
草地贪夜蛾三个SfruPBPs对本种及同域种劳氏粘虫性信息素及腺体组分的亲和力
2022-04-19 13:41郑树壕司玉晓郭慧芳董双林
昆虫学报 2022年3期
郑树壕, 司玉晓, 闫 祺, 郭慧芳, 董双林,*
(1. 南京农业大学植物保护学院, 农作物生物灾害综合治理教育部重点实验室, 南京 210095;2. 江苏省农业科学院植物保护研究所, 南京 210014)
草地贪夜蛾Spodopterafrugiperda属鳞翅目(Lepidoptera)夜蛾科(Noctuidae),具有很强的迁飞扩散能力,但长期以来局限于在美国、墨西哥、巴西等中美洲地区发生和危害(Pashleyetal., 1985; Pashley and Martin, 1987; Andrews, 1988; Cruzetal., 1999)。直到2016年,草地贪夜蛾在非洲被发现,并很快蔓延到欧洲和亚洲(Goergenetal., 2016; Feldmannetal., 2019);在我国,2018年12月首次在云南发现草地贪夜蛾,次年即扩散到许多其他省份(吴秋琳等, 2019)。草地贪夜蛾属多食性害虫,主要为害禾本科的玉米和水稻,2017年在非洲玉米主产地造成53%的损失(Dayetal., 2017),2019年云南靖江市由于草地贪夜蛾入侵造成的直接经济损失达2 900万元(张琼等, 2020)。根据寄主喜好性,草地贪夜蛾分为玉米型和水稻型(Pashleyetal., 1985; Pashley and Martin, 1987),两者在形态上高度相似,通常以细胞色素氧化酶Ⅰ(cytochrome oxidase I, COI)和磷酸甘油醛异构酶(triose phosphate isomerase, Tpi)基因序列来区分(Meagher and Gallo-Meagher, 2003; Nagoshi, 2010)。我国发生的草地贪夜蛾为玉米型,但掺杂少量水稻型的基因型(陈冬平等, 2020; 王亚如等, 2020),尚未有在田间大面积危害水稻的报道。对草地贪夜蛾发生区域和发生量的严密监测意义重大。
性信息素用于害虫监测和测报具有灵敏度高、特异性强、高效方便等优点,并已在草地贪夜蛾得到应用,但突出问题是诱芯的种特异性不高。生产上常用的二元或三元组分诱芯常常诱捕到相当比例的劳氏粘虫Leucanialoreyi(Meagheretal., 2019; 沈嘉彬等, 2019),给准确计数草地贪夜蛾虫量进而测报其种群动态带来困难。有关草地贪夜蛾性信息素的组成,迄今已从雌蛾性信息素腺体中鉴定到13种组分,但不同地理种群间有所差异(Sekul and Sparks, 1967; Tumlinsonetal., 1986; Descoinsetal., 1988; Batista-Pereiraetal., 2006; 江南纪和王琛柱, 2019; Jiangetal., 2021)。在这些组分中,Z9-14∶Ac和Z7-12∶Ac被普遍证实有田间雄蛾引诱活性(Tumlinsonetal., 1986),Z11-16∶Ac在美国宾夕法尼亚州地理种群中具有活性(Fleischeretal., 2005),生产上应用的诱芯主要由这些组分配制而成。值得注意的是,劳氏粘虫等几种粘虫外形和草地贪夜蛾相近,且性信息素组分高度重合,除共享Z9-14∶Ac和Z7-12∶Ac这2种主要行为活性组分外,12∶Ac等4种组分也在两种雌蛾腺体中并存(Sekul and Sparks, 1967; Tumlinsonetal., 1986; Descoinsetal., 1988; Hoetal., 2002; 江南纪和王琛柱, 2019; Jiangetal., 2021)。自然条件下,推测两种害虫依靠各自特有组分来达到性信息素通讯系统间的隔离。
