑度为0。
2.6 Cas蛋白的编辑互补
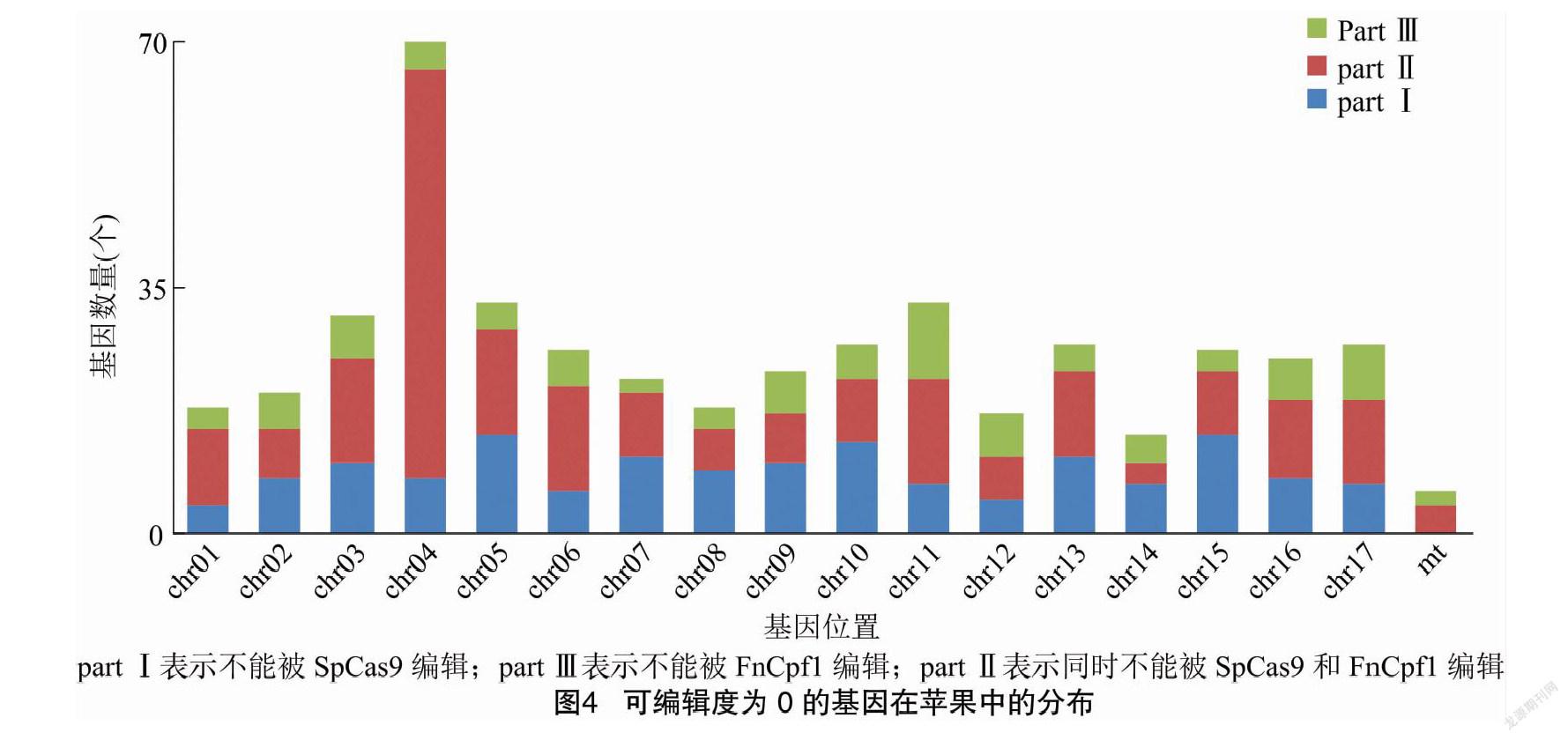
苹果中的大多数基因可同时被2种Cas蛋白编辑, 分别具有不同的可编辑度搭配(图3)。 可编辑度为0的基因在全部基因中的占比较小,在各DNA序列上均有分布;其中,chr04上的数量最多,有66个不能被SpCas9编辑、有62个基因不能被FnCpf1编辑(图4)。经过筛选,共有237个(0.5%)染色体基因、2个(2.9%)线粒体基因能被1种Cas蛋白编辑,填补了另一种Cas蛋白留下的编辑空白(图4,part Ⅰ、part Ⅲ);共有220个染色体基因(0.5%)、4个(5.7%)线粒体基因不能被任一种Cas蛋白编辑,即2种Cas蛋白同时留下编辑空白,没有互补(图4,part Ⅱ)。
3 讨论
作为重要的基因编辑工具[23],CRISPR-Cas系统在苹果基因组中有较好的整体适用性,主要表现在3个方面。一是有数量可观的PAM分散在苹果DNA序列的各个角落,出现频率很高,平均间隔很短。二是Cas蛋白的作用位点几乎覆盖了所有基因,个别不能被SpCas9识别的基因却含有FnCpf1的识别位点,反之亦然。三是拥有单一靶序列的基因占大多数,99.5%的染色体基因和94.3%的线粒体基因都能至少被其中1种Cas蛋白编辑。苹果DNA序列的测序结果表明,AT碱基的含量高于CG碱基[20-21],导致5′-TTN的数量远超5′-NGG、5′-TTN 的出现频率更高、带有5′-TTN的单一靶序列数量更多,也就是说Cpf1在苹果基因编辑中有更大的可挖掘潜力。
各染色體的PAM密度、组成和出现频率几乎一致,可视作苹果基因组的整体特征之一。虽然也存在于苹果细胞中,但线粒体通常被认为是有益的共生生物[24],其DNA不被计入基因组;这一点在本研究中也有突出体现,即与染色体的同项数据相比,线粒体都有明显差异。目前,对苹果线粒体基因的编辑还未见报道,其操作过程是否与基因组相同还需要更深入的研究验证,在本研究中仅是预测性的初步探讨;且线粒体DNA体量小、基因少[20],对基因组编辑的影响不大,在特定环境中可不做考虑。叶绿体也有类似情况[25],可在条件成熟时进一步研究讨论。
已有的测序结果含有未知碱基,在数亿碱基的苹果DNA序列中比例微小,对多项计算结果的影响可忽略不计[20-21]。但如果2个PAM之间存在未知碱基且结合上下游无法判断是否存在另一个PAM,就在确定PAM频率时摒弃这2个PAM的间距,避免出现超长间距的同时,也保证了是在计算相邻2个PAM的间距。对比人类基因组取间距中值作为PAM的出现频率,本研究也采用了同样的取值方法;苹果基因组平均间隔7 bp碱基就有1个5′-NGG,频率高于人类基因组的8 bp[10]。
