
棉大卷叶螟求偶行为节律和性信息素鉴定
2022-04-19 13:37朱俣伟司玉晓邓建宇董双林
昆虫学报 2022年3期
朱俣伟, 司玉晓, 邓 颖, 李 瑜, 邓建宇, 董双林, 闫 祺,*
(1. 南京农业大学植物保护学院, 南京 210095; 2. 浙江农林大学现代农学院, 杭州 311300)
棉大卷叶螟Syleptaderogata属鳞翅目(Lepidoptera)螟蛾科(Pyralidae),在我国除宁夏、青海、新疆等地区未见报道外,其余省区均有分布(陆佩玲等, 2008),以幼虫吐丝缀叶危害棉花、秋葵、冬寒菜、蜀葵、朱槿等农林植物。大发生时一片叶上可达数个叶卷,严重时吃光所有叶片,仅留枝、茎,影响作物产量与质量。随着转Bt基因抗虫棉的普遍种植,棉田杀虫剂使用量大幅下降,使得棉大卷叶螟在棉花上的发生和危害呈上升趋势(陈建等, 2008)。
目前,棉大卷叶螟的防控主要依靠化学防治,但由于棉大卷叶螟幼虫有卷叶取食的习性,降低了与化学药剂的接触率,防治效果并不理想,且长期使用化学药剂,极易带来环境污染,生态系统破坏,最终危害人类健康,为此,探索开发新型绿色环保的害虫防治方法十分必要。昆虫性信息素在害虫治理上具有安全高效、无污染、专一性强等优点,有着广泛的应用前景,而昆虫性信息素产品开发应用的基础是昆虫性信息素化学组分结构的鉴定。至今为止,全世界己鉴定的昆虫性信息素或类似物超1 000种,我国投入使用的性信息素产品有100多种(El-Sayed, 2020),如:徐善忠等(2019)利用有效成分为Z11-16∶Ald和Z13-18∶Ald的混合制剂干扰稻纵卷叶螟Cnaphalocrosismedinalis和二化螟Chilosuppressalis的交配,防治效果显著;程英等(2019)利用斜纹夜蛾Prodenialitura性诱剂对贵州地区辣椒斜纹夜蛾进行虫情预测,及时准确地反映贵州不同地区辣椒斜纹夜蛾的种群消长情况并作出预报预测。
目前国内外对于棉大卷叶螟的研究多为形态特征、生物学特性、吐丝机理等方面。虽然张清泉(2011)利用棉大卷叶螟雌蛾性腺浸提液在野外成功诱捕到一定数量的棉大卷叶螟,但关于棉大卷叶螟性信息素鉴定方面的研究尚未见文献报道。鉴于性信息素应用于防治害虫防治的诸多优点,本文将重点对棉大卷叶螟雌蛾的求偶行为及其动态、性信息素组分的鉴定开展研究,以期为开发棉大卷叶螟性信息素诱芯并应用于田间种群的高效监测和绿色防控提供依据和指导。
