
基于TRIM家族讨论泛素连接酶TRIM46研究进展
2022-03-25 07:59盛瀚萱张连峰
中国比较医学杂志 2022年2期
盛瀚萱,张连峰,张 丽∗
(1.中国医学科学院医学实验动物研究所,北京协和医学院比较医学中心,国家卫生健康委员会人类疾病比较医学重点实验室,北京 100021;2.中国医学科学院医学实验动物研究所,北京协和医学院比较医学中心,北京市人类重大疾病实验动物模型工程技术研究中心,北京 100021;3.中国医学科学院神经科学中心,北京 100730)
TRIM46(tripartite motif-containing46,TRIM46)是一个新基因,功能尚不明确,国内外相关文献很少。但就近期的文献报道来看,TRIM46十分重要,不仅在早期脑发育中参与神经元极化过程[1],还与肿瘤的增殖、迁移相关[2],并在先天免疫系统中扮演了重要角色[3]。TRIM46属于TRIM家族,而TRIM蛋白质家族成员在细胞周期、细胞凋亡、分化、代谢以及病毒的免疫应答等过程中均有重要作用。作者通过将TRIM46与其他家族成员乃至亚家族成员之间的比较,总结了TRIM46的最新研究进展,并对TRIM46蛋白未来的研究方向和手段进行了初步讨论。
1 TRIM家族及C-I亚家族
人类三重基序(tripartite motif,TRIM)蛋白质家族成员众多,迄今已发现近80个,涉及细胞周期调控、细胞凋亡、分化或病毒免疫应答等多种生物学过程。TRIM家族(又名RBCC家族)成员含有RING指结构域(RING-finger domain),因而可以作为泛素E3连接酶发挥功能。除了RING指结构域外,TRIM蛋白还含有一个或者两个锌离子结合结构域(B-boxes domain)和卷曲螺旋结构域(coiled coil domain),以及复杂的C-端结构域。
TRIM家族蛋白质根据C-端结构域的差异可分为11个亚家族(C-I~C-XI)。TRIM46属于C-I亚家族,C-I亚家族包括6个成员:MID I(TRIM18)、MID2(TRIM1)、TRIM9,TNL(TRIM67)、TRIM36(HAPRIN)和TRIFIC(TRIM46)。表1总结比较了这些基因的功能及相关疾病。C-I亚家族每个基因不仅含有一个RBCC结构域,包括两个B-boxes,还包括C末端的COS box,Fibronectin Type III motif和SPRY/B30.2样结构域。这些基因又可以分为3组:MIDI和MID2同源性高达76%,但是和其他成员同源性低于25%;TRIM9与TNL最为相似,同源性高 达65%;而HAPRIN(TRIM36)与TRIFIC(TRIM46)最为接近,同源性高达43%。
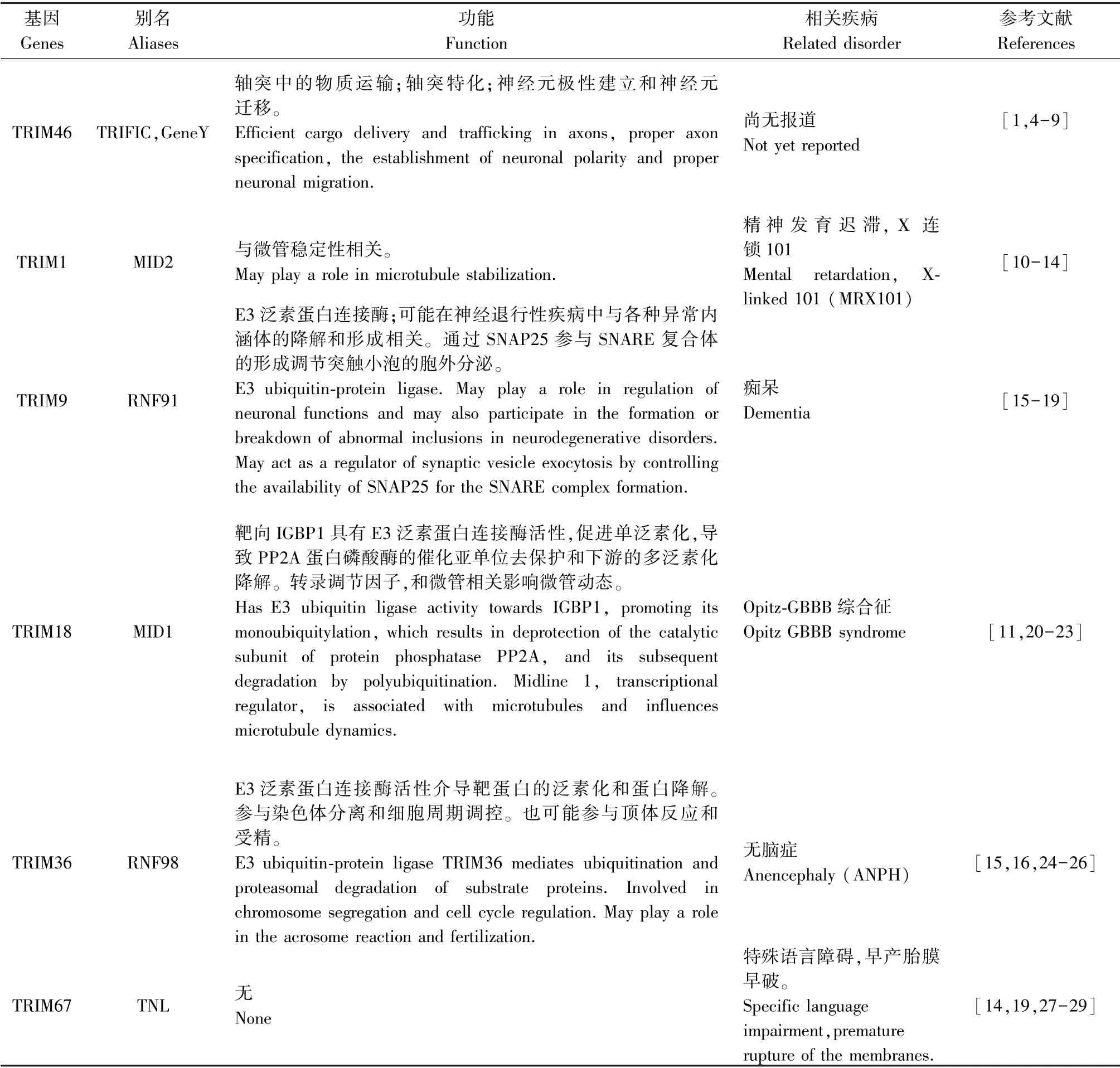
表1 TRIM C-I亚家族成员基因功能及相关疾病的比较Table1 Gene function and related disorders of TRIM C-I subfamily members