一般,判断PAM是否属于基因依据的是其位点是否在基因起止位点间,临近基因边缘的PAM就有可能实际作用到了间隔区。不同于此,本研究将判断依据改进为剪切位点是否在基因起止位点间,既避免了上述问题,也充分挖掘了隐藏的基因PAM。在此基础上截取的靶序列都具有明确的基因归属。靶序列的长度按常规被设定为20 nt,初步分析了因序列重复导致的脱靶情况;根据序列越短重复率越高的共识,可适当增加靶序列的长度提高基因的可编辑度。此外,脱靶的原因还包括相似匹配和种子序列的长度[11],可在未来的研究中做更深入的分析。
基因可编辑度与单一靶序列的数量成正比,与基因自身的长度成反比,表示的是单位长度内含有的备选靶序列密度。可编辑度为0的基因,小部分是因为不含有PAM,大多数是在屏蔽了重复靶序列后,备选数量为0。本研究采用了较严格的屏蔽标准,凡是在苹果DNA序列中出现的重复靶序列均计入重复簇;重复簇的类别划分较细,15个类别涵盖了靶序列所在4个区域的所有搭配,方便在试验设计时有侧重地取舍。在苹果全部基因中,2种Cas蛋白都有占比很小的编辑盲区,FnCpf1要好于SpCas9。盲区重叠的部分所含的224个基因不适宜使用这2种Cas蛋白编辑,可考虑换用识别不同PAM的其他Cas蛋白;其中超过半数的基因(139个)编码多种RNA,通常在实际研究中较少涉及到。
在Perl脚本的帮助下,各步骤的运算结果都在文本文件中详细列表构成了信息库,可直接打开查询感兴趣的信息;也可导入数据库加以专业化的管理和维护,成为网络服务平台的构建基础,这是开展下一步研究的一个重要方向。
参考文献:
[1]Arzani A,Ashraf M. Smart engineering of genetic resources for enhanced salinity tolerance in crop plants[J]. Critical Reviews in Plant Sciences,2016,35(3):146-189.
[2]Wang X H,Tu M X,Li Z,et al. Current progress and future prospects for the clustered regularly interspaced short palindromic repeats (CRISPR) genome editing technology in fruit tree breeding[J]. Critical Reviews in Plant Sciences,2018,37(4):233-258.
[3]Charrier A,Vergne E,Dousset N,et al. Efficient targeted mutagenesis in apple and first time edition of pear using the CRISPR-Cas9 system[J]. Frontiers in Plant Science,2019,10:40.
