
黄河口邻近海域浮游幼虫群落结构季节变化
2022-04-18 10:16左涛李永涛左明程兆龙王俊王安东
海洋学报 2022年4期
左涛 ,李永涛 ,左明,程兆龙 ,王俊 ,王安东
( 1. 中国水产科学研究院黄海水产研究所,山东 青岛 266071;2. 青岛海洋科学与技术试点国家实验室 海洋生态与环境科学功能实验室,山东 青岛 266237;3. 东营市海洋发展研究院,山东 东营 257091;4. 山东黄河三角洲国家级自然保护区管理委员会,山东 东营 257091)
1 引言
浮游幼虫包括永久性浮游幼虫以及诸多底栖无脊椎动物和鱼类发育过程所经历的阶段性浮游幼虫。浮游幼虫是海洋生态系统中一类复杂的生态类群,其类型组成、数量变动等生态特征可指示和预警生物环境区域性或长期变化[1-3]及预测和评估生物资源补充潜力[4]。此外,作为次级消费者—浮游动物主要构成部分及诸多底层海洋生物的补充群体,浮游幼虫还是海洋水层-底层生态系统耦合的关键联接生物类群[2-3,5-7]。在典型河口和海湾等生物繁衍和生长的关键栖息水域,来自底栖和潮间带等不同生境生物的浮游幼虫种类极其丰富,且具有季节性数量旺发等特征,因此,相较于永久性的浮游动物类群而言,浮游幼虫在河口和海湾生态系统碳循环以及不同生境间物质交换中发挥的作用更为突出[2-3,7-8]。
早在20世纪70年代末,国内学者即开始关注浮游幼虫生态研究[9]。之后,在东海、南海的中国近海海湾和河口开展了系列研究,如大亚湾[10]、遮浪湾[11]、闽南-台湾浅滩渔场[12]和南海北部[13]、珠江口[14-15]、北部湾[16]、长江口[17]等。相较而言,在黄河口邻近海域,浮游幼虫多是在浮游动物群落结构相关研究报道[18-22]中被简单提及,对其专门研究明显滞后[23-26]。
黄河口邻近海域位于渤海湾和莱州湾的交界处,生物资源丰富,其中具有典型河口地方性的重要经济鱼类和无脊椎动物记录有50余种[27],且规划了河口生态国家级海洋特别保护区和贝类、鱼类、沙蚕类等多个物种优质生物苗源保护地。这些保护物种在生长发育过程中大多需经历浮游幼虫阶段。海域内浮游幼虫类型多、数量大,是浮游动物组成的主要生态类群和季节性优势种[17-20]。了解和掌握黄河口邻近海域浮游幼虫的种类组成和数量分布对于该水域的生态保护、资源养护和可持续发展增养殖渔业有重要的意义。