1 材料与方法
1.1 供试昆虫
棉大卷叶螟幼虫采集自南京郊区的棉花Gossypiumspp.田或秋葵Abelmoschusesculentus田。试虫采回饲养于养虫室内,温度为25±1℃,相对湿度为60%±10%,光周期为14L∶10D。初孵幼虫饲养于培养皿中(直径7 cm),每个培养皿放入1~2片秋葵、冬葵寄主叶片,以及约15头幼虫,每2 d换一次新鲜叶片。待幼虫生长至3龄后,转移到饲养盒(10 cm×20 cm×5 cm)中,继续用新鲜寄主叶片饲养,每盒10头左右,直至幼虫化蛹。之后将蛹取出,区分雌雄后,分别置于饲养盒(10 cm×20 cm×5 cm)中待其羽化。每日光期开始后检查并取出羽化的成虫,按照1∶1的雌雄比例置于交配笼(13 cm×25 cm×11 cm)中交配,每个交配笼中放10对,成虫用10%的蜂蜜水补充营养。
1.2 棉大卷叶螟求偶行为及其动态节律
将羽化当日的雌蛾单头放入已编号的塑料杯(杯底直径6 cm、杯口直径9 cm、杯高12 cm)中,塑料杯底部中央放置浸有10%蔗糖溶液的脱脂棉球,将培养皿倒扣塑料杯。从暗期开始前0.5 h到暗期结束后0.5 h,每隔30 min观察记录1次雌蛾的求偶情况,以棉大卷叶螟雌蛾腹部末端产卵器持续伸出或者产卵器有规律地伸缩为标准(王焱等, 2009)。观察时用纱布遮挡红光灯灯源,以避免强光干扰雌蛾的求偶行为。从下一个暗期记为1日龄开始,连续观察7 d,共观察20头处女雌蛾。
1.3 棉大卷叶螟雌蛾性信息素的提取
选取3-4日龄未交配的棉大卷叶螟雌蛾,在暗期开始后4-5 h之间提取性信息素腺体。用手轻轻挤压雌蛾腹部,迫使其性信息素腺体完全伸出腹部(图1),用解剖剪将腺体剪下,放入含有约1 mL正己烷的样品瓶中,每瓶放入5个腺体,使腺体完全浸泡在正己烷中,静置30 min后取出腺体。将6个样品瓶的浸提液分别过滤合并后,用氮气浓缩至1 mL左右,密封并储存在-20℃冰箱中备用。

图1 棉大卷叶螟雌蛾性信息素腺体
1.4 棉大卷叶螟雌蛾性腺浸提液的GC-EAD分析
气相色谱-触角电位联用仪(GC-EAD)所用GC为安捷伦7890B,色谱柱为HP-5MS(30 m×0.25 mm, 0.25 μm; J&W Scientific, Folsom, CA, USA),分流器(OSS-2, SGE, 澳大利亚)接出口端,1∶1分流,一部分进入FID氢火焰离子检测器,另一部分经过加热套管加热通过洁净的气流吹向固定在EAG装置上的雄蛾触角。