
若尔盖地区主要覆被类型浅层土壤持水量和有机碳关系研究
2022-04-25 13:14付琳玉孙永玉胥汝宇刘文斗廖声熙唐国勇
草地学报 2022年4期
付琳玉, 孙永玉, 胥汝宇, 刘文斗, 廖声熙, 唐国勇,4*
(1. 中国林业科学研究院高原林业研究所, 云南 昆明 650233; 2. 南京林业大学, 江苏 南京 210037; 3. 国家林业局云南元谋荒漠生态系统定位观测站, 云南 昆明 650233; 4. 国家林业局云南建水荒漠生态系统定位观测站, 北京 100083)
水源涵养是林草生态系统重要的生态功能之一[1]。若尔盖地区地处青藏高原东缘,是我国高寒草甸和高原泥炭典型分布区,也是黄河、长江重要的水源补给区,在土壤碳水固持方面发挥着重要作用[2-3]。资料显示,若尔盖地区水源涵养总量约为 6. 57亿m3/年,向黄河补水 6.71±1.49亿m3/年,径流量与水源涵养量正相关关系[4-5]。近百年来人类活动和气候变化等干扰,导致若尔盖高寒草甸和湿地生态系统退化[6-7]。合理放牧、人工建植等恢复措施虽使水源涵养量呈上升趋势,有机碳储量显著增加[4,8-9],但为了更精准判断全球变化背景下区域碳水功能评估,明确该区域土壤持水量和有机碳关系这一议题应该得到重视。
土壤持水能力是陆地生态系统水源涵养的重要指标,在地表过程和水循环中发挥关键作用[10],主要受土壤理化性质的影响,如土壤容重、孔隙度、土壤粒度分布、土壤结构和有机质等[11],其中,土壤有机质与土壤持水能力具有极强的相关性[12-14]。土壤有机质积累可以通过改变土壤结构,或通过改变土壤孔性而改善水结合和保水能力,从而提高土壤持水能力[13-14],尤其是在土壤浅层[15]。Farley等[12]对厄瓜多尔帕拉莫原上的蒙特利松林分土壤研究表明,高土壤含水量可以延缓有机碳分解,提高碳储量,增加土壤中孔和微孔量,从而提高土壤持水能力;相反土壤水损失会引起碳损失,从而又降低土壤持水能力。高寒草甸地区土壤有机质是土壤孔性和结构的关键建构因子,对土壤持水性的影响大于土壤质地的贡献,是该地区水文模型的重要参数[13,16],在估算地表土壤持水量中发挥重要作用[15]。前人研究了高寒草甸土壤持水能力和有机碳含量等诸多问题,然而,对这两者具体的相关关系尚缺乏研究。本文选取四川省若尔盖县、红原县和甘肃省玛曲县部分地区的草甸和天然林地作为研究对象,研究土壤持水量和有机碳关系,为更精准判断全球变化背景下若尔盖地区碳水功能评估提供科学依据。