本研究于2019-2020年开展了4个季节、6个航次的生态调查,基于调查获得的数据,分析了黄河口邻近海域浮游幼虫的类群组成和丰度的时空分布,探讨了该河口水域浮游幼虫的群落结构与生境因子的关系。研究结果将为评价黄河口邻近海域生态环境健康状况,进而制定生物多样性保护与重要生物资源养护相关政策提供依据。
2 材料与方法
2.1 调查海区和样品采集
调 查 区 域(37.33°~38.17°N,119.00°~119.75°E)位于莱州湾中西部的环黄河入海口附近20 m水深以浅水域。自东营港码头向南至小清河河口外,布设20个生物环境调查站位(图1),使用渔船“鲁昌渔64756”(船长为23 m,宽为6 m,功率为110 kW),先后于2019年4月、6月、8月、10月及2020年1月、4月开展了6个航次的综合调查。
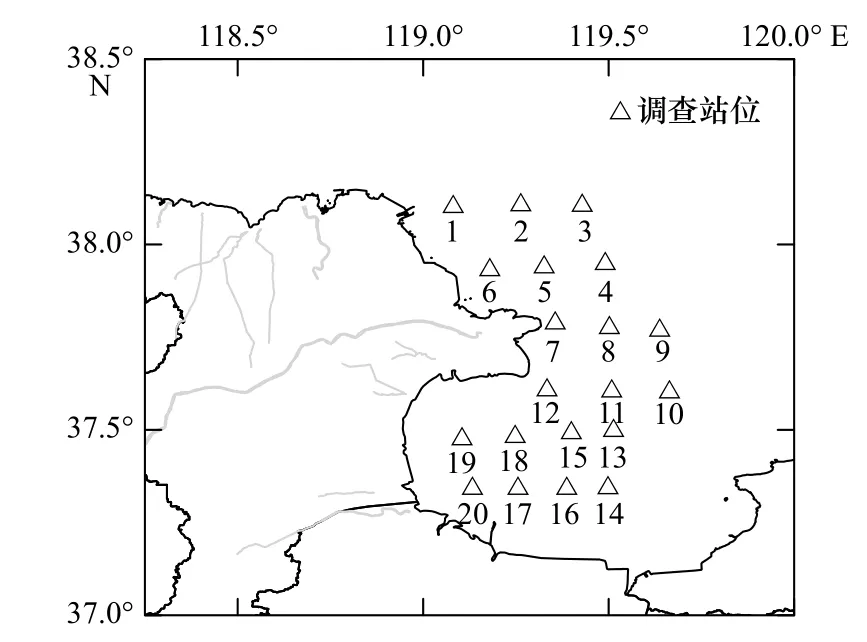
图1 调查区域和站位分布Fig. 1 Study area and stations distribution
按照《海洋调查规范 第6部分:海洋生物调查》(GB/T 12763.6-2007)[28],在各调查站位,使用浅水II型浮游生物网由底层至表层垂直拖网采集浮游幼虫和其他浮游动物;使用浅水III型浮游生物网由底层至表层垂直拖网采集浮游植物;使用采集面积为0.25 m2抓斗式采泥器采集底栖动物;使用多参数水质探测仪(RBR maestro,加拿大)测定水温、盐度、水深、pH、叶绿素浓度等水体环境参数。
2.2 数据分析
在实验室内对生物样品进行分类计数。借鉴郑重等[29]和蔡国雄[11]的研究方法,对样品中的浮游幼虫鉴定到类群。根据每立方米水体中浮游幼虫的个数计算其丰度(A, 单位:ind./m3),计算公式为

式中,n为各站浮游幼虫计数个数;V为采样滤水体积。
浮游幼虫的优势类别根据优势度值确定,优势度(Y)计算公式为

式中,ni为第i类浮游幼虫的丰度;N为所有浮游幼虫的总丰度;fi为第i类的出现频率。若优势度值大于0.02,则该种为优势种。优势种更替率(R)计算公式为
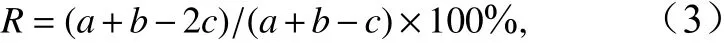
