
甘肃玉米茎腐病病菌禾谷镰孢复合种遗传多样性
2022-03-17 08:12王宝宝张小杰王春明周天旺李敏权郭致杰
西北农业学报 2022年3期
郭 成,王宝宝,邓 东,张小杰,王春明,周天旺,李敏权,郭致杰
(1.甘肃省农业科学院 植物保护研究所,兰州 730070;2. 甘肃农业大学 草业学院,兰州 730070;3. 石家庄市农林科学研究院,石家庄 050041;4. 中国农业科学院 作物科学研究所,北京 100081;5. 甘肃农业大学 植物保护学院,兰州 730070)
玉米是中国种植面积最大的农作物[1]。在2021年“秸秆畜牧业”以及“粮改饲”的形势和政策下,粮饲兼用型玉米具较大的发展潜力,能实现粮食和饲料有效补给,是国内粮食安全生产的关键环节[2]。一般年份,茎腐病田间发病率在5%~10%,重发年份达到20%~30%,高感品种的发病率可高达40%~100%,发病率每增加1%,单穗籽粒损失率约提高0.485%,千粒质量损失率约提高0.304%,若发病严重将引起30%以上的产量损失[3]。受玉米连年种植、秸秆还田以及气候逐年变暖等因素影响,田间土壤中病原菌基数逐年上升,玉米茎腐病呈逐年加重趋势[4]。据郭成等[5]连续多年调查发现,在甘肃及周边地区玉米茎腐病自然发生率偏高,玉米茎腐病对产量的制约已成为西北地区以及其他玉米主产区生产上的重大问题。
玉米茎腐病的致病菌种群结构和数量较为复杂,目前已报道玉米茎腐病的病原菌多达30种,其中禾谷镰孢(Fusariumgraminearum)和肿囊腐霉(Pythiuminflatum)为国内玉米茎腐病的两大主要致病菌[5]。禾谷镰孢复合种种群组成复杂,已探明包含超过10个菌种,其对环境有广泛的适应性,不仅能够侵染多种水果、蔬菜、花卉,还引起小麦和大豆、马铃薯、玉米等多种作物病害的发生[6-11]。病原菌群体遗传多样性越丰富表明病原菌对环境的适应力越强,较高遗传多样性的致病菌具有较强的进化和生存优势[12]。ISSR、VNTR、RAPD、AFLP和SSR等分子标记技术广泛应用于禾谷镰孢复合种群体遗传结构分析[13-18]。禾谷镰孢复合种侵染作物后,在寄主体内产生多种次生代谢物和毒素,这些毒素贮藏在植物体内潜在危害巨大[19-20]。
研究病原菌群体结构及其遗传多样性特征是应对植物病害的发生、流行及其防治的关键[21]。解析致病菌遗传分化特征及其致病力是抗病品种选育及病害防控的研究基础[22-23]。有效控制玉米茎腐病对保障中国玉米产业可持续发展具有重要意义。目前缺乏对甘肃省玉米茎腐病致病菌遗传多样性的系统报道,本研究以甘肃省玉米茎腐病优势病原菌禾谷镰孢复合种为对象,研究其在4大生态区的分布特征及遗传多样性,该结果将为甘肃省玉米生产提供风险警示,为玉米镰孢茎腐病防控提供基础信息。
1 材料与方法
1.1 菌株信息
从甘肃省4大生态区采集茎腐病样,采用组织分离法,即在样品病健交界处用剪刀剪取约 (0.2×0.5) cm大小的玉米茎秆及维管束组织,参照文献[24-25]进行分离、培养和纯化,获得菌株。经致病性测定后,采用形态学观察结合TEF-1α基因测序并将所得序列在NCBI中心的BLAST模块进行比对分析[18],鉴定到具体种,从中选取114株禾谷镰孢复合种(FGSC)菌种,平均分布在4个生态区。菌株分布地点见表1。

表1 禾谷镰孢复合种采集信息Table 1 Distribution and number of F.graminearum species complex
1.2 供试引物
遗传多样性引物采用任旭等[17]和Qu等[26]使用的8对VNTR(序号g1~8)和10对SSR引物(序号g9~18),引物由上海生工生物工程有限公司合成(表 2)。

