
产酸丙酸杆菌FS1171产丙酸发酵动力学研究
2022-03-22 02:39:38曾美端洪艺萍蔡玉凤周明炀成立文
微生物学杂志 2022年6期
曾美端, 洪艺萍, 蔡玉凤, 周明炀, 成立文, 郑 毅,3*
(1.福建师范大学 生命科学学院,福建 福州 350117;2.福州工业微生物科技有限公司,福建 福州 350007;3.福建师范大学 工业微生物教育部工程研究中心,福建 福州 350117)
丙酸是重要的化工原料,广泛应用于食品、饲料、塑料、香料、医学等领域[1]。丙酸的钙、钠和钾盐可用作防腐剂,是世界上公认的经济实惠且十分安全的食品添加剂[2-3]。丙酸可通过化学合成法和微生物发酵法生产。化学合成法合成效率高,但副产物治理难度大、治理成本较高[4]。微生物发酵法具有反应条件温和、菌种易得、易于培育、生产成本较低等优点[2]。因此,微生物发酵法成为了丙酸生产的主要方法。近年来,对于微生物发酵法生产丙酸的研究主要集中在以下几个方面:一是耐酸菌株的筛选,孙丹等[5-6]通过压力筛选的方法,获得了一株耐25 g/L丙酸的产酸丙酸杆菌;二是耐酸机制的研究,发现确定基于氨基酸代谢的耐酸系统是其响应丙酸胁迫的主要调控元件[7];三是发酵过程的优化,通过研究培养基组分对发酵生产丙酸的影响,发现当以葡萄糖和甘油为混合培养基碳源时,丙酸产量提高了26.36%[8];四是衍生物的开发利用,丙酸盐如丙酸钙等因对霉菌和芽胞杆菌有良好的抑制作用,常用做谷物和饲料的防腐及食品的保鲜[9-10];但在发酵动力学研究方面未见相关报道。通过发酵动力学的研究和发酵模型的建立,能够进一步了解发酵过程中菌体生长与产物形成的机制,以及环境因素对这些机制的影响,实现发酵过程的有效控制,达到提高产物发酵指标的目的[11]。本研究针对产酸丙酸杆菌(Propionibacteriumacidogenes) FS1171产丙酸分批发酵过程,利用经典发酵动力学模型和Origin软件优选模型方程,分别构建产丙酸分批发酵菌体生长模型、丙酸生成模型和底物甘油消耗模型。通过拟合方程可以预测菌体生长量、丙酸生成量以及甘油消耗情况,为进一步放大试验,优化和工业化生产提供参考。
1 材料与方法
1.1 材料
1.1.1 菌种 产酸丙酸杆菌(Propionibacteriumacidogenes)FS1171,由福建师范大学工业微生物教育部工程研究中心新资源开发课题组实验室保藏。
1.1.2 培养基(g/L) ①种子培养基:甘油5,蛋白胨5,酵母膏10,K2HPO42.5,KH2PO41.5,水1 000 mL,10% KOH调pH 6.8~7.2;②发酵培养基:甘油40,蛋白胨5,酵母膏10,K2HPO42.5,KH2PO41.5,水1 000 mL,10% KOH调pH 6.8~7.2。
1.1.3 主要试剂与仪器设备 甘油、酵母膏、蛋白胨、琼脂、磷酸氢二钾、磷酸二氢钾、氢氧化钙、氢氧化钾、乙腈、磷酸氢二钠、磷酸(分析纯)。高效液相色谱仪(20AD,日本岛津公司);立式蒸汽压力灭菌器(GI54DWS,致微(厦门)仪器有限公司);台式离心机(TDZ4-WS,长沙湘仪离心机仪器有限公司);超净工作台(ZHJH-C1112B,上海智城分析仪器制造有限公司)。
1.2 方法