式中,a和b分别为相邻两季节的优势种数;c为相邻两季节共同的优势种数。
浮游幼虫的多样性采用香农-威纳多样性指数(H′)[30]指标来评价。计算公式为
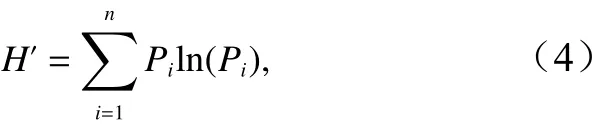
式中,n为总类群数;Pi为第i类浮游幼虫占总该浮游幼虫总数量的比例。
应用PRIMER6.0软件包进行浮游幼虫的群聚结构分析[31]。分析数据包括浮游幼虫丰度、非生物和生物环境因子参数。非生物环境因子数据包括同期表层水温、表层盐度、pH和黄河入海月径流量、水深。黄河入海月均径流量数据为利津水文站观测值(山东黄河网,http://sdb.yrcc.gov.cn/)。生物环境因子数据包括同期各调查站位采集的底栖动物、浮游植物和浮游动物(不含浮游幼虫)的丰度。群聚结构分析步骤依次为,首先将生物和非生物数据分别经对数和正态化转化,采用组平均连接的聚类分析(CLUSTER)和多维定标排序分析(MDS)确定浮游幼虫群落季节(月间)和空间(站位间)群聚结构,通过相似性百分比分析(SIMPER)确定聚类组的代表种,利用相似性分析(One-Way ANOSIM)和多元方差分析(MANOVA)检验浮游幼虫群聚和时空分布差异显著性,采用生物-环境联系BVSTEP逐步多重回归分析模块,明确与浮游幼虫群聚结构匹配的最佳(最大相关系数ρ)环境因子或组合,其匹配相关显著性运用RELATE程序模块检验。
3 结果
3.1 环境因子月变化
如图2所示,海水水温和黄河入海月径流量值以8月最高、1月最低,而盐度、pH以8月最低。浮游植物丰度在夏季8月达峰值,春季4月最低,而浮游动物和底栖动物的丰度分别在春季4月和夏初6月最高,冬季1月最低。上述因子中,水温与Chla浓度、浮游植物丰度以及月径流量呈显著正相关(p<0.05)。
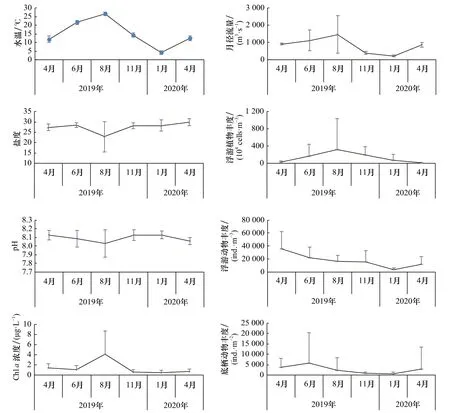
图2 主要环境因子均值的月变化Fig. 2 Monthly variation of mean value of main environment factor
3.2 多样性和优势种组成
