
黄河口及邻近海域鱼卵、仔稚鱼群落结构及多样性的年间变化
2022-04-18 10:15:58杨艳艳朱明明冯银银李焕军陈玮王秀霞陈建强苏海霞
海洋学报 2022年4期
杨艳艳,朱明明,冯银银,李焕军*,陈玮,王秀霞,陈建强,苏海霞
( 1. 山东省海洋资源与环境研究院 山东省海洋生态修复重点实验室,山东 烟台264006)
1 引言
黄河口及邻近海域位于莱州湾和渤海湾的交界处,是我国重要的河口之一[1]。黄河入海径流所携带的丰富营养盐和有机质等维持了黄河口及邻近海域较高水平的初级生产力和饵料生物多样性,使其成为多种鱼类的产卵场、索饵场和育肥场[2-3],同时也是多种溯河和降海洄游鱼类的通道,对鱼类种群的繁衍生息具有十分重要的意义[4]。
鱼卵、仔稚鱼在黄河口及邻近海域生态系统中扮演着重要角色[5],是黄河口及邻近海域海洋鱼类资源利用和补充的基础,其种群成活率的高低、剩存量的多寡对黄河口及邻近海域鱼类补充群体资源量的大小起决定性作用[5-7]。在6月(夏季)鱼类繁殖的重要时段内,开展鱼卵、仔稚鱼群落结构年间变动的研究,对于更加全面地了解河口生态系统的结构和功能、及时掌握鱼类种群的生存与延续状况以及河口鱼类资源的保护等都具有重要的理论和实践意义[8]。
国内近年来针对黄河口及邻近海域鱼卵、仔稚鱼的研究较少,虽然文献显示从20世纪80年代起,我国学者就开始对黄河口及邻近海域鱼卵、仔稚鱼的生物生态学展开调查,但历次调查中真正分布于黄河口及邻近海域的站位不多,所以可供参考的历史资料非常有限[4,9-12]。近十几年来,卞晓东等[13]、刘霜等[14]和秦雪等[15]分别针对黄河口及邻近海域鱼卵、仔稚鱼的种类组成和数量分布开展了相关研究,但有关黄河口及邻近海域鱼卵、仔稚鱼群落结构及多样性年间变动的研究较少。
本文根据2014-2018年开展的黄河口及邻近海域鱼卵、仔稚鱼调查结果及同步观测的环境资料,分析了该海域鱼卵、仔稚鱼的种类组成、数量分布、生物多样性等,探讨了该海域鱼卵、仔稚鱼群落结构的年间变动情况,以期为研究黄河口生态系统渔业资源的再生和补充积累基础资料,并为黄河口海域渔业资源的开发和保护提供依据。
2 材料方法
2.1 调查时间和站位
分别于2014年、2015年、2017年和2018年6月在黄河口及邻近海域进行4个航次的表层水平拖网取样调查。依据与黄河入海口的距离布设4个断面(图1),I断面(B1站、C1站、D1站)距入海口5 km,II断面(A2站、B2站、C2站、D2站、E2站)距入海口10 km,III断面(A3站、B3站、C3站、D3站、E3站)距入海口20 km,IV断面(A4站、B4站、C4站、D4站、E4站)距入海口40 km,共设置18个站位。
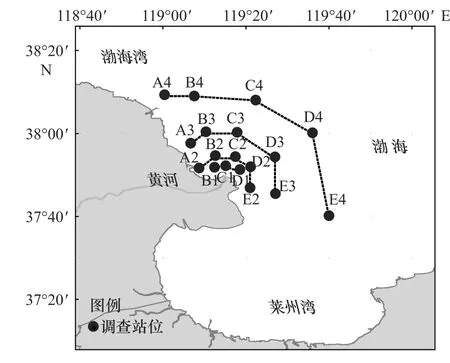
图1 黄河口及邻近海域调查站位Fig. 1 Sample stations in the Huanghe River Estuary and its adjacent waters
2.2 样品采集
鱼卵、仔稚鱼样品采集及分析方法严格按照《海洋调查规范 第6部分:海洋生态调查》(GB/T 12763.6-2007)[16]进行。采用大型浮游生物网(口径为80 cm、长为280 cm、网目尺寸为0.505 mm)逐站进行表层水平拖网采样,每站拖10 min,拖速为2 n mile/h,样品用5%福尔马林海水溶液固定保存带回实验室。根据鱼卵、仔稚鱼的形态特征(包括鱼卵和仔稚鱼的外部形态以及不同发育时期的个体形态和组织、器官、系统发育特征等)利用解剖镜进行种类鉴定,并计数[17-19]。定量资料取自水平网采数据,以每网的实际数量(粒、尾)为指标计算鱼卵、仔稚鱼的密度(粒或尾/网)[13]。
温度、盐度等水文环境因子使用XR-420型CTD现场测定。悬浮物、无机氮和活性磷酸等环境因子的样品采集和分析参照《海洋监测规范 第1部分:总则》(GB/T 17378.1-2007)[20]执行。
2.3 数据处理
2.3.1 优势种
利用相对重要性指数(Index of Relative Importance,IRI)对鱼卵和仔稚鱼的优势种进行计算[13],公式为