1 材料与方法
1.1 研究区概况
本研究选取四川省若尔盖县、红原县和甘肃省玛曲县部分地区作为研究区域,其平均海拔3 500 m,属高原亚寒带半湿润大陆性季风气候区,年平均气温 0.7℃,年均降水量 600~800 mm,其中85%降水量集中在5—9月,年均相对湿度69%,年均日照时数2 389 h。研究区主要土地类型是高寒草甸和天然针叶林,主要覆被为沼泽化草甸、非沼泽化草甸和天然林地,主要土壤类型为草甸土,有少量泥炭土、沼泽土和风沙土,主要植物物种、洞穴和放牧情况如表1所示,由于自然差异极其显著,可以推断非沼泽化草甸鼠兔害和放牧强度明显大于沼泽化草甸。

表1 若尔盖地区主要覆被类型基本信息表
1.2 样品采集
2020年8月(雨季中期),根据研究区遥感影像和实地考察,并充分考虑海拔、坡度、坡位、坡向、覆被类型等生态因子,在研究区内共设94个样点,包括沼泽化草甸、非沼泽化草甸和天然林地3种主要覆被类型。每个样点设5 m×5 m样方,在样方内按五点法取2个浅层(0~20 cm)土壤混合样(该地区沼泽化草甸雨季中期暂未淹水),用于测定土壤有机碳含量;每个样点在0~20 cm中部位置随机取6个铝盒样品和6个100 cm3环刀样品,用于测量土壤含水量、容重、孔隙度、持水量等指标(参照《中华人民共和国林业行业标准——森林土壤分析方法》),取平均值,并记录样点坐标信息、覆被类型、植被状况、海拔、坡度、坡向、坡位等基本信息。
1.3 室内分析
将土壤混合样风干,研磨过筛(80目)。采用烘干法测量土壤自然含水量;环刀法测量土壤容重、总孔隙度、毛管孔隙度、非毛管孔隙度以及相关持水特征(田间持水量、最大持水量、毛管持水量、非毛管持水量);重铬酸钾外加热法测土壤有机碳含量。
1.4 数据分析
用方差分析(ANOVA,LSD检验)比较不同覆被类型间土壤孔隙度、持水量、有机碳含量等理化参数差异的显著性,用相关分析(Pearson检验)探究土壤孔性、持水量、有机碳含量之间的相关性,用线性回归分析探究土壤水碳相关特征。以上统计分析在SPSS 23.0上完成,图表用Origin 2019和Excel 2007整理绘制。
2 结果与分析
2.1 主要覆被类型土壤持水能力特征