2 TRIM46基因概述
2.1 TRIM46蛋白结构和功能
TRIM46基因位于人染色体1q21,又命名为GENEY,编码的蛋白含有一个N末端的RING finger结构域、两个B-box motif、一个coiled-coil region、一个COS box、一个FN3和羧基端的一个B30.2结构域[14]。TRIM46蛋白是一个新发现的三重基序(tripartite motif,TRIM)家族蛋白。TRIM46蛋白含有759个氨基酸,分子量约为83×103。TRIM46含有RING指结构域(RING-finger domain),因而可能具备E3泛素-蛋白连接酶功能。从四级结构上预测,TRIM46可以与TUBB3蛋白以及TUBA4A蛋白发生相互作用。大多数TRIM蛋白从N端到C端的顺序依次是RING锌指结构,一个或两个B-box结构和一个卷曲螺旋结构域(coiled-coil,CC),又被称为RBCC结构域。如图1所示,TRIM46含有RBCC结构,在转染的COS-1细胞上,可与微管结构共定位。1-166蛋白区域决定了其在神经元近端轴突的定位,411-429蛋白区域决定了其与微管结合和共定位。
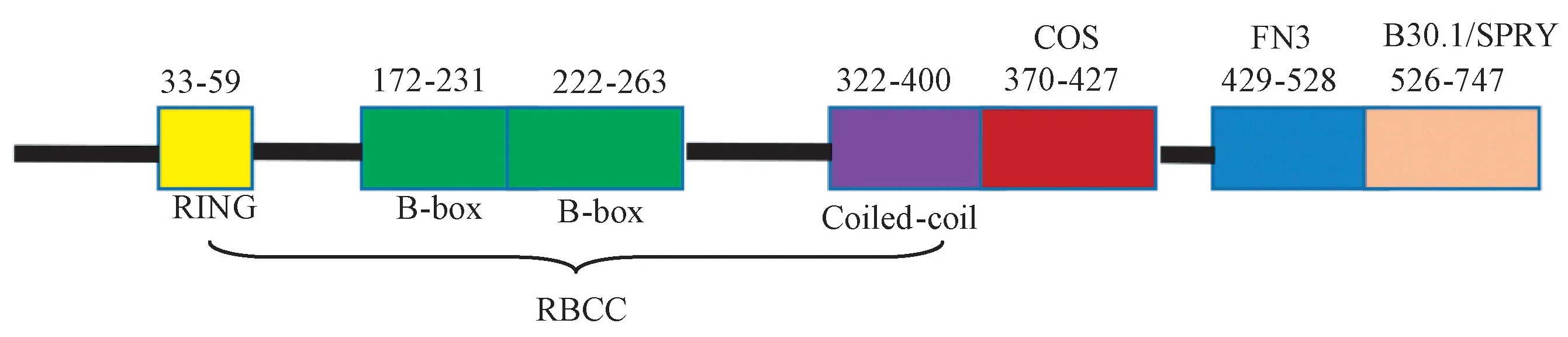
图1 TRIM46蛋白结构示意图Figure1 Schematic representation of overall domain architecture of human TRIM46protein
2.2 TRIM46的表达
人类蛋白图谱数据库(human protein atls,HPA)中RNA-seq数据显示,TRIM46蛋白在肾上腺、阑尾、脑、十二指肠、胆囊、淋巴结、脾、胃和睾丸等组织中广泛表达,其中脑和男性生殖系统附睾中表达最高。TRIM46在各个脑区均有表达,其中脑皮层、小脑和基底核中表达丰富,海马和杏仁核中等水平表达,丘脑表达相对较少。TRIM46在上皮细胞、内分泌细胞、神经细胞、胶质细胞、生殖细胞、血管细胞、肌肉细胞、间质细胞和血液或免疫细胞等多种类型细胞中表达,其中神经细胞、生殖细胞和单核细胞表达最多。在各种细胞系中,人神经母细胞瘤细胞SH-SY5Y中TRIM46的表达最高。目前已经有实验验证报道的,是TRIM46在神经元的轴突起始段(axon initial segment,AIS)特异表达[1,5]。
3 TRIM46功能与相关疾病
3.1 TRIM46与肿瘤
迄今,TRIM46与肿瘤相关研究较少。从基因位置上看,人和小鼠的TRIM46基因均坐落于肿瘤靶点基因MUC1的下游,在人类1号染色和小鼠3号染色体中,两个基因作为同一个基因簇的一部分。2015年国外的报道发现了在卵巢恶性肿瘤标本和细胞中特征性表达MUC1-TRIM46-KRTCAP2的嵌合RNA[30],而在非肿瘤卵巢组织中检测不到该嵌合RNA。此外,胃癌的基因组关联分析中显示MUC1/TRIM46的SNP位点rs2070803与患者的生存预后密切相关[31]。功能研究方面,2016年Zhang等[2]报道了TRIM46是小鼠mir-1894-3p的靶基因,敲低TRIM46表达可抑制乳腺癌细胞的增殖和迁