1.2.1 丙酸发酵培养方法 ①一级活化种子液制备:将甘油保存的FS1171菌种接种到装有100 mL液体培养基的250 mL锥形瓶中,30 ℃静置培养48 h,获得活化一级种子液;②二级种子液制备:将一级活化种子液按5%(体积分数)的接种量转接到新鲜的种子培养基中,30 ℃静置培养24 h,获得液体发酵的二级种子液;③丙酸发酵培养:将种子液按10%(体积分数)的接种量转接到发酵培养基中,30 ℃静置培养,发酵期间每隔24 h测定pH,同时用25%(质量分数)Ca(OH)2溶液将发酵液的pH调节至6.8~7.2。
1.2.2 分析方法 ①生物量的测定:采用核酸法测定[12],用菌体的核酸量作为生物量测定的指标;②甘油浓度的测定:采用比色法测定发酵液中甘油的残余量[13];③丙酸浓度的测定:采用高效液相色谱法(HPLC)进行测定[14]。
1.2.3 发酵过程曲线的绘制 产酸丙酸杆菌FS1171在产丙酸分批发酵过程中每隔24 h取样,用1.2.2的方法进行菌体生物量、丙酸生成量和底物甘油消耗量的测定。选用Origin 8.6软件制作产酸丙酸杆菌的发酵过程曲线。
1.2.4 经典模型丙酸杆菌发酵动力学的建立 应用Origin 8.6软件对实验所得数据进行非线性高级拟合分析,求解得出动力学模型参数,建立菌体生长、丙酸生成量和甘油消耗的动力学模型方程[14]。
①菌体生长动力学模型的建立:菌体生长动力学模型建立最常见的是Logistic模型和Monod方程。Logistic模型是一个非常典型的S型曲线方程,常被用来描述菌体浓度与营养物质之间的非线性关系[15]。采用Logistic方程构建丙酸发酵菌体生长的动力学模型。
(1)
方程式(1)积分得:
(2)
式中:X为菌体生物量(g/L);μm为菌体最大比生长率(d-1) ;Xm为最大菌体生物量(g/L);X0为初始菌体生物量(g/L);t为发酵时间(d )。
②产物丙酸合成动力学模型的建立:在微生物丙酸生物合成过程,选用经典的Luedeking-piret方程构建动力学模型,公式如下:
(3)
因为ɑ≠0,β≠0,结合公式(2)和(3),方程可化为:
(4)
式中:P0代表初始丙酸的产量,P0=0 ,X0对产量的影响可以忽略不计,方程可简化为:
(5)
式中:ɑ、β为生长偶联参数;t为发酵时间(d);P为丙酸浓度(g/L)。
③底物甘油消耗动力学模型的建立:选用Dose-Resp模型分析甘油的消耗过程,方程式为:
(6)
积分得:

(7)
式中:S为甘油的浓度(g/L) ;S0为初始甘油浓度(g/L);X为菌体生物量(g/L);t为发酵时间(d);P为产物丙酸浓度(g/L);M0为菌体细胞维持相关常数;YX/S为最大细胞得率数;YP/S为产物丙酸得率系数。
1.2.5 Origin软件筛选最优数学模型构建丙酸发酵动力学模型 采用Origin 8.6软件运用不同的模型(Logistic方程和BoLtzmann方程)对菌体生长动力学模型、产物生成动力学模型和底物消耗的动力学模型进行拟合。
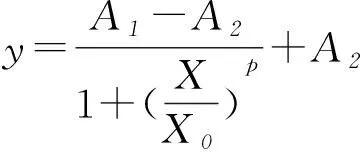
(8)
式中:A1为初始的菌体浓度;A2为最终的菌体浓度;X0、p为方程系数。
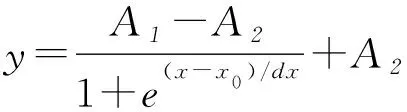
(9)
式中:A1为初始的菌体浓度;A2为最终的菌体浓度;X0、dx为方程系数。
2 结果与分析
2.1 产酸丙酸杆菌FS1171产丙酸分批发酵过程