不同覆被类型浅层土壤总孔隙度、毛管孔隙度和非毛管孔隙度均具有显著差异(P<0.05,图1)。沼泽化草甸总孔隙度最大(88.4%),分别是天然林地(82.2%)和非沼泽化草甸(64.6%)的1.1和1.4倍。沼泽化草甸土壤毛管孔隙度(56.1%)约是天然林地(49.3%)和非沼泽化草甸(47.1%)的1.2倍。天然林地非毛管孔隙度最大(32.8%),其次为沼泽化草甸(32.3%)和非沼泽化草甸(17.4%)。
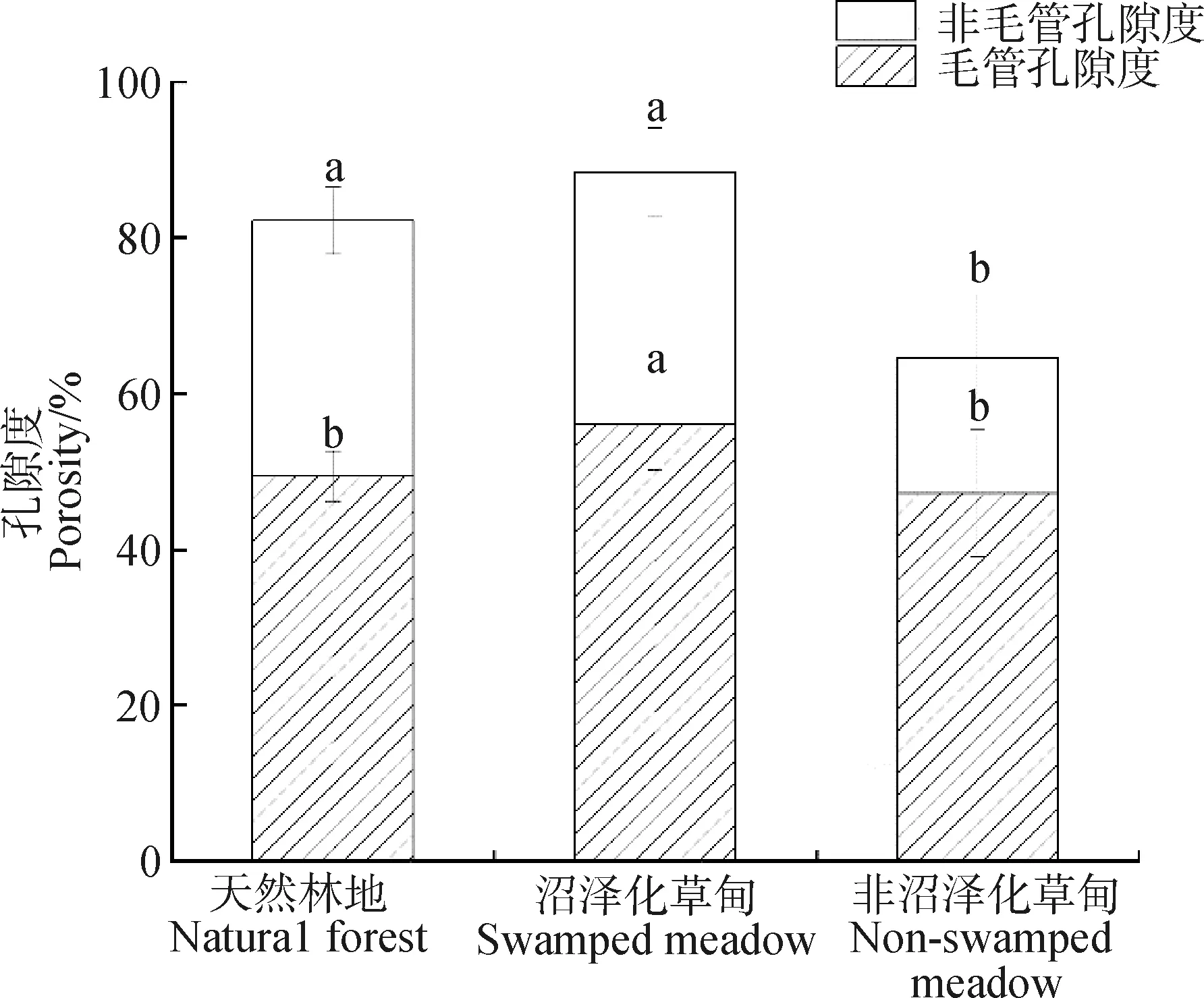
图1 若尔盖地区主要覆被类型浅层土壤孔隙度
雨季中期,不同覆被类型浅层土壤最大持水量具有显著差异(P<0.05,图2),沼泽化草甸最大,达到2 465.40 g·kg-1,其次为天然林地(1 464.66 g·kg-1),而非沼泽化草甸(613.52 g·kg-1)土壤最大持水量较低。不同覆被类型浅层土壤毛管持水量和田间持水量与最大持水量大小顺序类似。沼泽化草甸土壤最大持水量、毛管持水量和田间持水量约为天然林地的1.7倍,约是其他3类草甸的4.0倍。

图2 若尔盖地区主要覆被类型浅层土壤持水量
2.2 主要覆被类型浅层土壤有机碳含量及相关性
不同覆被类型浅层土壤有机碳含量差异显著(P<0.05,图3),沼泽化草甸>天然林地>非沼泽化草甸,沼泽化草甸土壤有机碳含量约为约为非沼泽化草甸和天然林地的4.2和2.7倍。相关性分析发现,土壤容重、有机碳含量、孔隙度、毛管持水量、田间持水量与土壤最大水量间相关性显著(P<0.01),土壤容重、孔隙度、最大持水量与土壤有机碳含量间相关性显著(P<0.01,表2)。
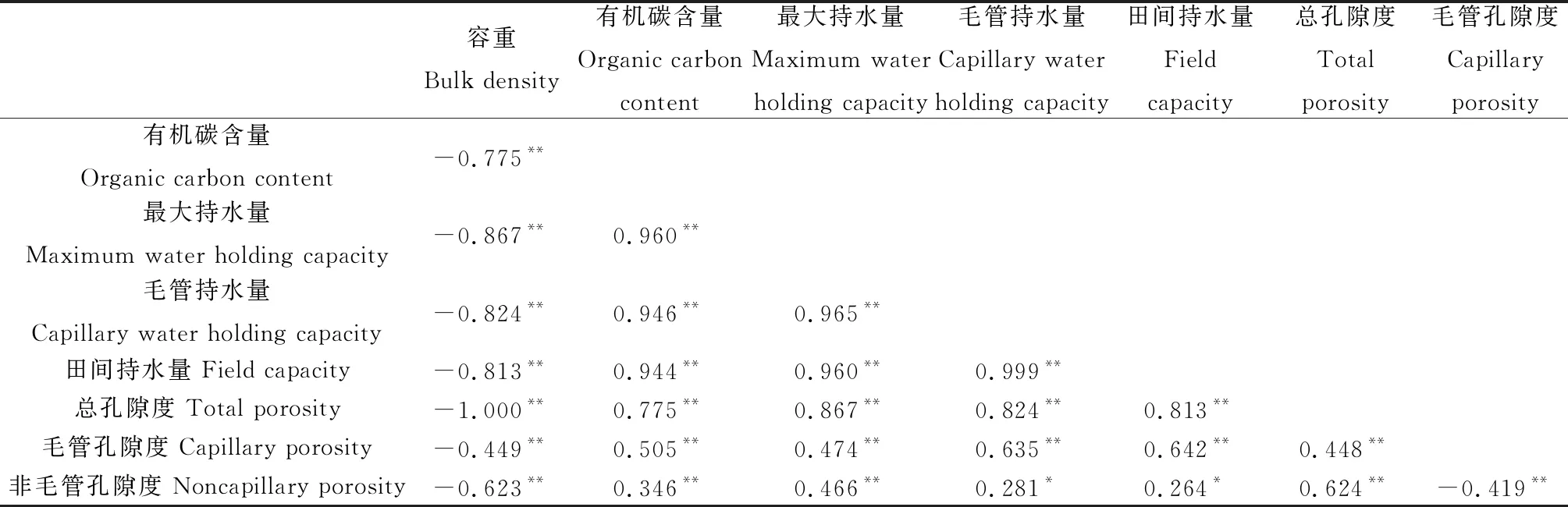
表2 若尔盖地区浅层土壤持水特征和有机碳相关性

图3 若尔盖地区主要覆被类型浅层土壤有机碳含量
2.3 主要覆被类型土壤持水量和有机碳含量
总体上(图4a),浅层土壤最大持水量与有机碳含量呈极显著正相关(R2=0.92,P<0.01,n=94),3种覆被类型浅层土壤两者之间也有类似关系(R2=0.74~0.89,P<0.01,图4b~d)。浅层土壤最大持水量和有机碳含量拟合曲线的斜率k可以反映土壤最大持水量对有机碳含量变化的敏感程度,k值越大,表示土壤最大持水量对有机碳含量变化越敏感。本研究区k值大小顺序为:天然林地(7.43)>非沼泽化草甸(5.49)>沼泽化草甸(4.11),表明天然林地土壤最大持水量对有机碳含量变化最敏感,而沼泽化草甸水碳关系相对稳定。总体上土壤最大持水量和有机碳含量的拟合曲线斜率为5.79,介于沼泽化草甸和天然林地拟合曲线斜率之间,且与非沼泽化草甸接近。