共鉴定出浮游幼虫16类,其中环节动物门1类、软体动物门3类、节肢动物门甲壳类7类、帚虫动物门1类、棘皮动物门2类以及脊索动物门2类。除无节幼虫和桡足幼虫外,其他类群均属阶段性浮游幼虫。浮游幼虫多样性指数均值以6月(1.81)和8月(1.44)最高,11月(0.78)和1月(0.72)最低。2019年和2020年两年的4月H′值相近,为1.01和1.12。多样性指数的月间差异显著(MANOVA,F4, 120=14.25,p<0.01),站位间差异不显著(F19, 120=0.95,p=0.52)。
表1显示,所有调查月都出现的浮游幼虫有腹足类幼虫、双壳类幼虫、无节幼虫和长尾类幼虫。其中,无节幼虫和双壳类幼虫是海区所有调查月的优势种。季节性出现的优势种还有腹足类幼虫、多毛类幼虫以及耳状幼虫、桡足幼虫等。优势种更替率以4-6月较高,为0.67,其他相邻调查月间优势种更替率仅为0.2~0.33。
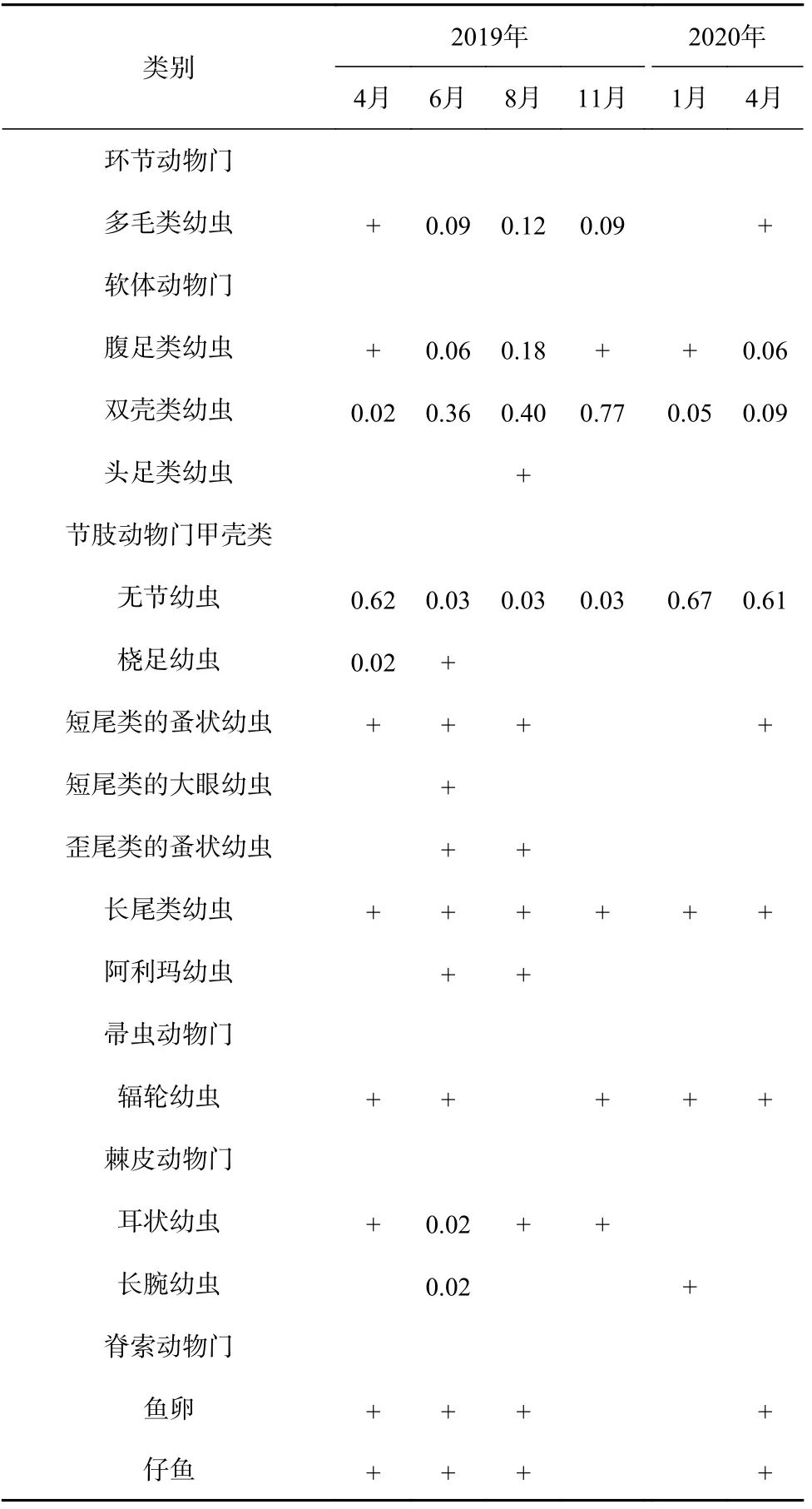
表1 黄河口邻近海域浮游幼虫及其优势种和优势度Table 1 Dominant taxa and their dominance value of planktonic larvae in the Huanghe River Estuary adjacent waters
3.3 丰度时空分布
总浮游幼虫丰度月均值为(1 848±1 067)ind./m3,以11月最高,1月最低。无节幼虫在4月和1月、双壳类幼虫在6-11月为浮游幼虫总丰度的主要贡献者(贡献率大于65%)。主要类群的丰度峰值出现的季节不同,无节幼虫在4月、耳状幼虫在6月、腹足类幼虫在8月、多毛类幼虫和双壳类幼虫在11月的丰度最高(图3)。

图3 总浮游幼虫及主要浮游幼虫类群的丰度均值Fig. 3 Mean abundance of total planktonic larvae and main planktonic larvae taxa
图4显示,总浮游幼虫丰度高值基本与双壳类幼虫和无节幼虫丰度高值的时空分布重合;主要类群丰度的月间变化梯度较站位间更为明显;主要类群的相对高丰度值虽然出现的季节不同,但其多分布于紧邻黄河入海口(6~8号站)和小清河口外、莱州湾中部(14~15号站,19~20号站)站位。
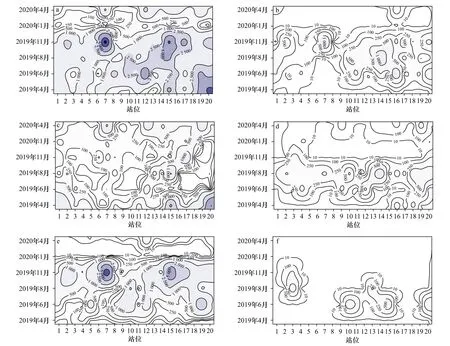