产酸丙酸杆菌FS1171产丙酸分批发酵过程中生物量(X)、丙酸产量(P)、甘油消耗(S)随发酵进程时间(t)的变化情况见图1,发酵过程中菌体很快就进入对数生长期。在对数生长期内,菌体迅速生长并伴随甘油的显著下降,丙酸合成,96 h后菌体生长缓慢后趋于稳定,为生长的稳定期。发酵后期,由于没有补充甘油以及丙酸的积累,限制了细胞继续生长及其代谢活动,影响了丙酸的生成,菌体量也有所下降[16-18]。发酵10 d后,菌体生物量为0.323 g/L,丙酸生成量为37.58 g/L。在发酵过程中定期调节发酵液的pH值,则有利于维持菌体的生长代谢活动和丙酸的生成[19]。由此可知,产酸丙酸杆菌在发酵过程中,甘油的消耗和丙酸的合成符合菌体生长的代谢规律。

图1 产酸丙酸杆菌FS1171产丙酸分批发酵曲线Fig.1 Fermentation curve of Propionibacterium acidogenic FS1171 producing propanoic acid
2.2 基于经典发酵动力学丙酸发酵动力学模型的构建
2.2.1 菌体生长动力学模型 选用Logistic方程来拟合丙酸杆菌的菌体生长动力学模型,积分公式为式(2)。利用Origin 8.6对实验数据进行非线性拟合,得丙酸杆菌菌体生长动力学参数为X0=0.038 5,Xm=0.328 7,μm=1.830 8。将相关参数代入式(2)得菌体生长的动力学方程如下:
(10)
菌体的生长动力学曲线如图2所示,R2为0.949 3,图中可以看出实验值和Logistic方程拟合的数据能够很好地吻合,可以较好地描述丙酸杆菌菌体生长过程。

图2 菌体生长拟合曲线Fig.2 Growth fitting curve of strain FS1171 producing propanoic acid
2.2.2 丙酸合成动力学模型 丙酸合成动力学模型选用经典的Luedking-piret方程来描述,积分公式为式(5)。将丙酸杆菌发酵过程中丙酸生成量的变化数据代入Origin 8.6进行非线性拟合,得丙酸合成动力学参数为α=51.357 2,β=8.153 6,为部分相关型发酵,丙酸生成存在与菌体生长相联与不相联部分[20]。将α、β的数值带入式(5),获得丙酸的浓度与发酵时间的关系式,结果如下:
(11)
丙酸合成的拟合曲线如图3所示,R2为0.979 9,由图3可知菌体进入生长稳定期后,丙酸生成量的增长也呈较稳定增长趋势,实验值和Luedking-Piret方程拟合的数据能够较好地吻合,可以较好地描述丙酸生成过程。
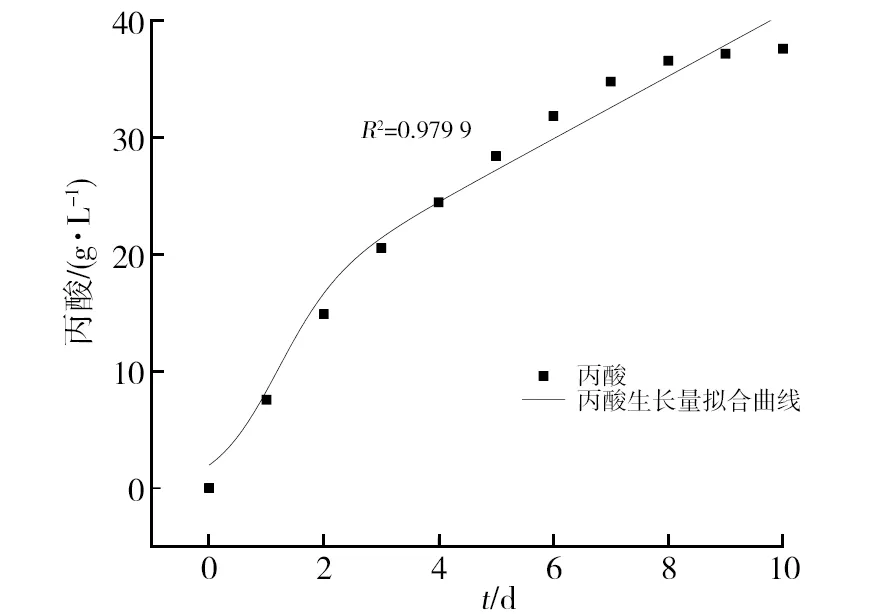
图3 丙酸生成量拟合曲线Fig.3 Fitting curve of propionic acid production of strain FS1171
2.2.3 甘油消耗动力学模型 采用Dose-Resp模型来分析发酵底物甘油的消耗过程,积分公式为式(7)。将丙酸杆菌发酵过程甘油消耗的变化数据代入Origin 8.6进行非线性拟合,得底物消耗动力学参数为S0=32.333 2,1/Y(X/S)=234.955 2,1/Y(p/S)=-3.511 9,M0=26.545 3;将相关数值代入公式(7)得:
S(t)=41.39-54.593 8*X(t)+
(12)
甘油消耗拟合曲线如图4所示,R2为0.981 5,从图4中可以看出实验值和Dose-Resp方程拟合的数据能够较好地吻合,可以较好地描述底物消耗过程。