式中,N%为某一种类个体数量占采获总量的百分比;F%为某一种类出现的站位数占总调查站位数的百分比,IRI不小于1 000的种类为优势种。
2.3.2 生物多样性
3种生物指数如下:
物种丰富度指数(D)计算公式[21]为

香农-威纳多样性指数(H′)计算公式为

均匀度指数(J′)计算公式为

式中,S为种类数;N为总个体数;Pi为第i种个体数占总个体数的比例。
2.3.3 群落结构分析
群落结构聚类分析使用R软件(version 4.1.0)的vegan、cluster等程序包进行。采样数据的分析采用平均聚类分析方法,首先,按照每一年采获鱼卵、仔稚鱼的种类,将采集数量分别整理排列,利用对数进行标准化转换,计算年份之间的Bray-Curtis相异系数,然后进行非加权组平均法(Unweighted Pair-group Method with Arith-metic Aeans,UPGMA)聚类。
采用相似性百分比分析(Similarity Percentage Analysis,SIMPER)方法分析造成不同组间相异的分歧单元和平均相异性贡献率[22-23]。
2.3.4 环境因子相关关系分析
将调查期间每一年每一站位的鱼卵、仔稚鱼生物多样性各指数与同步监测的水温(Sea Surface Temperate, SST)、盐度(Sea Surface Salinity, SSS)、水深(Depth)进行皮尔逊相关性分析,并运用R软件绘制各因子之间的皮尔逊相关性矩阵。
2.4 产卵亲体的适温类型、栖所类型和生态类型
根据鱼类适温类型[24],将产卵亲体划分为暖温种(Warm Temperate Species,WT)、暖水种(Warm Water Species,WW)和冷温种(Cold Temperate Species,CT)。
根据栖所类型[25]将黄河口及邻近海域产卵亲体划分为大陆架浅水底层鱼类(Continental Shelf Demersal Fish,CD)、大陆架岩礁性鱼类(Continental Shelf Reef-associated Fish,CRA)、大陆架浅水中上层鱼类(Continental Shelf Pelagic-neritic Fish,CPN)、大陆架浅水中底层鱼类(Continental Shelf Benthopelagic Fish, CBD)、大陆架大洋洄游性中上层鱼类(Oceanic Pelagic Fish,OEP)和大洋深水底层鱼类(Oceanic Bathydemersal Fish,OMP)。
根据鱼类的生态类型,参考文献[26-27],将产卵亲体划分为半咸水型鱼类、沿岸型鱼类、近海型鱼类。
图件绘制使用ArcGIS 10.6、Excel 2016和R软件(version 4.1.0)完成。
3 结果
3.1 种类组成及年际变化
4个航次共采集鱼卵3 829粒,仔稚鱼234尾。经分析鉴定共18个种类,其中鉴定到种的15种,隶属于7目14科15属,另有1种虾虎鱼科仔稚鱼鉴定到科,1种鱼卵和1种仔稚鱼未鉴定出种类(表1)。鲈形目(包括虾虎鱼)鱼类出现7种,占38.89%;鲽形目鱼类出现3种,占16.67%;鲱形目鱼类出现2种,占11.11%;颌针鱼目、鲻形目、刺鱼目和鲉形目鱼类各出现1种,分别占5.56%。在已经鉴定到种的鱼卵、仔稚鱼中,亲体以暖温种为主,共8种,占53.33%;其次为暖水种,5种,占33.33%;冷温性种类最少,2种,占13.33%。从亲体的栖所类型来看,调查海域内主要以浅水中上层、浅水底层、浅水中底层以及岩礁性鱼类4种生态类型鱼类为主要组成。