气味结合蛋白(odorant binding protein, OBP)在昆虫性信息素等气味分子的识别和感受中起重要作用。OBP是一类水溶性小分子蛋白,高浓度存在于昆虫嗅觉感器的感器液腔中,负责识别并运输气味分子穿越感器液腔到嗅觉神经元树突膜上的气味受体(odorant receptor, OR)部位,此处气味分子被释放后单独或以气味分子-OBP复合体的形式激活OR(Leal, 2013)。在鳞翅目昆虫中,OBP分为普通气味结合蛋白(general odorant binding protein, GOBP)、信息素结合蛋白(pheromone binding protein, PBP)和触角结合蛋白(antennal binding protein, ABP)(Vogtetal., 1991, 1999)。其中,PBP在性信息素敏感的感器中表达,负责对性信息素分子的结合和运输,具有很高的配体亲和力和特异性(Vogt and Riddiford, 1981),与性信息素通讯系统的高度灵敏性和种特异性相一致。研究PBP对草地贪夜蛾及同域近缘种劳氏粘虫性信息素组分的结合能力,将有助于了解两种昆虫在性信息素通讯系统水平上的隔离机制,也可为种特异性诱芯的开发提供依据。草地贪夜蛾具有4个PBP基因,其中3个在触角中表达,暗示其在性信息素感受中起作用(刘苏等, 2020)。本研究针对这3个触角中表达的PBP基因,通过体外重组表达并纯化后,测定了PBP蛋白对草地贪夜蛾及劳氏粘虫性信息素及腺体组分的亲和力,以期为明确两种昆虫性信息素通讯间的隔离机制及开发种特异性的性信息素诱芯提供基础。
1 材料与方法
1.1 昆虫饲养与组织收取
草地贪夜蛾幼虫于2019年7月采集自江苏东台玉米田(32°45′N, 120°23′E),在室内利用甜菜夜蛾Spodopteraexigua人工饲料(黄春霞等, 2002)进行继代饲养。饲养温度为27±1℃,相对湿度为70%±5%,光周期为14L∶10D。成虫饲喂10%蜂蜜水。收取3日龄雄虫触角50对,重复3次,置于-80℃保存备用。
1.2 RNA提取和cDNA第1链的合成
使用Trizol Reagent (Invitrogen, 美国)提取总RNA。 RNA的质量和浓度由NanoDrop2000(Thermo Scientific, 美国)检测,OD260/OD280比值均为1.8~2.0。根据试剂盒说明书,使用HiScript®III RT SuperMix (诺唯赞, 南京)从1 μg总RNA中合成cDNA第1链,-20℃保存备用。
1.3 PBP基因原核表达载体的构建
根据NCBI上草地贪夜蛾PBP1, PBP2和PBP3的cDNA序列(GenBank登录号分别为MN541407, MN541408和MN541409),去除信号肽编码序列后设计上下游引物,用于PBP基因的克隆;去除信号肽序列后,设计包含BamHⅠ和XhoⅠ限制性内切酶位点的同源重组引物(表1),用于基因的原核表达。引物由南京金斯瑞公司合成。

表1 引物信息
以成虫触角cDNA为模板,采用高保真性DNA聚合酶2×Phanta Max Master Mix(诺唯赞,南京)按照说明书进行PCR扩增。PCR反应体系(25 μL): 2×Phanta Max Master Mix 12.5 μL,上下游引物(10 μmol/L)各1 μL, cDNA模板1 μL, ddH2O 9.5 μL。反应条件: 95℃预变性3 min; 95℃变性15 s, 55℃退火15 s, 72℃延伸40 s, 35个循环; 72℃总延伸5 min。使用1.2%凝胶电泳检测PCR产物并使用Axyprep DNA凝胶回收试剂盒(康宁生命科学,苏州)进行胶回收。PCR产物回收后连接到pEASY-Blunt3载体(全式金,北京)并转化到大肠杆菌EscherichiacoliTrans1-T1感受态细胞(全式金,北京)中,提取质粒DNA并测序验证。将含有目的基因片段的重组质粒DNA和pET-30a(+)质粒(Novagen,德国)DNA分别进行双酶切(BamHⅠ和XhoⅠ)(Fermentas,加拿大),然后将目的片段连接到pET-30a(+)载体,再转化到Trans1-T1感受态细胞中。用含卡那霉素的LB平板进行筛选,挑取阳性克隆进行测序。