图4 总浮游幼虫丰度(ind./m3)及主要类群丰度(ind./m3)的时空分布Fig. 4 Temporal-spatial abundance (ind./m3) distributions of total planktonic larva and major taxa
月间和站位间的多因子方差分析(MANOVA)结果显示(表2),浮游幼虫主要类群的丰度月间差异显著(p<0.05),站位间的差异不显著(p>0.05)。
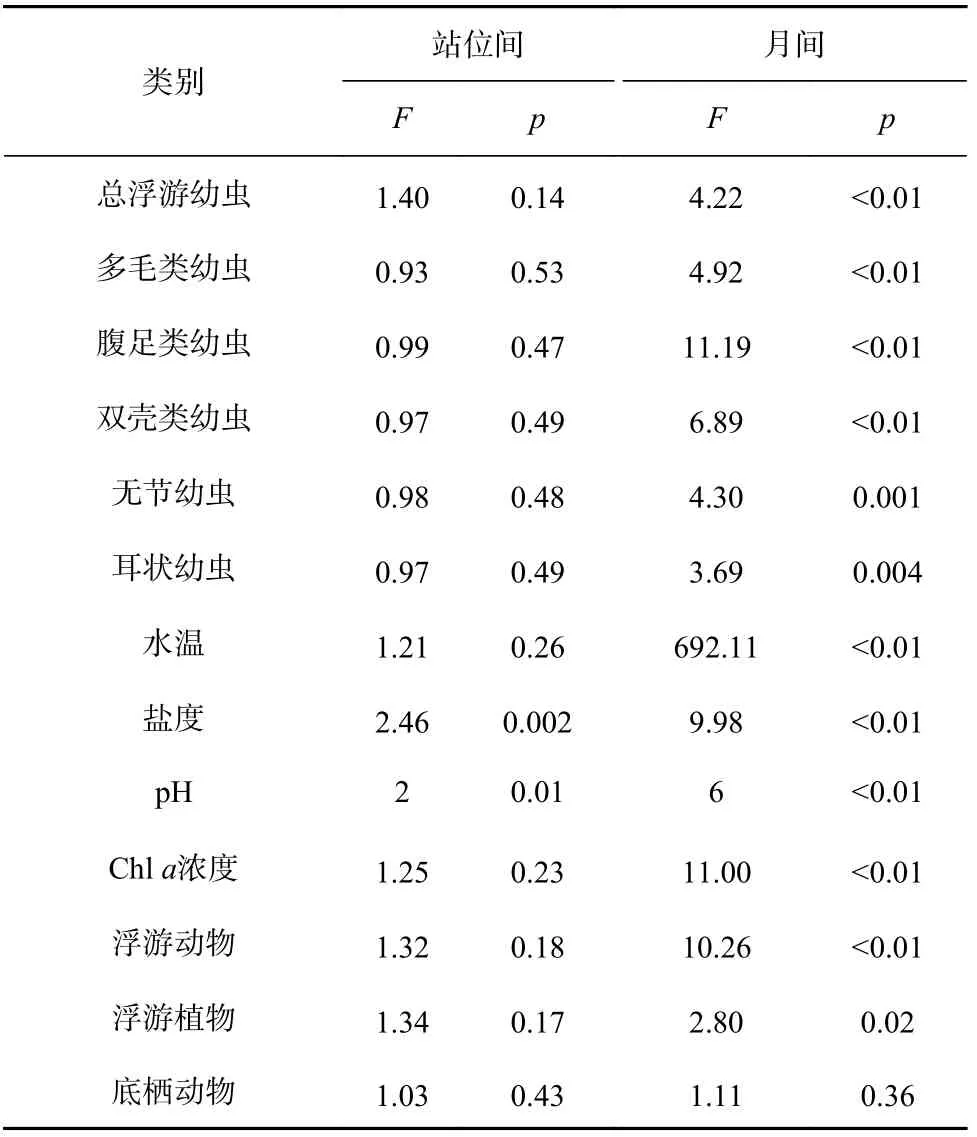
表2 黄河口邻近海域浮游幼虫及环境因子多元方差分析统计信息Table 2 Statistical information from MANOVA analysis of planktonic larvae and environmental factors in the Huanghe River Estuary adjacent waters
3.4 群聚结构及其环境因子关系
聚类分析结果显示,6个调查月和20个调查站在70%和88%相似水平下可分为3个聚类组(图5),两聚类结果的多维度排序分析(MDS)的压力系数分别为0.01和0.15,均小于0.20,且各聚类组间差异达显著水平(p<0.05),该分组达可接受水平。SIMPER分析确定4月(M2)和6-11月(M3)聚类组的代表类群(累积贡献率大于50%)分别是无节幼虫和双壳幼虫。1月聚类组(M1)中浮游幼虫类群少,以无节幼虫丰度较高。3个站位聚类组代表类群都为双壳类幼虫和无节幼虫,且其组成站位地理分布交错。月间和站位间的两因子交叉相似性分析(two-way crossed ANOSIM)显示,浮游幼虫群聚组成的月间差异显著(ρ=0.188,p=0.001),站位间差异不显著(ρ=0.029,p=0.149)。
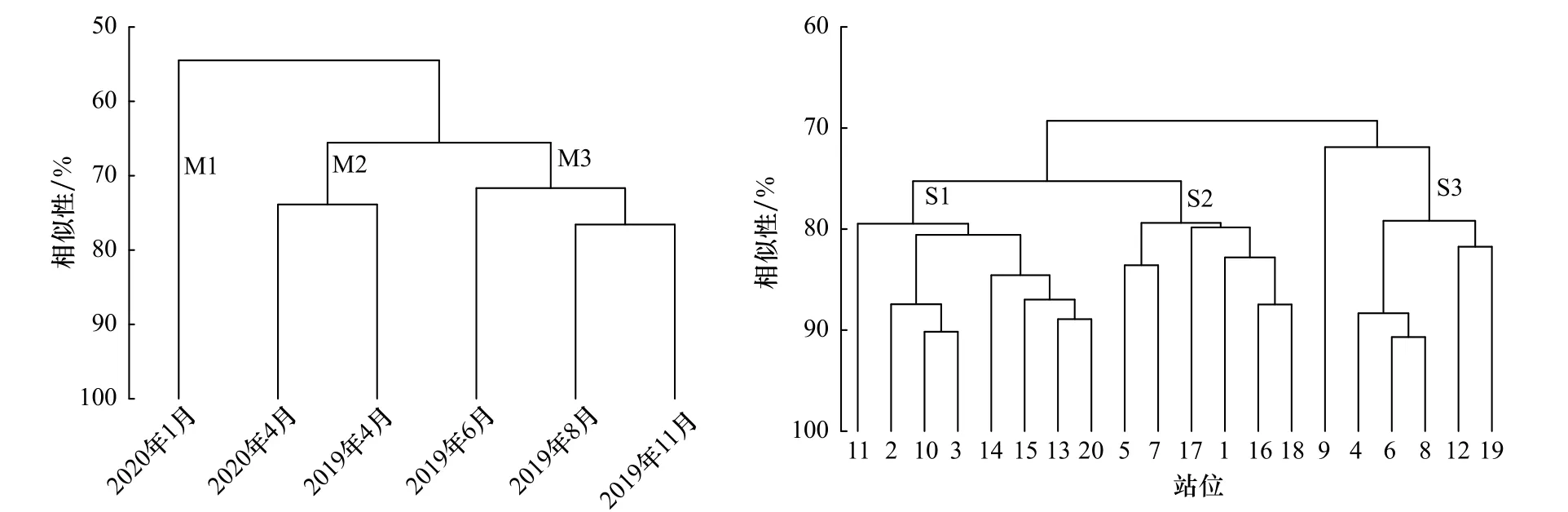
图5 浮游幼虫丰度聚类分析Fig. 5 Cluster analysis of planktonic larva based on the abundance among months and stations