表1 黄河口及邻近海域鱼卵、仔稚鱼种类Table 1 The species of fish eggs and larvae in the Huanghe River Estuary and its adjacent waters
4年间调查海域内鱼卵、仔稚鱼的种类数和数量变化较大,2014年和2015年采集的鱼卵、仔稚鱼总种类数均为10种,高于2017年和2018年。其中,2015年采集鱼卵、仔稚鱼的总数量为1 431粒或尾,为4年最高,2014年采集的总数量最低,仅为2015年的42.07%。此外,调查期间鱼卵的种类数和数量变化较大;仔稚鱼种类数整体变化不大,数量方面以2018年采集的仔稚鱼最多,其次为2017年、2014年和2015年(表2)。

表2 黄河口及邻近海域鱼卵、仔稚鱼种类和数量的年间变化Table 2 Annual variation of species and quantity of fish eggs and larvae in the Huanghe River Estuary and its adjacent waters
3.2 数量分布空间和时间变化
2014年共采集鱼卵580粒,各站位数量在0~217粒/站之间波动,平均密度为32.22粒/站,其中共有4个站位密度大于50粒/网,以位于黄河入海口北部的C4站鱼卵数量最多;2015年采集鱼卵1 414粒,各站位数量在1~906粒/站之间波动,平均密度为78.56粒/网,其中共有7个站位密度大于50粒/网,以位于黄河入海口西部的A3站数量最多;2017年采集鱼卵676粒,各站位数量在4~226粒/网之间波动,平均密度为37.56粒/网,其中共有3个站位密度大于50粒/网,以位于黄河入海口南部的E3站数量最多;2018年采集鱼卵1 159粒,各站位数量在0~682粒/网之间波动,平均密度为37.89粒/网,其中共有5个站位密度大于50粒/网,以位于黄河入海口东部的D3站数量最多(图2)。

图2 黄河口及邻近海域鱼卵空间分布Fig. 2 Space distribution of fish eggs in the Huanghe River Estuary and its adjacent waters
2014年采集仔稚鱼22尾,各站位数量在0~8尾之间波动,平均密度为1.22尾/站,最高值出现在黄河入海口西部的B1站位。其中虾虎鱼科未定种Gobiidae sp.数量最多,占本航次仔稚鱼数量的68.19%;2015年采集仔稚鱼17尾,各站位数量在0~12尾/站之间波动,平均密度为0.94尾/站,最高值出现在黄河入海口西部的A2站位。其中鳀(Engraulis japonicus)仔稚鱼数量最多,占本航次仔稚鱼数量的64.7%;2017年采集仔稚鱼72尾,各站位数量在0~30尾之间波动,平均密度为4尾/站,最高值出现在黄河入海口东部的D1站位。其中斑鰶(Konosirus punctatus)仔稚鱼数量最多,占本航次仔稚鱼数量的98.6%;2018年采集仔稚鱼123尾,各站位数量在0~120尾/站之间波动,最高值也出现在黄河入海口东部的D1站位。其中鳀仔稚鱼数量最多,占本航次仔稚鱼数量的97.6%(图3)。

图3 黄河口及邻近海域仔稚鱼空间分布Fig. 3 Space distribution of larvae in the Huanghe River Estuary and its adjacent waters
总体来看,调查海域鱼卵、仔稚鱼的分布呈现出明显的区域特征,鱼卵的分布范围相对来说更广,以距离黄河入海口较远的III断面和IV断面尤为密集,仔稚鱼的分布则主要集中在距离黄河口入海口较近的I断面和II断面上。
3.3 优势种
2014年鱼卵的优势种为短吻红舌鳎(Cynoglossus joyeri)和鳀,仔稚鱼的优势种为虾虎鱼科未定种;2015年鱼卵的优势种为鳀,仔稚鱼没有出现优势种,鳀和斑鰶构成仔稚鱼的重要种;2017年和2018年鱼卵的优势种也均为短吻红舌鳎,2017年仔稚鱼的优势种为斑鰶;2018年仔稚鱼没有出现优势种,鳀为仔稚鱼的重要种(表3)。

表3 黄河口及邻近海域鱼卵、仔稚鱼优势种组成和相对重要性指数(IRI)Table 3 Important species composition of fish eggs and larvae and the index of relative importance(IRI) in the Huanghe River Estuary and its adjacent waters
3.4 群落多样性及其与环境因子相关性
2014年和2015年的物种丰富度指数(D)、多样性指数(H′)和均匀度指数(J′)基本持平,从2017年开始D、H′和J′呈逐渐下降的趋势,2018年的D、H′和J′均降至4年最低,整体来看,4年间黄河口及邻近海域鱼卵、仔稚鱼的群落多样性呈现先持平后下降的趋势(图4)。