1.4 PBP的诱导表达与纯化
PBP的诱导表达与纯化参考已报道的方法(Zhangetal., 2021)。将1.3节经测序验证后的质粒转入大肠杆菌BL21(DE3)感受态细胞中,接种于LB 培养基后培养至 OD600= 0.6~0.8 后加入 IPTG(终浓度为1 mmol/L)进行PBP诱导表达。诱导后菌液经超声波破碎并用SDS-PAGE检测。将含有包涵体蛋白的溶液通过Ni-NTA琼脂糖磁珠(Ni磁珠)(金斯瑞,南京)进行纯化,得到带有His标签的融合蛋白。为防止His标签对荧光竞争结合实验结果造成影响,使用肠激酶(金斯瑞,南京)在22℃条件下反应10~14 h切除His标签。
1.5 荧光竞争结合实验
共测试14种气味物质,均为草地贪夜蛾或劳氏粘虫性信息素腺体浸提液组分,其中E7-12∶Ac,Z9-12∶Ac, 11-12∶Ac,Z10-14∶Ac,Z11-14∶Ac,Z9-14∶Ald和Z11-16∶Ald为草地贪夜蛾特有组分,Z7-14∶Ac为劳氏粘虫特有组分,其他为共有组分(Sekul and Sparks, 1967; Tumlinsonetal., 1986; Descoinsetal., 1988; Hoetal., 2002; 江南纪和王琛柱, 2019; Jiangetal., 2021)(表2)。这14种气味物质中,Z9-14∶Ac和Z7-12∶Ac已证实对两种害虫具有田间雄蛾引诱活性(Tumlinsonetal., 1986),因此是性信息素组分;其他组分在田间诱捕试验中没有发现明显的雄蛾引诱活性,本文中称为“腺体组分”,但不排除其中的某些组分可能在近距离内促进两性间交配而起到性信息素的作用。本研究所使用的14种气味物质中,Z11-14∶Ac和16∶Ac购自沈阳北欣贸易有限公司,另外12种气味物质均购自常州宁录生物科技有限公司。荧光竞争结合实验测定方法同实验室前期报道(Liuetal., 2012; 魏丹等, 2013)。首先检测PBP 蛋白对荧光探针1-NPN的结合能力。将PBP重组蛋白溶液加入50 mmol/L的Tris-HCl缓冲液中,总体系为250 μL,使蛋白的终浓度为2 μmol/L,按2, 4, 6, 8, 10, 12, 14, 16, 18和20 μmol/L的终浓度梯度加入1-NPN(浓度为0.5 mmol/L),每次反应2 min,在337 nm激发,记录荧光强度,计算蛋白与1-NPN的结合常数Ki, 然后利用竞争结合实验测定气味物质和PBP重组蛋白的结合能力。在竞争结合实验中, PBP重组蛋白和1-NPN的浓度均为2 μmol/L,反应时间为2 min,在337 nm激发,记录荧光值(初始荧光值),然后被测气味物质(浓度0.05 mmol/L)按0.2, 0.4, 0.6, 0.8, 1.0, 1.2, 1.6, 2.0, 2.4, 2.8, 3.2, 3.6和4.0 μmol/L的终浓度梯度加入到PBP重组蛋白与荧光探针的混合溶液中,反应2 min后记录荧光值。实验在25℃条件下进行。利用公式Ki=IC50/(1+[1-NPN]/K1-NPN)计算各气味物质的结合常数,其中[1-NPN]为未结合的1-NPN浓度,K1-NPN为1-NPN与PBP的结合常数。利用GraphPad Prism 6.0软件分析PBP与1-NPN的结合实验数据。
为叙述方便,本文将PBP和气味物质间结合能力按Ki<1.00 μmol/L, 1.00