昆虫触角电位仪(EAG)为德国Syntech公司生产,采用IDAC-4信号放大器采集数据。气相色谱仪条件:检测器为FID氢火焰离子检测器,氮气作为载气,采用无分流进样,进样口温度为220℃,检测器温度260℃。GC柱箱升温程序:起始温度80℃,保持1 min,以8℃/min的速率升温至210℃,保持10 min,最后以15℃/min的速率升温至260℃,保持2 min。GC-EAD分析时,选择处于暗期3 h左右3日龄未交配的棉大卷叶螟雄蛾,将1根雄蛾的触角用解剖剪从基部剪下后,用刀片在触角两端分别切除约1 mm,利用导电胶(Spectra 360)将触角固定在电极两端,然后将电极插入EAG探头中。手动进样2 μL性腺浸提液(约为3头雌蛾当量,3 FE),经分流后一部分进入FID氢离子检测器,另一部分经过加热套管加热吹向雄蛾触角,记录到的触角电位通过信号放大器IDAC-4采集数据,使用EAG Pro软件同步记录GC图谱和EAG图谱。
1.5 棉大卷叶螟雌蛾性腺浸提液的GC-MS分析
气相色谱-质谱联用仪(GC-MS)所用GC为安捷伦7890A串联安捷伦MS 5977B,GC-MS中的色谱柱与GC-EAD相同,以氮气为载气。电离方式为电子轰击电离(EI),电离能量为70 eV,离子源发生器温度为220℃,质量扫描范围为m/z33~400 amu。GC柱箱升温程序:起始温度80℃,保持1 min,以8℃/min的速率升温至210℃,保持10 min,最后以15℃/min的速率升温至250℃保持2 min。采用手动进样,每次进样量为2 μL(约3 FE)。通过对比GC-EAD图谱和GC-MS总离子流图(TIC)的峰形和保留时间(RT),分析候选化合物的特征离子碎片,匹配仪器联机的Nist14数据库中的标准化合物质谱图来确定性信息素组分的具体化学结构。
1.6 棉大卷叶螟雌蛾性腺浸提液和10,12-16∶Ald混合标准样品的GC分析
分析10,12-16∶Ald的4种同分异构体标准品混合溶液(含E10,E12-16∶Ald,E10,Z12-16∶Ald,Z10,Z12-16∶Ald和Z10,E12-16∶Ald 4种组分)所用GC为赛里安456-GC。气相色谱条件:检测器为FID氢火焰离子检测器;各性信息素组分的分离依靠BR-5石英毛细管色谱柱,进样口温度为220℃,检测器温度为250℃,以氮气作为载气,采用不分流手动进样。