图4 2014-2018年黄河口及邻近海域鱼卵、仔稚鱼生物指数Fig. 4 Species indexes of fish eggs and larvae in the Huanghe River Estuary and its adjacent waters from 2014 to 2018
从3种指数与环境因子相关关系分析来看,多样性指数(H′)和均匀度指数(J′)与表层盐度呈显著正相关(p<0.05),与温度和水深不存在相关性(图5)。

图5 生物指数与环境因子的皮尔逊相关性矩阵Fig. 5 Pearson correlation matrix between diversity of species indexes and environmental factors
3.5 群落结构
利用Bray-Curtis相似性系数公式计算不同年间种类组成的相似性系数,然后运用R语言聚类树对其进行聚类分析,根据融合水平值定义划分水平,选择具有最大跳跃的分组水平,将4个年份的鱼卵、仔稚鱼划分为3个组,进一步聚类分析结果显示,3个分组分别为2014年和2018年的组1、2015年的组2和2017年组3(图6,图7),其中,组1和组2,组1和组3以及组2和组3之间的主要分歧种均为鳀,鳀对其年间平均相异性贡献率分别为56.5%、53.4%和60.9%。

图6 聚类树的融合水平值Fig. 6 Fusion level values of cluster tree

图7 黄河口及邻近海域鱼卵、仔稚鱼群落结构年间变化聚类分析Fig. 7 Cluster analysis of annual changes in the community structure of fish eggs and larvae in the Huanghe River Estuary and its adjacent waters
4 讨论
4.1 种类组成和数量分布的变化特征
本研究中4个航次调查共采获鱼卵、仔稚鱼18种,占该海区全年记载总种类数的47%[9,24],主要以鳀、斑鰶、鲬(Platycephalus indicus)等近海型种类为主。4年间采获的鱼卵、仔稚鱼中鳀的数量超过总量的50%,短吻红舌鳎4年间有3年均作为鱼卵的优势种出现,说明鳀以及短吻红舌鳎目前是黄河口及邻近海域最具代表性的物种,其鱼卵、仔稚鱼正在以其数量和对群落的贡献控制着黄河口及邻近海域鱼类早期补充群体的群落演替格局[28]。
从种类组成的年间变化来看,除鳀、斑鰶、多鳞鱚(Sillago sihama)和短吻红舌鳎外,其余种类均演替出现,例如:小带鱼和虾虎鱼科鱼类在2014年出现之后,2015年和2017年消失,2018年又出现;还有些种类仅在个别年份出现,如皮氏叫姑鱼(Johnius belangerii)仅在2014年出现,日本下鱵(Hyporhamphus sajori)仅在2017年出现,鮻(Liza haematocheilus)仅在2015年出现。鱼卵、仔稚鱼种类组成的这种年间演替一方面与产卵亲体正常的种类变动有关,另一方面考虑到调查水域特殊的河口生态环境,其生境状况较一般海域更加复杂多变,如本研究调查期间,月均径流量差接近30×108m3[29],同步监测的平均表层水温差超过2℃,表层盐度差接近6。此外,氨氮、磷酸盐、总氮和总磷等营养盐浓度也均处于波动状态,所以笔者分析这种较为强烈的生境条件波动可能会对鱼类物种的年间变化有一定的影响[28,30],但是受本研究时间跨度较小所限,无法验证这种年间演替是否与生境变化有关,还需今后在河口水域长时间跨度研究中进一步探讨。
从数量分布来看,4年间鱼卵的数量高值区多集中在调查区域内距黄河入海口相对较远的III、IV断面上,分析原因,一方面与鱼类的产卵习性有关,产卵时期海域的盐度、水流流速和温度等因素均会对产卵亲体的分布造成一定的影响。III、IV断面由于距入海口相对较远,受入海冲淡水的影响较小,盐度适宜,没有低盐区。此外,因为距入海口较远,受入海径流扰动较小,所以水流也相对较缓,更适合鱼类栖息产卵。另一方面由于鱼卵不具备活动能力,在未孵化前要经历被水流挟带被动扩散的过程,所以其分布与海流等海洋要素密切相关[31]。受到黄河口以南的顺时针环流系统、仙河镇外海的顺时针环流系统以及黄河口以北的逆时针环流系统3个环流系统的影响,鱼卵的密集中心多在余流尾部形成[32-33]。仔稚鱼的数量高值区主要集中在靠近黄河入海口较近的I、II断面,其原因一方面与仔稚鱼的游泳能力有关。