2 结果
2.1 草地贪夜蛾3个SfruPBPs的诱导表达与纯化
经过IPTG诱导表达10 h后,收集菌液离心得到菌体后进行超声波破碎,破碎液离心后经SDS-PAGE分析发现,目的蛋白主要以包涵体的形式存在于沉淀中,在上清中很少。对带有His标签的目的蛋白进行纯化,大小为21 kD左右。为防止His标签对实验造成的影响,对融合蛋白进行肠激酶酶切,去除His标签。去除标签后3个SfruPBPs大小均在16 kD左右,与预测的3个SfruPBPs的分子量一致(图1)。

图1 草地贪夜蛾3个SfruPBPs重组蛋白的表达与纯化
2.2 草地贪夜蛾3个SfruPBPs与不同气味物质的结合能力
结果表明,3个SfruPBPs溶液的荧光值随1-NPN浓度的增加而增加并趋于饱和;SfruPBP1, SfruPBP2和SfruPBP3与1-NPN的结合常数(K1-NPN)分别为4.61±1.0, 9.05±1.1和3.72±0.7 μmol/L(图2)。因此,1-NPN可以作为探针用于后续的竞争性结合实验。

图2 3个SfruPBPs重组蛋白与1-NPN的结合曲线
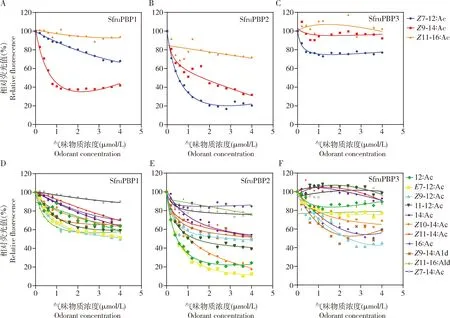
利用荧光竞争结合实验测定了3个SfruPBPs对14种气味物质的结合能力,发现3个SfruPBPs对气味物质的结合谱明显不同。SfruPBP1仅对草地贪夜蛾性信息素主组分Z9-14∶Ac结合并具有强结合能力(Ki=0.80 μmol/L);SfruPBP3则仅对腺体组分Z9-12∶Ac具有中等结合能力(Ki=1.36 μmol/L);而SfruPBP2对包括性信息素次要组分Z7-12∶Ac在内的7种组分具有结合能力,其中对Z7-12∶Ac, 12∶Ac,E7-12∶Ac和Z10-14∶Ac具有强结合能力(Ki<1.00 μmol/L),对Z9-14∶Ac,Z9-12∶Ac和11-12∶Ac有中等结合能力(1.00 μmol/L 本研究测定了草地贪夜蛾3个SfruPBPs对草地贪夜蛾及劳氏粘虫性信息素或性信息素腺体组分共14种气味物质的结合特性,发现草地贪夜蛾3个SfruPBPs均可结合本种的性信息素或腺体组分,但不同PBP的配体谱明显不同;此外,3个SfruPBPs对劳氏粘虫腺体特有组分Z7-14∶Ac均无明显结合能力(图3; 表2)。研究结果为进一步明确草地贪夜蛾对性信息素的感受机制,以及草地贪夜蛾与劳氏粘虫性信息素通讯系统间的隔离机制提供了重要基础。 图3 3个SfruPBPs重组蛋白与14种气味物质的竞争结合曲线 表2 草地贪夜蛾3个SfruPBPs重组蛋白对本种及劳氏粘虫性信息素和其他腺体组分的亲和力 就腺体内含量及田间雄蛾引诱活性而言,Z9-14∶Ac和Z7-12∶Ac分别是草地贪夜蛾性信息素的主组分和次要组分,常以不同比例构成二元诱芯用于田间害虫测报和诱捕(Tumlinsonetal., 1986; Unbehendetal., 2014; Jiangetal., 2021)。在被测14种气味中, SfruPBP1对性信息素主组分Z9-14∶Ac特异性强结合(Ki=0.80 μmol/L)(表2);SfruPBP2则对性信息素次要组分Z7-12∶Ac强结合,并对主组分具中等结合能力;SfruPBP3对2种性信息素组分均无明显结合能力(图3;表2)。在个别草地贪夜蛾地理种群中,Z11-16∶Ac也被证明具有一定的田间诱捕活性(Fleischeretal., 2005),但本研究发现3个SfruPBPs对该组分均没有明显的结合能力,这与在大部分研究中该组分不具田间行为活性的结论(Tumlinsonetal., 1986; Unbehendetal., 2014)是一致的。结合刘苏等(2020)对草地贪夜蛾3个PBPs的组织表达分析结果,我们推测SfruPBP1和SfruPBP2在雄虫识别性信息素中起主要作用,SfruPBP3起辅助作用。 