生物-环境关系分析显示,与浮游幼虫群聚结构匹配的最佳环境因子组合为水温和浮游动物丰度,相关性检验达显著水平;对浮游幼虫站位群聚结构差异影响最大的环境因子组合为pH、浮游动物丰度,但其相关性未达显著水平(表3)。
主要浮游幼虫类群中,以无节幼虫的高密区(丰度大于103ind./m3)的水温最低(10~15℃),其平均分布温度为13.8℃;腹足类幼虫的高密区(丰度大于103ind./m3)对应水温最高(>20℃),其平均分布温度为22.3℃。双壳类幼虫和多毛类幼虫的高密区(丰度大于103ind./m3)对应的水温均大于10℃,其平均分布温度接近,分别为21.3℃和20.2℃,但是双壳类幼虫的极高密区(丰度大于5×103ind./m3)的水温为15℃左右(图6)。
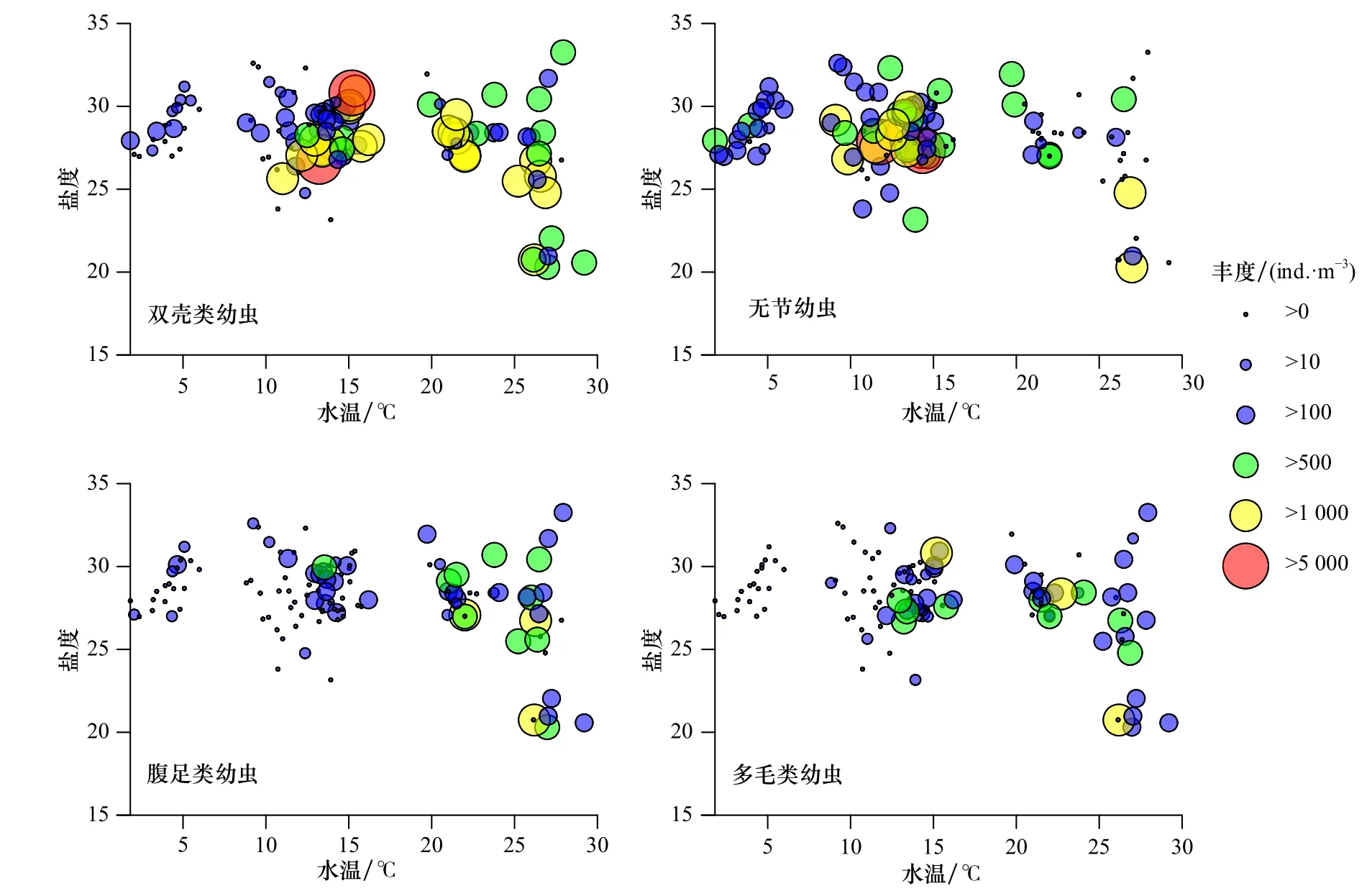
图6 主要浮游幼虫类群的丰度与水温和盐度的相对分布Fig. 6 Relative distributions of the main planktonic larvae taxa abundance with water temperature and salinity
4 讨论
本研究结果表明,黄河口邻近海域浮游幼虫的丰度、多样性以夏、春季较高,秋、冬季较低,群聚结构季节变化明显,这些特征与该海域的浮游动物群落时空变化特征相似[18,20]。研究水域内,无节幼虫和双壳类幼虫数量最多,是各季节的优势类群,多毛类幼虫和腹足类幼虫是夏季的优势类群。这些幼虫亦是2010年9月调查中浮游动物优势种类[21]。研究表明,与闽南-台湾浅滩[12]和北部湾[16]等类似,底栖动物的阶段性浮游幼虫是黄河口近海浮游幼虫类群组成的主要部分,其丰度在浮游幼虫的丰度组成中亦占有相当大的比例。黄河口及其邻近海域浮游幼虫主要类群组成季节性变化特点可能与底栖动物的生殖周期和分布有关[15,29]。底栖动物往往呈季节性的集中产卵,其浮游幼虫周期变化很大(从数小时到数周或月)[32],遇到对它生存有利的底质时,会在很短时间内完成变态实现附着,浮游幼虫数量急剧降低,因此,浮游幼虫类群种类组成表现出明显的周期性和季节更替[4,10]现象。