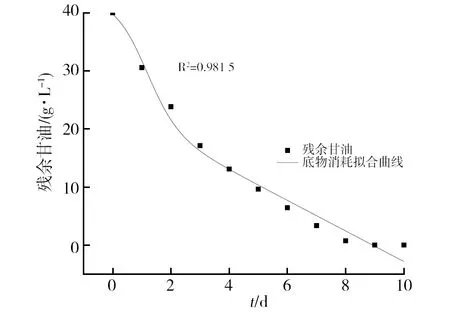
图4 底物消耗拟合曲线Fig.4 Substrate consumption fitting curve of strain FS1171
2.2.4 模型的验证 表1是基于经典发酵动力学模型(Logistic、Luedeking-Piret、Dose-Resp方程)构建的模型拟合值与实验值的比较。从表1中可以看出,模型拟合值与实验值之间的误差率大都在10%以下,虽然在发酵末期,甘油消耗模型的拟合值与实验值之间已达到10%以上,但此时甘油浓度很低且到后面保持不变,此后的甘油浓度对发酵结果影响不大。因此,该发酵动力学方程构建的模型能较好地描述产酸丙酸杆菌FS1171发酵过程中菌体生长、丙酸合成以及甘油消耗的变化规律。

表1 基于经典发酵动力学方程构建模型拟合值与实验值的验证比较Table 1 Verification comparison of fitting value and experimental value of model based on classical fermentation kinetic equation
2.3 基于Origin软件筛选最优数学模型构建丙酸发酵动力学模型
2.3.1 不同动力学模型方程的筛选与评价 采用Origin 8.6软件中的两种模型(Logistic和Boltzmann方程)分别对菌体生长、丙酸合成以及甘油消耗的动力学模型进行拟合,评价结果见表2。结果表明,Boltzmann模型的拟合效果比Logistic更为理想,能更好地描述产酸丙酸杆菌FS1171在发酵过程中菌体生长、丙酸合成以及甘油消耗的情况。

表2 两种模型发酵动力学拟合评价Table 2 Fitting evaluation of fermentation kinetics for two models
2.3.2 基于Boltzmann方程构建拟合曲线 根据Boltzmann方程构建菌体生长、丙酸合成及甘油消耗的动力学模型拟合曲线,结果如图5~图7所示,其拟合相关系数R2分别为0.992 3、0.998 3和0.997 6,可以很好地反映菌体生长、丙酸合成以及甘油消耗的发酵过程。
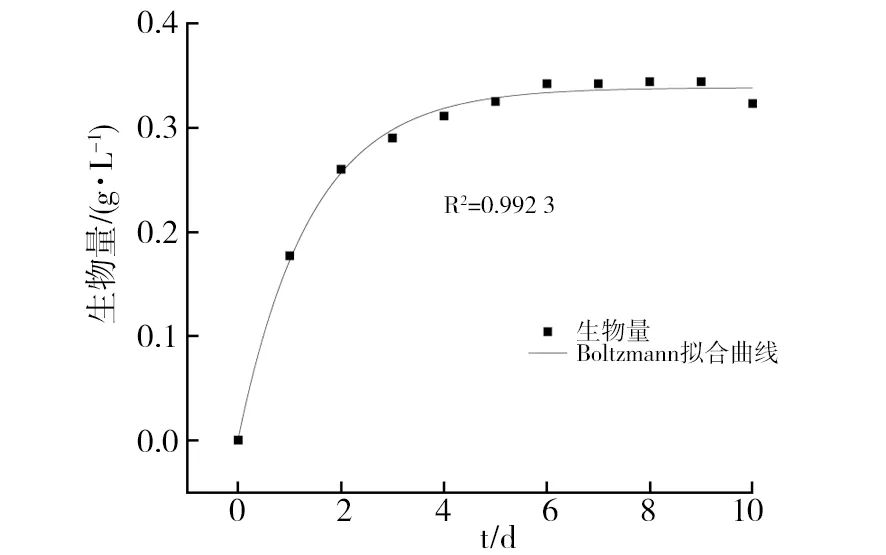
图5 基于Boltzmann FS1171模型生长拟合曲线Fig.5 Growth fitting curve of strain FS1171 based on Boltzmann model
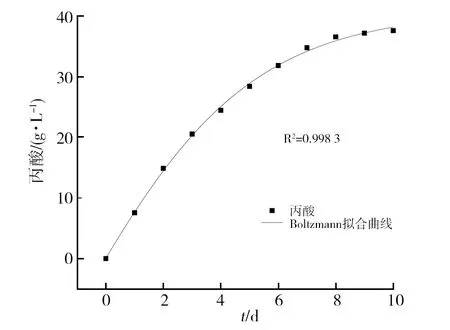
图6 基于Boltzmann模型的丙酸生成拟合曲线Fig.6 Fitting curve of propionic acid production based on Boltzmann model
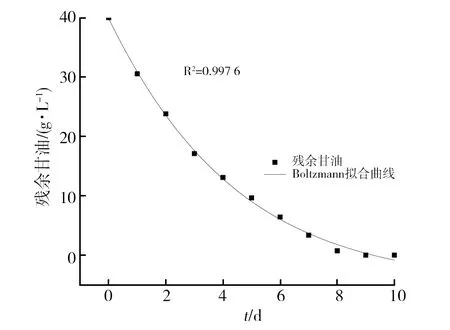
图7 基于Boltzmann模型的底物消耗拟合曲线Fig.7 Substrate consumption fitting curve based on Boltzmann model
2.3.3 模型的验证 表3是基于Boltzmann方程构建的模型拟合值与实验值的比较。同表1出现类似的现象,在参数低浓度区域拟合相对误差较大,出现一定失真,这可能是构建模型应该注意的,好在低浓度区域对整个发酵过程相对影响度较少。