GC柱箱升温程序:起始温度60℃,保持2 min,以3℃/min的速率升温至180℃,最后以10℃/min的速率升温至250℃保持5 min。 Comp. II和10,12-16∶Ald的4种同分异构体的保留指数(Kovats indix, KI)的计算以色谱图上位于待测物质两侧的相邻正构烷烃的保留值为基准,用对数内插法求得。
1.7 棉大卷叶螟雄蛾触角对标准化合物的EAG分析
昆虫触角电位仪(EAG)为德国Syntech公司生产,采用IDAC-4信号放大器采集数据。分别配制E10,Z12-16∶Ald的浓度为0.01, 0.1, 1, 10和100 ng/μL的正己烷溶液,放在4℃冰箱中冷藏备用。将滤纸剪成约0.8 cm×5 cm的纸条,用移液枪将所配的E10,Z12-16∶Ald溶液以及正己烷分别在不同的滤纸条上滴加10 μL。待滤纸条上的正己烷挥发2 min后,将滤纸条放进巴氏玻璃管中。EAG分析时,选择处于暗期4 h左右3日龄未交配的棉大卷叶螟雄蛾,将雄蛾的触角用解剖剪从基部剪下后,用刀片在触角两端分别切除约1 mm,利用导电胶(Spectra 360)将触角固定在电极两端,然后将电极插入EAG探头中。 将装有滤纸的巴氏玻璃管口打开,按照空气、正己烷、浓度由低到高的E10,Z12-16∶Ald的顺序刺激触角,测出一组EAG值,以不同雄蛾的触角为重复,测得5组EAG数据。记录到的触角电位通过信号放大器IDAC-4采集数据,使用EAG Pro软件记录数据。
1.8 数据分析
求偶起止时间、持续时间以及触角电位反应等实验数据采用单因素方差分析[最小显著性差异法(least significant difference, LSD)]。 所有数据用Microsoft Excel 2016初步统计,然后用SPSS 17.0 Windows数据包分析处理,用Graphpad Prism 8软件作图。
2 结果
2.1 棉大卷叶螟雌蛾求偶行为
对20头棉大卷叶螟处女雌蛾连续观察7 d表明,雌蛾在求偶时,触角左右摆动,双翅高高抬起,快速振动,产卵器持续外伸。在暗期内不同时间段,各日龄雌蛾的求偶行为呈现明显的时动态节律;在暗期开始0.5 h,各日龄雌蛾均无求偶行为;此后个体求偶率逐渐增加,在暗期中期(4-7.5 h)达到最大,期间(5-5.5 h)出现短暂下降,总体表现为双峰型;暗期7.5 h至暗期结束,求偶率不断下降,进入光期后所有雌蛾无求偶行为(图2)。