表2 禾谷镰孢复合种遗传多样性引物Table 2 Genetic diversity primer of F.graminearum species complex
1.3 PCR 扩增体系
参照贺小伦等[27]的方法以25 μL反应体系进行PCR扩增,退火温度按照引物进行相应的 设定。
1.4 凝胶电泳
对PCR扩增后的产物用6%的变性聚丙烯酰胺凝胶,以恒定功率75 W,电泳90 min,将凝胶置于显影液中进行条带自显影,条带显影后用蒸馏水冲去残液,将凝胶平铺在灯箱上统计条带,并拍照。
1.5 电泳谱带的统计及数据分析
统计好电泳DNA图谱的条带信息,有条带,统计为“1”,无条带,统计为“0”,进而将多态性位点,也以0和1的方式,转换为二进制数据。利用软件POPGENE来计算引物的遗传距离和遗传相似系数、多态位点数、Nei’s基因多样性指数(H)、群体间的Shannon’s多样性指数(I)[28]。利用软件NTSYSpc 2.11的非加权组平均法进行不同地区的菌群的SAHN聚类,构建聚类树。
2 结果与分析
2.1 TEF-1α基因序列聚类结果
禾谷镰孢复合种经形态学观察结合TEF-1α基因测序,在114株中鉴定出16株禾谷镰孢和98株布氏镰孢(F.boothii),选取部分菌株构建聚类树(图1)。

图中节点处的数字表示每100次聚类分析中能聚到同一个分支的次数
统计发现,在4大地区中均以布氏镰孢为主。陇南地区仅有布氏镰孢分布,23株,未分离得到禾谷镰孢;河西走廊布氏镰孢有27株,禾谷镰孢有3株;陇东地区布氏镰孢有20株,禾谷镰孢有11株;陇中地区布氏镰孢有28株,禾谷镰孢有2株(表3)。

表3 4个生态区禾谷镰孢复合种分布Table 3 Distribution of F.graminearum complex species in four regions

(续表3 Continued table 3)
2.2 18对引物的扩增结果
对VNTR和SSR引物扩增结果表明,114株禾谷镰孢复合种菌株多态性位点数为26。扩增片段集中在50~300 bp,其中引物扩增出的条带数均在1条以上,表明其多态性可观。
2.3 禾谷镰孢复合种不同地理种群遗传变异和遗传分化
POPGENE分析结果显示,4个生态区的禾谷镰孢复合种群体内的遗传分化系数为 0.924 9,而其种群间的遗传分化系数是0.075 1,群体内基因流为0.02,种群间的基因流为 6.153 5,说明禾谷镰孢复合种遗传变异主要来源于群体内。禾谷镰孢复合种群体总基因多样度为0.425 9,各地理群体内基因多样度为0.393 9,不同地区间基因多样度为0.032,综上,来自同一个生态区禾谷镰孢复合种菌株间携带的基因最为接近,遗传变异主要来源于群体内。
Shannon’s信息指数和Nei’s基因多样性指数均以陇中为最高,分别为0.608 7和0.422 9,表明该地区菌株的基因丰富度最高;陇东地区的次之,分别为0.601 7和0.416 8;而陇南地区的则分别为0.564 1和0.392 6;河西走廊的最低,分别为0.501 7和0.343 2,说明基因多样性与地理区域有较大的相关性。4大生态区Shannon’s信息指数均在0.50以上,Nei’s基因多样性指数均在0.34以上,表明甘肃省禾谷镰孢复合种菌群的遗传多样性十分丰富。如表4所示,4个生态区禾谷镰孢复合种的有效等位基因数(Ne)在 1.612 6~1.775 7,平均有效等位基因数为 1.714 0;多态位点百分率在88.46%~100%,平均为95.19%;多态位点数在23~26,平均为 24.75。研究结果表明,甘肃陇东地区和陇中地区菌株遗传多样性最为丰富。
2.4 禾谷镰孢复合种种群的遗传相似度和遗传距离
如表4所示,陇中地区和陇东地区的遗传距离最小,为0.033 1,两者的遗传相似度最大,为0.967 4,说明两地禾谷镰孢复合种菌株基因组成最接近;而陇东和陇南地区的遗传相似度最小,为0.888 0,其遗传距离最大,为0.118 8,说明两地菌株间差异性最大,亲缘关系最远。4个地区间遗传相似度为0.888 0~0.967 4,Nei’s遗传距离为0.033 1~0.118 8(表5),4个生态地区的菌株均具有丰富的遗传多样性。

表4 4个不同生态区禾谷镰孢复合种种群的遗传变异参数Table 4 Genetic diversity parameters of F.graminearum species complex among four different geographical populations