移。随后2017年又有国内研究报道[32],TRIM46通过抑制SH2结构域酪氨酸磷酸酶1(Src homology2(SH2)-containing tyrosine phosphatase1,SHP1)表达,促进HCT116结直肠癌细胞的增殖和侵袭。2019年的研究将FAM72B、GNE和TRIM463基因视为前列腺癌病人的预后基因表达分类器[33]。由于TRIM46与TRIM36序列高度相似,后者参与了细胞膜间的黏附[34],提示TRIM46也可能通过与细胞外基质互作,促进肿瘤细胞的增殖和迁移。最新的研究揭示了TRIM46通过促进PPARα的泛素化激活NF-κB信号通路参与骨肉瘤细胞存活和凋亡抑制的新机制[35]。
3.2 TRIM46与神经元极化
轴突始段(axon initial segment,AIS)在神经元极性形成、动作电位引发、以及脑损伤和脑疾病中起重要作用。研究证实,TRIM46特异定位于近端轴突,在AIS结构维持以及神经元极性的生成/维持中发挥了重要功能[1,4-9]。TRIM46可诱导平行微管束化[4],平行维管束通过跨桥连接而成,决定了神经元极化的方向[1,4]。同时这些维管束对于神经元轴突内的有效运输也非常重要。TRIM46部分敲低,导致微管运输增加,微管在远端轴突异常积累[9]。但是,目前关于TRIM46蛋白在神经组织中参与神经元极化过程的机制尚不明确,研究很少,目前已知的是TRIM46组织轴突起始段的微管束化,决定极化的方向,可能是通过KIF3/KAP3/TRIM46转运调节MARK2信号对轴突起始段进行时空组织构建[6]。如图2所示,TRIM46与AIS其它蛋白成分480AnkG和NF-186等协同作用,参与轴突形成和神经元极化。
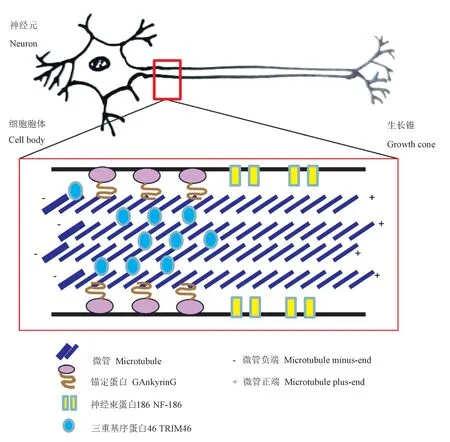
图2 TRIM46参与近端轴突的微管组织Figure2 TRIM46participates in microtubule organization in the proximal axons
3.3 TRIM46与痛风、炎症
TRIM家族与先天免疫相关[36],TRIM46也同样可能在免疫类疾病的调节方面扮演重要角色。基因组关联分析显示,TRIM46的单核苷酸多态性(SNP)与血清中尿酸浓度相关[37]。高尿酸血症与一种复杂的关节炎症痛风相关。一项国内的研究显示,TRIM46的两个SNP位点rs4971101和rs2070803与痛风的敏感性密切相关[38],提示了痛风的遗传因素,未来通过基因修饰动物构建也可解决痛风模型困难问题[39]。艰难梭菌B(Clostridium difficiletoxin B)诱发肠道炎症导致腹泻,严重可以致死。最新研究发现,TRIM46可作为一个新的免疫调节因子,调控艰难梭菌毒素B(TcdB)诱导的慢性炎症[3]。具体机制是TRIM46参与DUSP1泛素化,TRIM46敲低表达抑制TcdB诱导的MAPKs与NF-κB信号激活和炎症因子IL-1β和TNF-α的产生,反之,TRIM46过表达则促进炎症。