图2 棉大卷叶螟雌蛾求偶行为节律
不同日龄雌蛾的求偶节律相似,但最高个体求偶率存在明显差异(图2)。1日龄雌蛾的求偶率最高为50%,随日龄的增加求偶率逐渐升高,至2日龄的60%并在3日龄达到最大为70%;从4日龄开始求偶率逐渐下降,至7日龄求偶率降至40%。
通过对棉大卷叶螟各日龄雌蛾的平均求偶起始时间及持续时间进行分析(图3),结果表明1日龄雌蛾的平均求偶起始时间(暗期开始后4.78 h)显著迟于3-5日龄(P<0.05),3-7日龄间的差异则不显著;各日龄雌蛾平均求偶终止时间均在暗期后的7-8 h,以4日龄雌蛾最晚,此后雌蛾平均求偶终止时间逐渐提前,但各日龄之间不存在显著性差异。从平均求偶持续时间来看,1-3日龄雌蛾随日龄增长而逐渐延长,在3日龄雌蛾求偶持续时间达到最大(3.5 h);此后,雌蛾求偶持续时间逐渐下降,至7日龄仅为1.94 h;3-4日龄雌蛾平均求偶持续时间与1日龄、7日龄雌蛾之间存在显著差异(P<0.05)。综合分析表明,3-4日龄雌蛾维持较高的求偶率和持续求偶时间,表明3-4日龄为棉大卷叶螟雌蛾求偶高峰日龄。
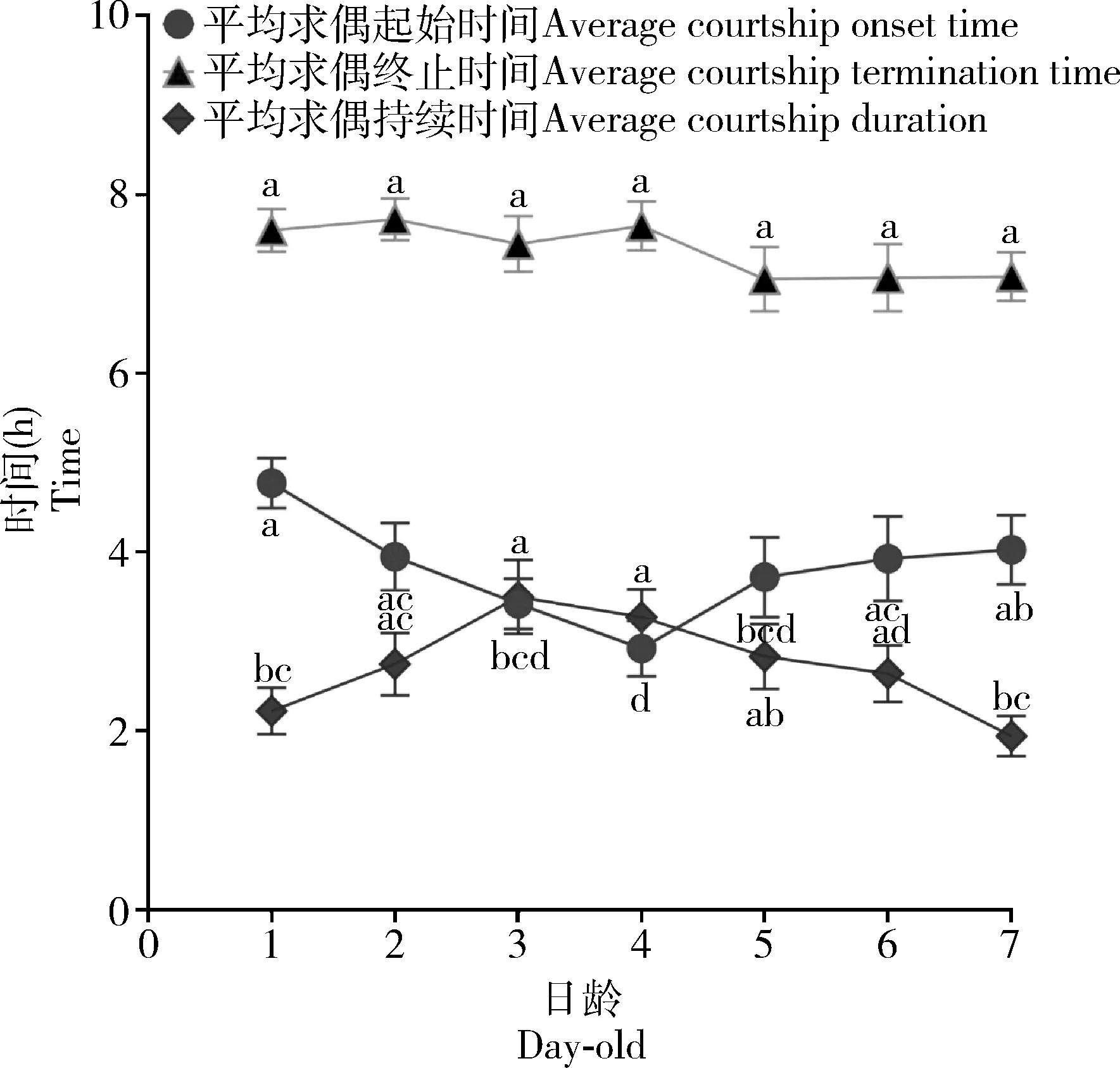
图3 不同日龄棉大卷叶螟雌蛾求偶起止及持续时间比较
2.2 棉大卷叶螟雌蛾性腺浸提液的GC-EAD分析
根据雌蛾求偶行为节律观察结果,在暗期4-5 h之间以正己烷为溶剂浸提3-4日龄棉大卷叶螟处女雌蛾的性信息素腺体,并对腺体浸提液进行GC-EAD分析,选用3日龄雄蛾触角进行实验。GC-EAD结果显示,在保留时间11.90 min以及14.45 min处存在着能引起雄蛾触角电位反应的活性物质Comp. I和Comp. II(图4),该结果具有3次以上的重复。
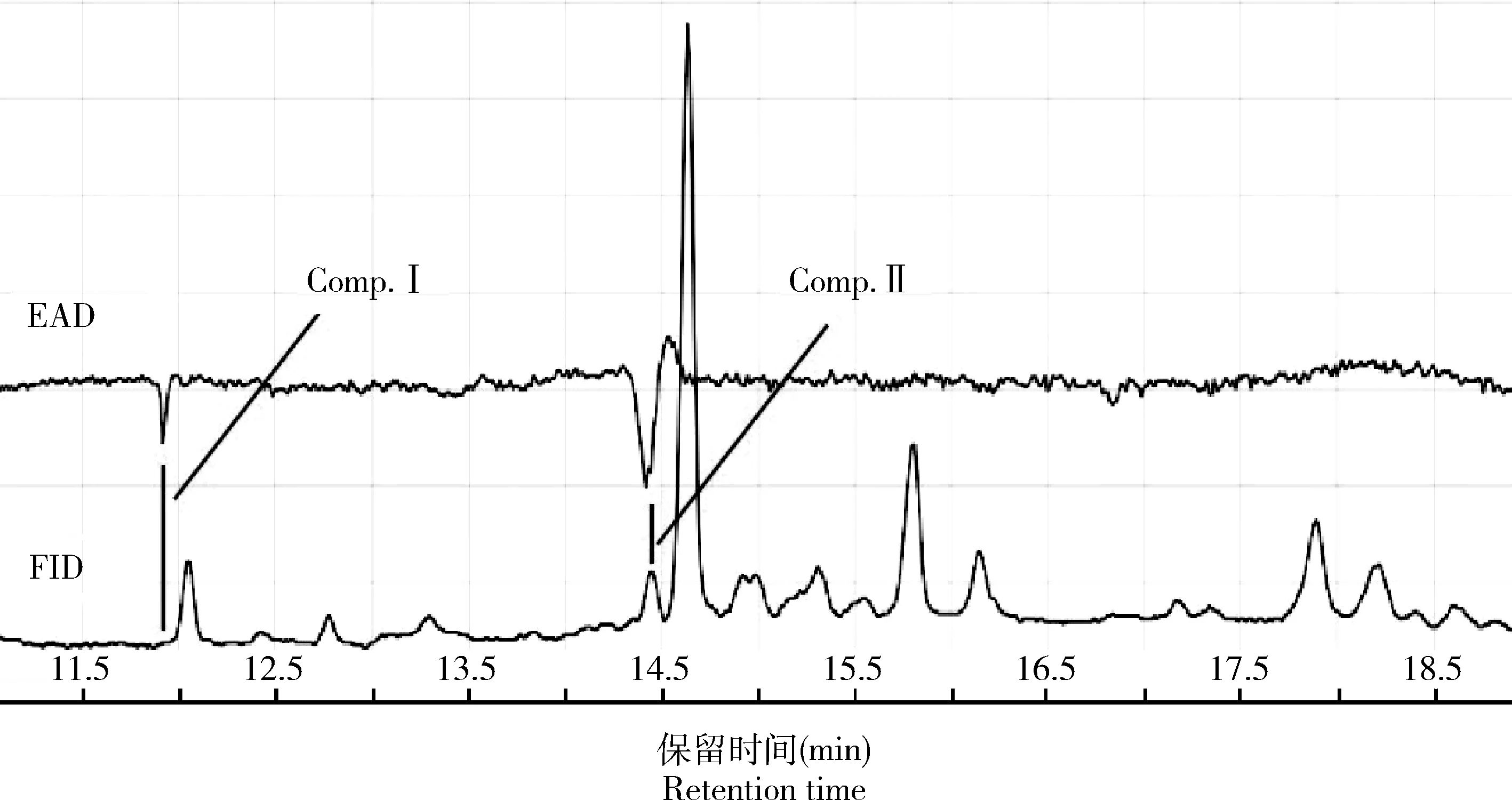
图4 棉大卷叶螟雄蛾触角对雌蛾性信息素腺体浸提液的GC-EAD分析图谱
2.3 棉大卷叶螟雌蛾性腺浸提液的GC-MS分析
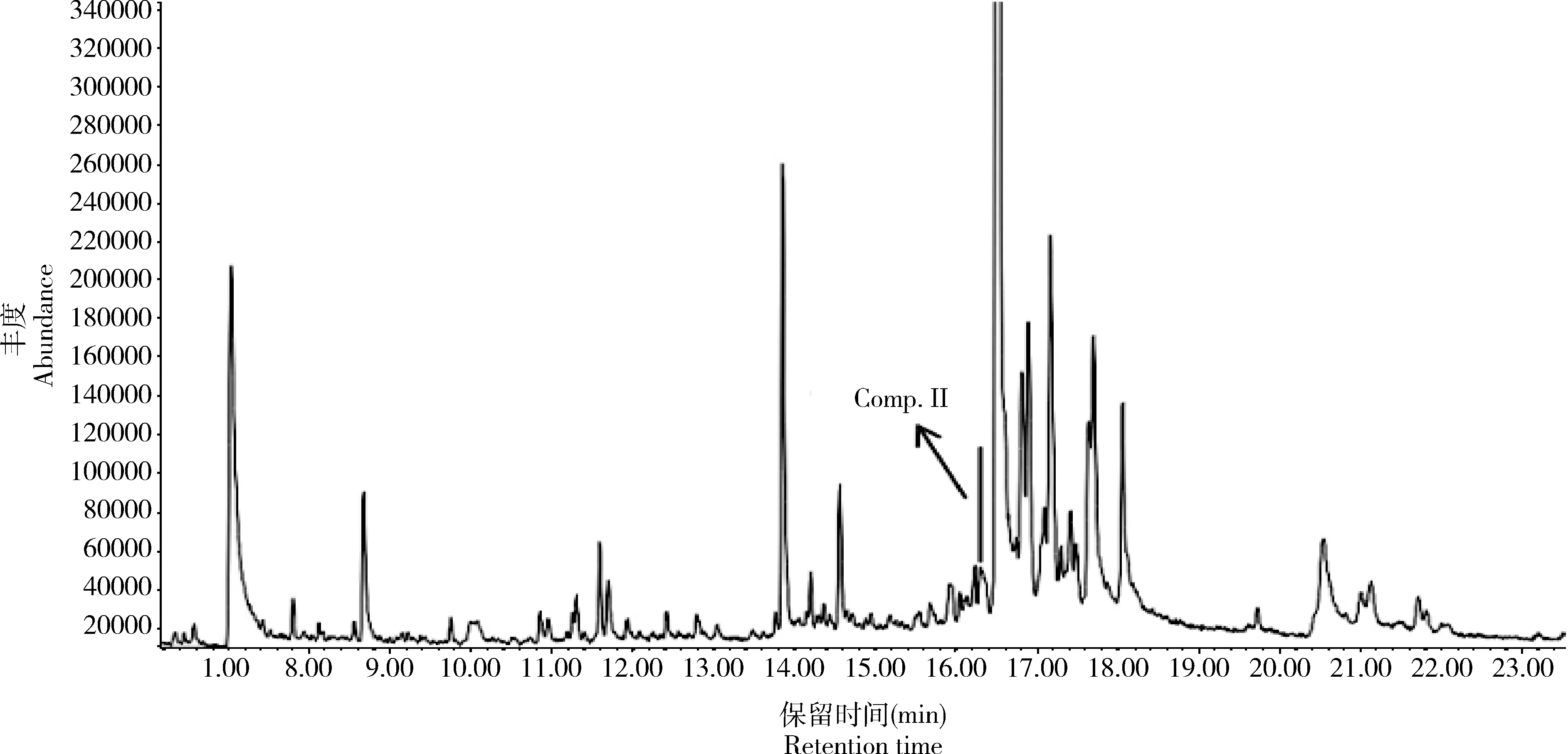
图5 棉大卷叶螟处女雌蛾性信息素腺体浸提液的总离子流图

图6 棉大卷叶螟雌蛾性信息素组分Comp. II的质谱图
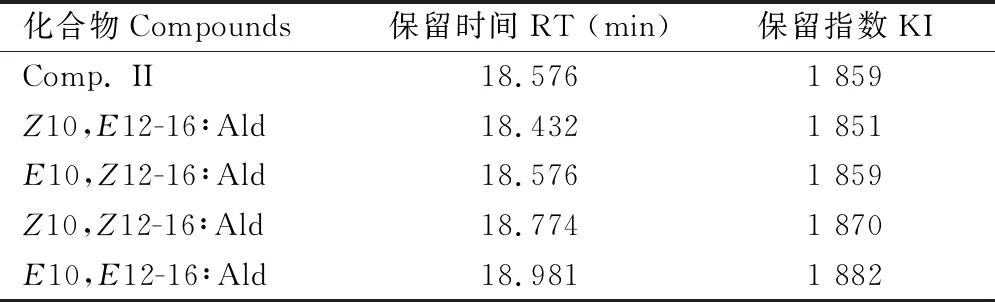
表1 棉大卷叶螟雌蛾性信息素组分Comp. II和10,12-16∶Ald同分异构体在BR-5色谱柱上的保留时间和保留指数