本研究显示,水温是对浮游幼虫的聚集结构具有显著影响的环境因子。一般认为,水温和饵料生物旺发是触发底栖动物产卵和幼虫丰度峰值出现的重要因素[7,33]。在水温较高、饵料较多的春、夏、秋3季,底栖动物成体繁殖较快,幼虫的种类和数量随之增多。反之,在水温较低,饵料较少的冬季,成体停止繁殖或繁殖很慢,幼虫的种类和数量势必大大减少[32]。温度是控制双壳类生殖期时间的主要因素[34-35],研究区的双壳类和腹足类幼虫平均分布水温较高(>21℃)。此外,河口陆地径流量、盐度、潮汐、海洋等水文分布也是影响浮游幼虫分布的重要因素[33,36-39],如表3所示,虽然月径流量和浮游植物未被列入最佳影响因子组合中,但它们与浮游幼虫的月群集结构间具有较高的相关系数,而春季无节幼体丰度激增,可能与初级生产增加相关[7]。
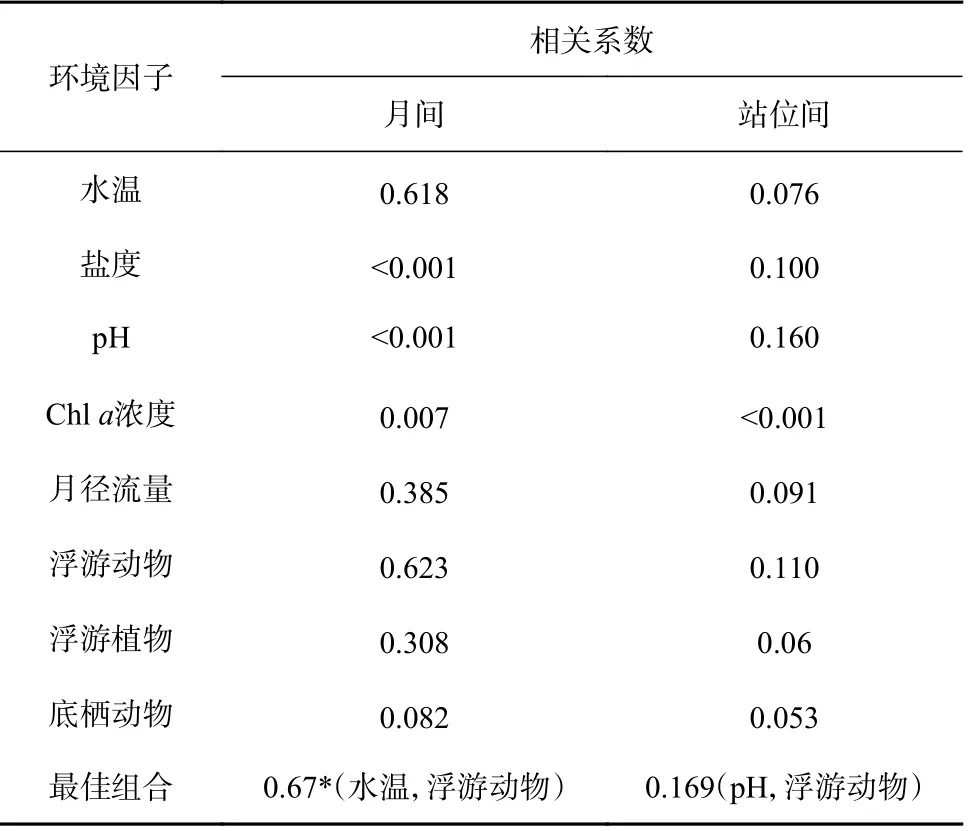
表3 环境因子及其最佳匹配组合与浮游幼虫群聚结构的Weighted Spearman 等级相关系数Table 3 Weighted Spearman rank correlation coefficient for environmental factors and their best combinations with the planktonic larvae assemblage structure
本文的研究结果显示,包括双壳类幼虫和腹足类幼虫等在内的贝类幼虫是海区内浮游幼虫优势类群,特别是双壳类幼体在黄河口邻近海域分布广泛,全年都有出现,且是各季节的优势类群。在2012-2013年的周年调查也发现,双壳类幼虫丰度值高,是春、秋季黄河口海域浮游动物的优势种[20]。图4显示,在6月、8月和11月,调查区内出现多个双壳类幼虫丰度大于1 000 ind./m3的高密区,特别是在11月,有些站位的丰度大于104ind./m3。在2010年9月的调查中也发现,双壳类的浮游幼虫是除夜光虫(Noctiluca scintillans)外,数量密度最高的浮游动物类群,其丰度达3 485 ind./m3[21]。海区内贝类幼虫分布广、数量多与黄河口邻近海域有丰富的贝类资源密切相关。黄河口有绵延广阔的泥沙质滩涂,为众多贝类提供了优越的栖息繁殖场所,分布于黄河口滩涂的贝类资源生物达40余种[27]。