表5 甘肃不同生态区禾谷镰孢复合种种群的遗传相似度和遗传距离Table 5 Genetic similarity coefficient and genetic distance among different geographical groups of F.graminearum species complex among four different geographical populations in Gansu
2.5 聚类分析
依据遗传相似度,用NTSYSpc version 2.11软件对4个地区种群进行聚类分析(图2),当遗传相似系数为0.938 0时,4个生态群体被聚为3个分支。陇南地区与河西走廊的菌株各自聚为一大类群,分别为第Ⅰ类群和第Ⅱ类群;而陇东地区和陇中地区的菌株共同聚为一大类,为第Ⅲ类群。结果表明,陇东地区和陇中地区禾谷镰孢复合种菌群亲缘关系最为接近,4个生态区禾谷镰孢复合种群体按其亲缘关系可分为3大类,其遗传亲缘关系与地理来源有一定的相关性。
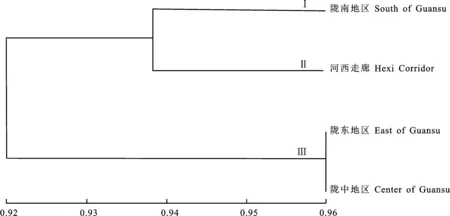
图2 禾谷镰孢复合种4个地理种群的UPGMA聚类图Fig.2 The UPGMA cluster map of 4 geographical populations of F.graminearum species complex
2.6 禾谷镰孢复合种种群变异分析
禾谷镰孢复合种不同地理种群的基因变异方差分析表明,在3大类群间的遗传变异为5.5%,4个种群间的变异占总变异的3.79%,种群内的变异占总变异的90.71%,同时P<0.001 0(表6),表明禾谷镰孢复合种种群的遗传变异主要发生于种群内部。

表6 禾谷镰孢复合种地理种群的种群变异分析Table 6 Analysis of molecular variance of F.graminearum species complex for geographical populations
3 结论与讨论
为明确甘肃4大生态区玉米镰孢茎腐病优势病原菌禾谷镰孢复合种遗传多样性,应用8个VNTR标记和10个SSR标记对自甘肃省4大生态区选取的 114 株禾谷镰孢复合种菌株进行分析,按遗传相似度划分为3大类群,表明禾谷镰孢复合种的亲缘关系与菌株的地理来源有一定相关性。Qu等[26]研究表明,来自中国不同地区的禾谷镰孢菌群体存在高度的遗传多样性,整个群体可分为两个类群,两个类群有不同的地理分布特征,每个类群又可进一步划分为几个亚类。与贺小伦等[27]对来自河南和河北冬小麦区收集的假禾谷镰孢菌株(F.pseudograminearum)进行遗传多样性研究,发现相似系数为0.966时,来自6个地理群体的166株菌被划分成 2 个大的类群;马红霞等[28]以相似系数 0.56为标准,将来自 11 个省(自治区)禾谷镰孢复合种的92 个菌株划分成 9 个类群;任旭等[17]将46 株禾谷镰孢复合种菌株以遗传相似系数0.45为标准划分为3个类群;何婧[29]在相似系数为0.682时,将115株禾谷镰孢划分为2个类群等研究结果相吻合,进一步说明禾谷镰孢复合种群体具有丰富遗传多样性且存在较明显的生态地理类型。
本研究发现,甘肃4个不同生态区禾谷镰孢复合种种群遗传变异主要来自种群内部。许多学者采用多种分子技术对禾谷镰孢的遗传多样性进行分析,得出的结果与本研究一致,如陈捷等[30]应用RAPD技术明确禾谷镰孢菌群间的遗传变异性,易受地域、环境和寄生部位选择作用的影响,出现明显的遗传分化特征; 纪莉景[31]利用TEF-1α基因测序结合RAPD技术发现中国部分地区玉米穗腐病和小麦赤霉病致病菌包括禾谷镰孢、亚洲镰孢(F.asiaticum)和布氏镰孢3个类群,且不同地理来源的类群内的菌株均存在丰富的遗传多样性;何婧[29]使用ISSR标记对玉米茎腐病致病菌禾谷镰孢分析发现,禾谷镰孢即使在同一聚类分支或是同一地区的菌株在遗传多样性上也存在差异,表明禾谷镰孢菌株间有明显的生理分化; Talas等[32]利用高通量测序技术对德国13块试验田的213个禾谷镰孢菌株进行群体基因组学分析,发现田块内群体具有较高的遗传多样性。
本研究首次阐释甘肃省玉米茎腐病优势致病菌禾谷镰孢复合种具有较为丰富的遗传多样性,根据遗传相似度将4个生态区的菌群划分为3大类群,且其种群遗传变异主要来自种群内部,但其致病力、产毒类型和种群地理分布特征三者之间是否存在相关性有待进一步研究。
猜你喜欢
新疆农业科学(2022年6期)2022-07-13
中国农学通报(2022年12期)2022-06-01
作物学报(2022年4期)2022-02-10
绿色科技(2019年14期)2019-11-19
中学生物学(2019年7期)2019-10-17
江苏农业科学(2019年5期)2019-09-02
江苏农业科学(2017年17期)2017-11-15
湖北农业科学(2017年15期)2017-09-09
江苏农业科学(2015年2期)2015-03-12
绿色科技(2014年1期)2014-11-28