2.4 雄蛾触角对E10,Z12-16∶Ald标准品的触角电位反应
配制浓度为1 ng/μLE10,Z12-16∶Ald标准品溶液10 μL进行EAG分析,以空气和溶剂正己烷作为对照,选用3日龄雄蛾触角。结果显示,与对照相比,10 ngE10,Z12-16∶Ald可以引起棉大卷叶螟雄蛾触角显著的触角电位反应(图7)。
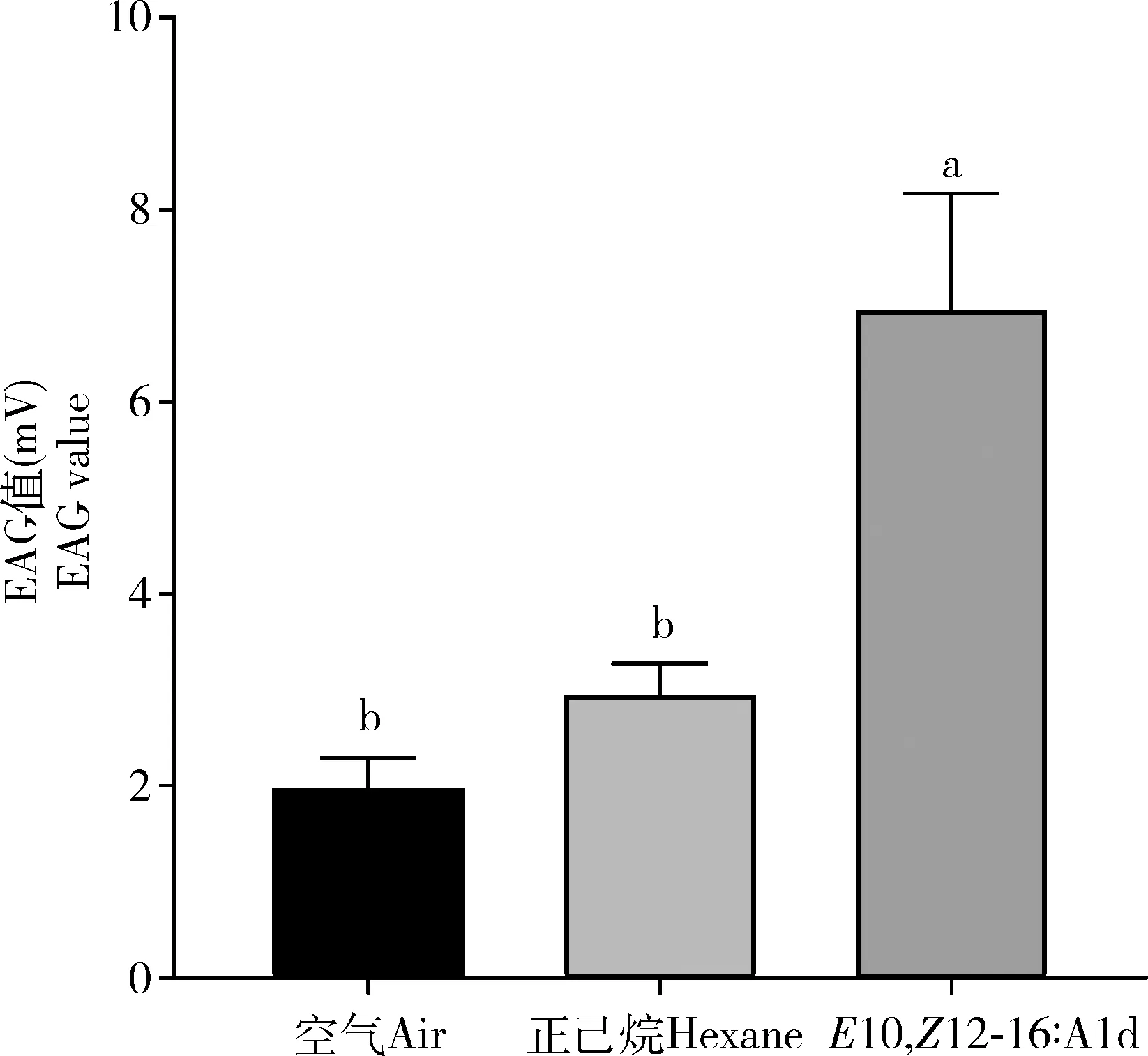
图7 棉大卷叶螟雄蛾触角对E10,Z12-16∶Ald (10 ng)的触角电位反应
进一步对棉大卷叶螟雄蛾进行了不同剂量的E10,Z12-16∶Ald的触角电位实验,结果显示,随着刺激剂量的增加,雄蛾触角对E10,Z12-16∶Ald的触角电位反应不断增强,呈现出明显的剂量效应(图8)。
3 讨论
一般而言,雌蛾的求偶行为伴随着性信息素的释放,求偶高峰期也是性信息素释放的高峰期,因此在求偶高峰期提取性信息素能达到最佳提取效果。受自身性成熟特性和外界环境的影响,不同昆虫求偶的日、时节律均存在较大差异。就昆虫发生求偶行为的日龄看,豹蠹蛾Zeuzeraleuconotum羽化当天即可求偶,在2日龄时就已经处于求偶高峰期(刘金龙等, 2013)。而黄野螟Heortiavitessoides可能由于雌蛾性腺没有发育成熟,在羽化当日没有求偶行为(王忠等, 2018),小地老虎Agrotisipsilon雌蛾在羽化当天的暗期开始求偶,在3日龄暗期7-9 h时求偶最活跃(向玉勇, 2007)。而棉大卷叶螟在羽化当天没有求偶行为,求偶率自1日龄时开始逐渐升高,最大值出现在3日龄时,最大求偶率为70%,随后开始下降。雌蛾求偶起止时间、持续时间的结果也表明,3-4日龄的雌蛾具有更高的求偶趋性,这与很多其他蛾类昆虫如甜菜夜蛾Spodopteraexigua(Dong and Du, 2001)类似。蛾类昆虫通常在暗期进行求偶,在暗期不同时段也呈现节律性变化。棉大卷叶螟雌蛾的求偶高峰期在暗期中期(4-7.5 h),与雌雄蛾间交配行为的高峰时段(暗期2-3 h)(张清泉, 2009)稍有差异,推测可能由于观察棉大卷叶螟交配实验所设置的光周期为13L∶11D,从而导致交配高峰时段的提前。