3.4 TRIM46与副肿瘤综合症
副肿瘤综合症(paraneoplastic neurological syndromes,PNS),是一种与肿瘤产物相关的异常的免疫反应,可诱发神经、消化、内分泌和造血等多系统病变和相应的临床症状出现。PNS病人血清或者脑脊液会存在各种抗神经元抗体,而这些抗体的检测是PNS诊断和寻找潜在肿瘤的有用工具。最近研究报道发现,TRIM46抗体可在多种神经综合征中检测到,且与小细胞肺癌(small-cell lung carcinoma,SCLC)相 关[40]。TRIM46特 异 定 位 于AIS,而其他AIS成分蛋白抗体在PNS及其他神经疾病中已有报道。比如AIS另一成分蛋白NF186抗体可在外周神经病变中检测到。
4 研究展望
TRIM家族蛋白具有3种类型的结构域:锌指结构(RING finger)、B-box和卷曲螺旋(coiled coil)。B-box是引起炎症反应的功能结构域,RBCC结构域决定了与微管细胞骨架的结合。CC结构域决定了蛋白自身的相互作用和同源二聚体的形成。TRIM蛋白还作为E3连接酶参与泛素化过程,转移泛素到特定的靶点上。正是由于结构的多样性,决定了其功能的多样性。
TRIM46与家族其他成员类似,与细胞骨架相关参与微管组装调控,也参与蛋白的泛素化,推测其与神经系统疾病或者精神类疾病有着密不可分的关系。同时,TRIM46目前的研究大多采用siRNA敲低其表达,离体条件下研究其功能缺损导致的表型变化。尚缺乏基因敲除动物或者转基因过表达动物模型,以及利用动物模型开展的功能获得或者功能丢失的条件下的活体表型研究。因此,TRIM46未来的研究可能会聚焦3大方向:神经系统疾病、免疫系统疾病以及肿瘤领域也将涌现出更多关于TRIM46作为肿瘤标记物或者预后标记物的研究。从技术手段上讲,利用新的基因编辑技术创建TRIM46过表达或者敲除动物模型便于更好开展TRIM46新功能研究,以及与疾病相关的分子机制研究。
猜你喜欢
安徽医科大学学报(2022年10期)2022-11-03
中国生物化学与分子生物学报(2022年4期)2022-09-07
生物化学与生物物理进展(2022年8期)2022-08-20
湖北农业科学(2022年11期)2022-07-18
浙江临床医学(2021年12期)2021-11-29
心血管病学进展(2021年8期)2021-09-13
湖北农业科学(2020年24期)2021-01-21
实用肿瘤学杂志(2020年4期)2020-12-08
医学综述(2011年12期)2011-12-09
亚热带农业研究(2011年3期)2011-09-29