图4 若尔盖地区浅层土壤最大持水量与有机碳含量曲线拟合
3 讨论
3.1 主要覆被类型浅层土壤持水和有机碳特征及其相关关系
土壤持水能力主要受土壤总孔隙度、毛管孔隙度、容重、有机质、生物量、枯落物、腐殖质层等的影响[11]。本研究天然林地浅层土壤总孔隙度显著高于非沼泽化草甸(图1),可能是因为研究区天然林地通常位于坡顶或陡坡地段,土壤粒径较粗,石砾含量也高,因此孔隙度较高,尤其是非毛管孔隙度(图1)。有机碳与土壤颗粒相结合,形成不同粒级的团聚体,增加了孔隙度、改善土壤结构,进而提高土壤的持水量[17],使天然林地土壤持水量显著大于非沼泽化草甸土壤持水量(图2)。天然林地和沼泽化草甸土壤总孔隙度差距不大,总孔隙度和最大持水量呈现极显著正相关关系,但是天然林地和沼泽化草甸最大持水量却出现了1.7倍的差距,这有可能是因为:(1)若尔盖地区沼泽化草甸土壤有机碳含量显著高于天然林地(图3),土壤有机碳含量越高,持水能力越强[18-19];(2)大量研究认为,浅层土壤持水能力受控于土壤容重,而土壤有机碳含量与土壤容重之间存在显著的负相关关系[20-21],与本研究结果一致(表2),这也间接的反映若尔盖地区天然林地相对于沼泽化草甸土壤更为紧实,不利于土壤水保持;(3)由天然林地和沼泽化草甸土壤毛管孔隙度具有显著性差异造成(图1),然而由于缺乏土壤毛管孔隙度和持水量相关分析数据,该推测仍需进一步分析论证。森林生态系统结构复杂,加之针叶凋落物分解缓慢,大量养分元素累积于枯枝落叶,而沼泽化草甸会季节性滞水或淹水,土壤的空隙和氧气含量降低,动植物腐殖质残体处于厌氧环境,从而抑制土壤微生物呼吸与气体交换过程,土壤有机碳矿化速率减弱,使得更多有机碳积累于土壤中[22-24],这可能是本研究天然林地土壤有机碳含量显著低于沼泽化草甸的重要原因(图3)。沼泽化草甸和非沼泽化草甸由于物种组成、凋落物量、净初级生产力的差异以及放牧等因素,影响有机质种类、性质和输入量,从而导致两者土壤有机碳和水源涵养能力有所差异[25]。研究区沼泽化草甸微生物厌氧分解作用弱,有利于土壤有机碳的维持,非沼泽化草甸由于高强度放牧,牛羊牲畜反复践踏压实了浅层土壤,大幅降低了土壤孔隙度,尤其是非毛管孔隙度(图1)。此外,非沼泽化草甸鼠兔害较沼泽化草甸严重,鼠兔类动物啃食地下根系,加之牲畜啃食地上植物,使得进入土壤的枯落物减少,导致非沼泽化草甸浅层土壤有机碳含量显著低于沼泽化草甸(图3)。因此,应该适度放牧,这样才能维持高寒草甸土壤养分和水源涵养能力[26]。