鳞翅目昆虫性信息素大多属于10~23个碳原子的直链不饱和化合物(Andoetal., 2004),使用GC-MS分析性信息素的化学结构十分高效,但由于在质谱电子轰击(EI)条件下,不饱和脂肪族化合物碳碳双键不容易发生裂解,化合物在形成分子离子后氢原子容易发生沿碳链重排,所以仅凭对质谱图的分析难以确定化合物具体的顺反构型(Ando and Yamakawa, 2011)。一种确定提取物中候选性信息素组分双键构型的方法是通过已知化合物来确定,即通过比较已知化合物和待定组分的保留时间(RT)或保留指数(KI)进行确定具体构型。如Do等(2011)在鉴定梨潜皮细蛾Spulerinaastaurota性信息素的过程中,利用GC比较4种共轭双键位于第9和11位的十六碳烯醇同分异构体(E9,E11-16∶OH,E9,Z11-16∶OH,Z9,E11-16∶OH和Z9,Z11-16∶OH)在不同色谱柱条件下的保留时间与保留指数,从而确定Z9,Z11-14∶OH为梨潜皮细雌蛾性信息素主组分。Yan等(2014, 2018)通过同样的方法确定了甘薯茎螟Palpitanigropunctalis、二点委夜蛾Athetislepigone等害虫性信息素组分的顺反构型。另一种确定提取物中候选性信息素组分双键构型的方法则是利用气相色谱-傅里叶变换红外光谱联用技术(GC-FT/IR)。对于性信息素组分的长链不饱和醛、醇、乙酸酯等结构,除了羧基、羟基有特定的红外吸收谱之外,顺式双键和反式双键的红外特征吸收谱也不相同。自Zarbin等(2012)和Soldi等(2012)将GC-FT/IR相继用于鉴定半翅目昆虫Edessameditabunda和Phthiapicta的性信息素甲基支链的鉴定之后,Shibasaki等(2013)首次将该技术用于鳞翅目昆虫Monemaflavescens的性信息素顺反异构的鉴定中。本研究通过比较Comp. II与10,12-16∶Ald的4种同分异构体在色谱柱上的保留时间和保留指数,从而确定了Comp. II为E,Z构型。
鳞翅目是昆虫纲第二大目,已描述约15万种,目前已鉴定出超过700种蛾类的性信息素,各个种间性信息素的组分和比例都不尽相同,这与种群间的生殖隔离有密不可分的关系。尽管许多昆虫的性信息素中存在相同的主组分,但次要组分在昆虫性信息素的种特异性中也起着重要的作用(Chenetal., 2018)。10,12-16∶Ald作为一种鳞翅目昆虫中常见的性信息素组分,其4种不同的构型及其他次要组分是特定昆虫能够特异性识别自身性信息素的重要原因之一。