草地贪夜蛾与斜纹夜蛾Spodopteralitura和甜菜夜蛾为同属近缘种,其中后两者均为我国重要农业害虫,比较3种昆虫PBP同源基因的表达模式及对性信息素组分的选择性差异,可以为PBP基因的进化及功能分化研究提供线索。在入侵种草地贪夜蛾中,3个PBPs对性信息素主组分Z9-14∶Ac的结合能力顺次减弱(SfruPBP1>SfruPBP2>SfruPBP3)(图3;表2),与斜纹夜蛾和甜菜夜蛾3个PBPs对各自主组分Z9,E11-14∶Ac或Z9,E12∶Ac结合能力的结果(Liuetal., 2012, 2013)类似。但不同的是,草地贪夜蛾SfruPBP1特异性结合主组分Z9-14∶Ac,对不同组分具明显的选择性,而斜纹夜蛾和甜菜夜蛾的3个PBPs虽然对性信息素组分的结合能力顺次减弱,但相同PBP在不同组分间没有明显的选择性(Liuetal., 2012, 2013),这种差异的原因需要进一步探索。比较发现,草地贪夜蛾3个PBPs在雌雄虫触角中的表达模式与斜纹夜蛾和甜菜夜蛾也存在不同。根据刘苏等(2020)结果,草地贪夜蛾PBP1和PBP2均为雄虫触角高表达(雄/雌分别约为2.8和3.7),且在雄蛾触角中PBP2的表达量高于PBP1(PBP2/PBP1=1.3)。而在斜纹夜蛾和甜菜夜蛾中,仅PBP1为雄虫触角高表达(雄/雌分别为6.0和2.7),且PBP1的表达量显著高于PBP2(PBP1/PBP2分别为3.0和11.0)(Liuetal., 2012, 2013)。表达模式的不同可能与其PBP的配体选择性的差异相关。 在使用草地贪夜蛾性信息素二元(Z9-14∶Ac和Z7-12∶Ac)或三元(Z9-14∶Ac,Z7-12∶Ac和Z11-16∶Ac)不同比例的诱芯进行田间雄蛾诱捕时,常常能同时诱捕到相当比例的劳氏粘虫(Meagheretal., 2019; 沈嘉彬等, 2019),但自然情况下两种害虫并不会交叉吸引,说明某些种特异性的性信息素次要组分在起作用。结合现有文献报道(Hoetal., 2002; 江南纪和王琛柱, 2019),在雌蛾性信息素腺体组分中,除6种共有组分外,草地贪夜蛾还特有7种组分(E7-12∶Ac,Z9-12∶Ac, 11-12∶Ac,Z10-14∶Ac,Z11-14∶Ac,Z9-14∶Ald和Z11-16∶Ald),而劳氏粘虫特有2种组分(Z7-14∶Ac和Z7-16∶Ac)。本研究对这些特有组分(Z7-16∶Ac除外)的测定结果表明,草地贪夜蛾SfruPBP对本种特有组分E7-12∶Ac,Z9-12∶Ac, 11-12∶Ac和Z10-14∶Ac有不同程度的结合能力,而对劳氏粘虫特有组分Z7-14∶Ac无明显结合能力(表2),说明这些组分均可能在两种害虫性信息素通讯隔离中起作用。此外,Ho等(2002)报道,劳氏粘虫雄蛾对其2个特有组分Z7-14∶Ac和Z7-16∶Ac均有明显的触角电位反应,也说明这2种组分可能在种间隔离中发挥作用。进一步分析劳氏粘虫PBP对两种害虫共有及特有组分的结合能力,将有助于更全面认识两种害虫性信息素通讯间的隔离机制,并为开发两种害虫的特异性田间诱芯提供指导。3 讨论

猜你喜欢
今日农业(2022年13期)2022-09-15
昆虫学报(2022年4期)2022-05-19
波谱学杂志(2021年3期)2021-09-07
儿童故事画报·智力大王(2020年3期)2020-04-26
中国医科大学学报(2020年4期)2020-01-08
水上消防(2019年6期)2019-12-02
今日农业(2019年12期)2019-08-15
今日农业(2019年14期)2019-01-04
今日农业(2019年13期)2019-01-03
作物研究(2017年2期)2017-06-01