表3 基于Boltzmann方程构建模型拟合值与实验值的验证比较Table 3 Verification comparison of fitting value and experimental value of model based on Boltzmann equation
3 讨 论
基于经典发酵动力学模型(Logistic、Luedeking-Piret、Dose-Resp方程,简称方法Ⅰ)成功构建产酸丙酸杆菌(Propionibacteriumacidogenes) FS1171丙酸分批发酵过程中菌体生长、丙酸合成及甘油消耗的动力学模型,其拟合度R2分别为0.949 3、0.979 9和0.981 5,数值均大于0.9,说明模型能够很好地反映该菌丙酸发酵动力学。通过比较3个模型的拟合值和实验值可以发现,拟合相对误差较大的区域主要发生在参数低浓度区域,如菌体生长的前期、产物合成的初期与底物消耗的末期,主要原因在于方程主要反映菌体浓度的增加和营养物质的消耗对自身生长的抑制与对产物合成的影响。在低浓度区域,这些参数变化幅度不明显,因此拟合情况相对不够理想。
基于Boltzmann方程(简称方法Ⅱ)构建的动力学模型,其拟合度R2均大于0.95,在参数低浓度区域拟合相对误差也较大。对比两种发酵动力学构建方法,方法Ⅰ所构建模型拟合度(R2值)均小于方法Ⅱ拟合度(R2值),前者模型拟合值与实验值之间的误差率大都在10%以下,而后者模型的偏差率大都在5%以下,因此明显发现前者所构建模型拟合度整体不如后者,但前者所构建的丙酸生成与底物消耗模型都是以菌体生长动力学模型为基础的,3个模型之间有较好的关联性及参数解析,而方法Ⅱ构建的3个模型参数之间的关联性较差,呈现一定独立性,仍需对参数含义进一步解读,以期获得更好的应用效果。
通过丙酸发酵过程动力学研究,可更准确地对实验指标进行预测,实现发酵过程的优化,并可利用小实验得到的数据设计大型的发酵工艺,为进一步放大实验和工业化生产提供理论基础。Zakeri等[21]利用非结构模型建立的动力学方程可以成功预测甘蔗糖蜜生产黄原胶的发酵过程,为黄原胶的工业化生产奠定了一定的理论基础。现有报道通过单因素实验、正交实验以及响应面法来实现丙酸发酵过程的优化[22-25],但通过构建丙酸发酵动力学模型来进行发酵过程优化的研究未见相关报道。今后的试验可以进一步从优化发酵工艺入手,对补料发酵过程进行系统化的研究,构建补料发酵的动力学模型,获得良性循环的补料策略,为丙酸杆菌实现工业化生产甚至应用于连续发酵奠定基础。
猜你喜欢
中国当代医药(2019年20期)2019-09-25 05:03:40
农业机械学报(2019年1期)2019-02-15 07:30:26
能源(2017年7期)2018-01-19 05:05:04
中学科技(2017年11期)2017-12-26 10:14:01
中国美容医学(2016年4期)2016-07-25 06:16:52
中国民族医药杂志(2016年4期)2016-05-09 07:41:11
现代食品(2016年24期)2016-04-28 08:12:06
化工进展(2015年3期)2015-11-11 09:07:41
医学研究杂志(2015年5期)2015-06-10 06:43:26
应用化工(2014年3期)2014-08-16 13:23:50