据报道,华丽野螟Agathodesostentalis(Wakamura and Arakaki, 2004)、绿翅绢野螟Diaphaniaangustalis(Maetal., 2017)、豆野螟Marucatestulalis(Schlägeretal., 2015)等蛾类的性信息素主要组分均为E10,E12-16∶Ald;E10,Z12-16∶Ald也广泛存在于其他鳞翅目昆虫中,比如烟草天蛾Manducasexta(Starrattetal., 1979)、翠纹钻夜蛾Eariasvittella(Corketal., 1988)、三角斑褐蠶蛾Trilochavarians(Daimonetal., 2012)等;Hemarisdiffinis以Z10,E12-16∶Ald作为性信息素主要组分(Reedetal., 1987)。那么对于这些含有相同主组分的昆虫而言,其他次要组分(如10,12-16∶OH,10,12-16∶OAc或其他化合物)则在特异性识别同类和顺利交配中发挥着重要作用。本研究对棉大卷叶螟性信息素提取物的GC-EAD分析中有2种能引起触角电位反应的物质(Comp. I和II),但只对含量较高的主组分Comp. II(E10,Z12-16∶Ald)进行了化学鉴定,Comp. I在腺体中的含量较低,未能在GC-MS分析中获得有效的质谱图。对于Comp. I,我们按照保留时间、同科他种的性信息素文献数据做了一些推测,对化合物E10-16∶Ald和Z12-16∶Ald等做了分析,但它们的保留时间均与Comp. I不完全匹配,E8,Z10-14∶Ald,E8-14∶Ald和Z10-14∶Ald也是可能的候选化合物,但因无法获得该化合物,未做进一步分析,今后如能对该活性物质的结构进行鉴定,则有望用于性诱剂的研发,进而为棉大卷叶螟的性信息素绿色防控技术提供理论基础。
猜你喜欢
上海农业科技(2022年5期)2022-10-19
长江蔬菜(2022年11期)2022-07-29
中国听力语言康复科学杂志(2021年6期)2021-12-21
食品安全导刊(2021年20期)2021-11-28
华人时刊(2020年19期)2021-01-14
上海农业科技(2019年3期)2019-06-25
扬子江(2019年1期)2019-03-08
能源(2018年5期)2018-06-15
数理化学习·教育理论版(2013年9期)2013-12-27
中国火炬(2013年6期)2013-07-24