仔稚鱼已经具备一定的活动能力,可以随着索饵水域的水文条件变化等表现出趋近岸移动等行为[28],本研究中靠近入海口的近岸区域监测的悬浮物浓度相对较高(I断面和II断面的平均悬浮物浓度为27.12 mg/L,III断面和IV断面的平均悬浮物浓度为22.20 mg/L),高浊度及其浅水区复杂的水文地理条件使栖息在该区内的肉食性鱼类较少,仔稚鱼的捕食者较少,增加了仔稚鱼的生存机会[34]。另一方面与黄河口及邻近海域的环境条件和物质基础有关。夏季是黄河的丰水期,进入6月以后径流量明显增加,黄河冲淡水为近岸浅水区带来大量的营养物质,孕育了丰富的饵料资源,为仔稚鱼的生长和发育提供充足的物质基础。此外,由径流入海带来的扰动也随之增加,而仔稚鱼阶段虽然已经具备游泳能力,但是总体来说活动能力不强,视觉灵敏度也较低,由径流入海造成的扰动条件大大增加了仔稚鱼与食物相遇的概率,使其更易获得食物[35]。
4.2 群落多样性的年间变化
本研究结果显示,2017年和2018年的生物多样性处于较低水平,尤其是2018年,各指数仅约为2014年和2015年的1/3~1/2,为4年最低。这说明2018年黄河口及邻近海域的鱼卵、仔稚鱼种群均衡性较低、种间分布不均匀。分析原因,一方面与鱼卵、仔稚鱼种类的减少有关(2017年和2018年采获鱼卵、仔稚鱼种类分别为2014年和2015年的50%及70%),导致了群落的丰富度指数D和多样性指数H′下降[36-37];另一方面与主要优势种类鳀丰度的下降以及出现频率变小也有一定的关系,影响了群落分布的均衡性。除此之外,有文献表明,群落多样性的降低还与非生物环境条件的变化和扰动的增加密切相关[38]。对本研究调查期间的径流量和输沙量进行分析后发现,2018年6月黄河利津站监测的入海径流量为38.62×108m3[29],大于其他3个年份6月径流量之和;输沙量为1.35×107t[29],为其他3个年份的6.55~26.3倍。径流量和输沙量如此大幅的增加,必然会使黄河口及邻近海域的盐度、温度等环境条件发生改变。同时由径流和输沙入海所带来的扰动也必然会大幅增加,所以笔者认为,生物环境条件的变化叠加非生物环境条件的改变和过度的干扰等因素综合作用使2018年的群落多样性降至4年最低[39]。
4.3 群落结构变动评价
黄河口及邻近海域内接黄河,外连渤海[13],独特的地理位置使该区域具有更为复杂的理化特性,形成了较为特殊的河口生态环境和生物群落[40]。从本研究年间聚类结果分析,除2014年和2018年外,其余年份间共同出现种类均较少,年间种类相似程度不高,说明2014-2018年间黄河口及邻近海域鱼卵、仔稚鱼的种类组成结构变化较大。此外,从栖息类型来看,调查中采获中上层和底层鱼类的鱼卵、仔稚鱼种类最多,但是底层鱼类鱼卵、仔稚鱼的采获数量所占比例却很低,仅占采获总量的16%,并且90%以上均为短吻红舌鳎。分析原因,一方面是部分底层鱼类对河口区域复杂多变的环境条件耐受性较差[41],影响了其在黄河口及邻近海域的分布;另一方面,近30年来,黄河径流量的锐减甚至断流改变了河口水域咸淡水的生态格局[41-43]。同时,海洋环境污染及过度捕捞等因素给黄河口及邻近海域带来巨大的压力,其生态环境受到重度干扰,黄河口海域渔业资源的营养级不断从高级向低级发展,渔业资源结构也发生了较大的变化[43-44]。一些重要种类如:带鱼(Trichiurus japonicas)、小黄鱼(Larimichthys polyactis)、鲆鲽类等传统底层经济鱼类资源严重衰退[10],鱼类群落组成进一步向小型中上层的低质种类上转化[45],进而导致该水域鱼类早期补充群体的群落结构也发生相应的改变[46-47]。
猜你喜欢
阳光(2024年1期)2024-01-29 10:44:02
水产科学(2022年1期)2022-01-26 08:43:00
渔业科学进展(2021年6期)2022-01-04 06:26:42
特区文学·诗(2021年6期)2021-12-22 14:15:41
食品工业(2020年6期)2020-07-18 04:07:20
创新作文(3-4年级)(2019年8期)2019-12-17 07:59:21
装饰装修天地(2019年5期)2019-10-21 01:37:09
创新作文(小学版)(2019年23期)2019-01-11 10:37:45
海洋渔业(2016年6期)2016-04-16 03:00:32
应用海洋学学报(2015年3期)2015-11-22 07:39:22