本研究结果表明,黄河口邻近海域双壳类和腹足类幼虫丰度季节变化明显,以夏、秋季居多,在空间上则呈明显的斑块分布,高丰度集中分布于紧邻黄河口入海口、小清河口及莱州湾等区域(图4)。上述双壳类和腹足类幼虫数量的时空分布特点可能与黄河口不同贝类成体的生活空间具有区域界限、生殖期具有季节差异有关。海区内多个贝类优势种中,光滑篮蛤(Potamocorbula laevis)、脉红螺(Rapana venosa)、西施贻贝(Coelomactra antiquata)和菲律宾蛤仔(Ruditapes philippinarum)等的栖息密度亦以夏、秋季居多;四角蛤蜊(Mactra veneriformis)、文蛤(Meretrix meretrix)等以夏、冬季居多,彩虹明樱蛤(Moerella iridescens)和红明樱蛤(Moerella rutila)以春季居多[27]。
浮游幼虫涉及的生物门类广,同种浮游幼虫随发育期和环境变化而形态各异,同属种类的幼虫高度相似,因此,从形态学角度进行浮游幼虫物种水平上的分类鉴定较为困难。本研究仅对样品中各浮游幼虫鉴定到类别,结果并不能完全反映黄河口邻近海域浮游幼虫的多样性和物种丰富度水平。近年来,已证实基于已有的生物分子序列信息,利用分子生物学高通量测序、DNA条形码等技术可从类属、物种水平区分和鉴定浮游幼虫种类[39-41]。当前,黄河口邻近海域的浮游生物和底栖生物的分子生物学本底信息比较缺乏。因此,进一步研究工作中,应结合形态学和分子测序技术,发展和构建相关的种类比对信息数据库,提高浮游幼虫的种类鉴定水平,为细致刻画黄河口邻近海域浮游幼虫群落时空分布格局提供技术支撑,提高对黄河口生态系统中浮游幼虫生态功能作用的认识。
猜你喜欢
当代水产(2022年6期)2022-06-29
音乐天地(音乐创作版)(2022年3期)2022-06-14
金桥(2021年11期)2021-11-20
金桥(2021年11期)2021-11-20
农业科技与信息(2020年14期)2020-12-18
青年歌声(2020年10期)2020-12-03
流行色(2019年10期)2019-12-06
绿色科技(2019年14期)2019-11-19
装饰装修天地(2019年9期)2019-10-21
江苏农业科学(2019年5期)2019-09-02