本研究中,浅层土壤最大持水量与容重、孔隙度和有机碳含量显著相关(表2),因为土壤孔隙度和容重对有机质高度依赖,有机质很大程度影响了土壤持水能力,而土壤碳积累和矿化也离不开水的参与,因此土壤持水能力与碳固定是相互作用、相互影响[16]。Manns等[14]提出了线性模型解释土壤有机碳和含水量之间的结合趋势,并用偏最小二乘回归、主成分分析和地统计学半变异函数检验了土壤有机碳和土壤水碳的线性关系。若尔盖地区主要覆被类型浅层土壤最大持水量和有机碳含量表现出良好的线性关系(图4),这也证实了在若尔盖高寒生态系统中存在这种水碳相互作用关系。若尔盖高寒地区年降水量较大,沼泽化草甸土壤会出现季节性滞水。过高的土壤水分含量会抑制土壤有机质矿化[22],使土壤有机碳积累,而有机碳会促进土壤稳定团聚体的形成,产生大量中孔和微孔,减少大孔隙的丰度,从而改善土壤孔性和结构,容纳更多毛细管水和吸湿水,又提高了土壤持水能力[27],促进了土壤水碳关系良性循环和生态功能提升演变。这种良性的正反馈现象,Farley等[12]在对帕拉莫草原上蒙特利松林分土壤研究时也得出类似结论。研究发现,向土壤中添加生物质炭能够显著降低土壤容重、增加土壤孔隙度和饱和含水量等,从而提高土壤持水能力[28-29]。
3.2 浅层土壤最大持水量和有机碳含量关系
若尔盖地区浅层土壤最大持水量与有机碳含量拟合曲线斜率较大,且斜率差异明显(图4),区域水平上拟合曲线斜率为5.79(图4a),与非沼泽化草甸拟合曲线斜率(5.49)接近(图4d)。而分覆被类型其拟合曲线斜率在4.11~7.43之间,这意味着如果气候或环境变化引起了土壤有机碳含量每变化1个单位,土壤最大持水量会相应变化4.11~7.43个单位,表明土壤持水量变化幅度远高于有机碳含量的变化幅度。类似地,Gould[30]通过对美国密歇根州农场土壤的研究也得出类似结论。