[4]Zhou J H,Li D D,Wang G M,et al. Application and future perspective of CRISPR/Cas9 genome editing in fruit crops[J]. Journal of Integrative Plant Biology,2020,62(3):269-286.
[5]Yan F C,Wang W,Zhang J Q. CRISPR-Cas12 and Cas13:the lesser known siblings of CRISPR-Cas9[J]. Cell Biology and Toxicology,2019,35(6):489-492.
[6]Strecker J,Jones S,Koopal B,et al. Engineering of CRISPR-Cas12b for human genome editing[J]. Nature Communications,2019,10:212.
[7]Makarova K S,Zhang F,Koonin E V. Snapshot:class 2 CRISPR-Cas systems[J]. Cell,2017,168(1/2):328-328e1.
[8]Hu J H,Miller S M,Geurts M H,et al. Evolved Cas9 variants with broad PAM compatibility and high DNA specificity[J]. Nature,2018,556(7699):57-63.
[9]Moradpour M,Abdulah S N A. CRISPR/dCas9 platforms in plants:strategies and applications beyond genome editing[J]. Plant Biotechnology Journal,2020,18(1):32-44.
[10]Hsu P D,Lander E S,Zhang F. Development and applications of CRISPR-Cas9 for genome engineering[J]. Cell,2014,157(6):1262-1278.
[11]Zetsche B,Gootenberg J S,Abudayyeh O O,et al. Cpf1 is a single RNA-guided endonuclease of a class 2 CRISPR-Cas system[J]. Cell,2015,163(3):759-771.
[12]楊 帆,李 寅. 新一代基因组编辑系统CRISPR/Cpf1[J]. 生物工程学报,2017,33(3):361-371.
[13]Ma X L,Zhu Q L,Chen Y L,et al. CRISPR/Cas9 platforms for genome editing in plants:developments and applications[J]. Molecular Plant,2016,9(7):961-974.
[14]Wang X H,Tu M X,Wang D J,et al. CRISPR/Cas9-mediated efficient targeted mutagenesis in grape in the first generation[J]. Plant Biotechnology Journal,2018,16(4):844-855.
[15]Rodríguez-Leal D,Lemmon Z H,Man J,et al. Engineering quantitative trait variation for crop improvement by genome editing[J]. Cell,2017,171(2):470-480,e8.
[16]Nishitani C,Hirai N,Komori S,et al. Efficient genome editing in apple using a CRISPR/Cas9 system[J]. Scientific Reports,2016,6:31481.
[17]Malnoy M,Viola R,Jung M H,et al. DNA-free genetically edited grapevine and apple protoplast using CRISPR/Cas9 ribonucleoproteins[J]. Frontiers in Plant Science,2016,7:1904.
[18]Pompili V,Costa L D,Piazza S,et al. Reduced fire blight susceptibility in apple cultivars using a high-efficiencyCRISPR/ Cas9-FLP/FRT-based gene editing system[J]. Plant Biotechnology Journal,2020,18(3):845-858.
[19]Endo A,Masafumi M,Kaya H,et al. Efficient targeted mutagenesis of rice and tobacco genomes using Cpf1 from Francisella novicida [J]. Scientific Reports,2016,6:38169.
[20]Daccord N,Celton J M,Linsmith G,et al. High-quality de novo assembly of the apple genome and methylome dynamics of early fruit development[J]. Nature Genetics,2017,49(7):1099-1106.
[21]Zhang L Y,Hu J A,Han X L,et al. A high-quality apple genome assembly reveals the association of a retrotransposon and red fruit colour[J]. Nature Communications,2019,10:1494.
[22]Alsuwaiyel M H.Algorithms:design techniques and analysis (revised edition)[M]. Singapore:World Scientific Publishing,2016:20-34.
[23]Koonin E V,Makarova K S,Zhang F. Diversity,classification and evolution of CRISPR-Cas systems[J]. Current Opinion in Microbiology,2017,37:67-78.
[24]Nykky J,Vuento M,Gilbert L.Role of mitochondria in parvovirus pathology[J]. PLoS One,2014,9(1):e86124.
[25]Waters M T,Langdale J A.The making of a chloroplast[J]. The EMBO Journal,2009,28(19):2861-2873.
猜你喜欢
小天使·一年级语数英综合(2017年5期)2017-06-10
小天使·六年级语数英综合(2017年4期)2017-04-18
小学生导刊(低年级)(2016年8期)2016-09-24
学苑创造·A版(2016年8期)2016-07-06
阅读与作文(小学低年级版)(2016年3期)2016-03-08
学苑创造·A版(2015年10期)2015-11-13
小天使·一年级语数英综合(2015年2期)2015-01-14
小雪花·成长指南(2014年10期)2014-10-31