由于沼泽化草甸土壤季节性滞水或淹水,使得土壤有机质腐殖化程度高(矿化程度低)且性质相对稳定均一[22],这可能导致与土壤持水能力相关的土壤孔性和团聚体结构等理化性状相对平衡,因而沼泽化草甸水碳关系相对稳定。这也暗示,在气候或环境变化导致土壤碳变化时,沼泽化草甸土壤持水能力的变化相对稳定。但是倘若沼泽化草甸积水环境减弱,可能会演变成非沼泽化草甸类型,进而显著降低土壤有机碳含量和持水能力(图3,图4b)。研究区天然林地通常位于坡顶或陡坡,土壤粒径较粗,石砾含量也高,因此与土壤持水能力相关的土壤孔性和结构高度依赖于有机质含量和性质[16],因而天然林地土壤最大持水量对有机碳含量变化最敏感。此外,天然林地土壤有机组分中疏水性强的胡敏酸和富里酸含量较高[31],在若尔盖高寒地区天然林地土壤长期浸润背景下,土壤入渗速率随有机质含量提高而逐渐增大,其持水量变化明显[32]。这也暗示,在气候或环境变化导致土壤碳变化时,天然林地土壤持水能力的变化非常剧烈。若尔盖地区非沼泽化草甸载畜量大、鼠兔害较严重,局部草场草种较差、退化裸露,浅层土壤有机物输入量有限,这可能导致与土壤持水能力相关的土壤孔性和团聚体结构等理化性状相对较差[26],因而非沼泽化草甸浅层土壤水碳关系相较于沼泽化草甸不稳定。这也暗示,在气候或环境变化导致土壤碳变化时,非沼泽化草甸浅层土壤持水能力可能会有较明显的变化。
本研究结果表明,土壤持水量和有机碳含量关系在若尔盖高寒林草生态系统稳定性中具有重要参考意义,其曲线拟合斜率宜纳入高寒林草系统生态模型,为科学判断全球变化背景下区域碳水功能评估提供数据基础。
4 结论
若尔盖地区3种主要覆被类型中,沼泽化草甸浅层土壤持水能力和固碳能力均最强,其次为天然林地,非沼泽化草甸持水固碳能力较弱。若尔盖地区浅层土壤最大持水量与有机碳含量呈显著正相关关系,其拟合曲线斜率在4.11~7.43之间,以天然林地最大,沼泽化草甸最小,表明天然林地土壤最大持水量对有机碳含量的变化最敏感,沼泽化草甸水碳关系较稳定。
猜你喜欢
中南林业科技大学学报(2022年8期)2022-09-24
农业工程学报(2022年8期)2022-08-08
北京大学学报(自然科学版)(2022年3期)2022-06-17
绿色科技(2022年4期)2022-03-21
大众文艺(2021年13期)2021-07-31
海外文摘·文学版(2021年3期)2021-05-19
西部资源(2018年3期)2018-11-06
青年文学家(2016年34期)2017-03-31
延河·绿色文学(2016年8期)2016-05-14
